關帝山油松天然林林下植物組成及環境解釋
2017-04-01 02:37:51王慧南宏偉劉寧
生態環境學報 2017年1期
王慧,南宏偉,劉寧
山西農業大學林學院,山西 太谷 030801
關帝山油松天然林林下植物組成及環境解釋
王慧,南宏偉,劉寧*
山西農業大學林學院,山西 太谷 030801
作為森林生態系統的重要組成部分,林下植物在調控森林結構和功能方面發揮著重要的作用。通過對華北山地干旱區油松(Pinus tabulaeformis Garr.)天然純林林下植物群落的調查,探究林下植物物種組成與林分、環境因子之間的關系及其關鍵控制因子,旨在為華北山地干旱區林下生物多樣性保護和維持提供依據。結果表明,(1)在調查的樣地中,油松林下共出現植物物種72種,隸屬33科65屬,其中薔薇科、菊科、豆科物種種類豐富,占全部種數38.8%;林下灌木的物種多為喜光或兼性樹種,與油松林特殊的冠形有關,林下草本以多年生草本占優勢。(2)CCA排序較好地揭示了該區油松天然林林下植物的分布格局與環境因子的關系,生物因子與非生物因子共同對林下物種的分布格局產生作用,對林下灌木層和草本層的解釋率分別為59.92%和66.16%;其中,地形因子的作用大于林分因子和土壤因子,在油松天然林林下植物分布格局中起著舉足輕重的作用。林下灌木層物種多樣性在下坡最高、上坡最低,并表現為西北坡高于其他坡向;林下草本層物種多樣性隨坡位的變化與灌木層正好相反,在上坡最高、下坡最低,在所選樣方范圍內,草本層物種多樣性隨海拔的升高而升高。
華北山地;油松天然純林;林下植物;分布格局;環境因子
林下植物是森林生態系統中的重要組成部分,是森林生態系統中生物多樣性的主要來源,在維持森林群落內的營養平衡、生物多樣性、林下更新和生態系統長期穩定性方面有著不可忽視的作用(Nilsson et al.,2005;Chastain et al.,2006;Gamfeldt et al.,2013;Wu et al.,2014)。近年來,除林下植物結構特征、分布格局、生態功能等被廣泛研究外,環境因子與林下植被分布格局的影響也已被廣泛討論,逐步得到重視(叢靜等,2013;劉宏文等,2014;尤業明等,2016)。
山地植物群落物種多樣性的變化規律一直是生態學家感興趣的問題(叢靜等,2013)。干旱區因難以保持水分需求平衡而成為全球生態脆弱區,也是生物多樣性保護和維持的重要研究區域。華北山地半干旱區森林生態系統是研究干旱區山地植物群落多樣性的理想場所。油松(Pinus tabulaeformis Garr.)是華北地區的特有樹種,也是華北中、低海拔山地的主要優勢樹種,在該區海拔1200~1800 m廣泛分布且長勢良好;遼東櫟(Quercus liaotungensis)和山楊(Populus davidiana)為其主要伴生的闊葉樹種。一般認為,油松林所具有的獨特的林冠結構和群落條件,導致其林下環境極差難以形成連續的林下植被層,以致油松純林結構單一而形成“綠色沙漠”(郭華等,2005),因此華北山地垂直層次分明的油松林則具有一定的群落多樣性研究價值。
本研究以華北山地油松天然林群落為研究對象,利用典型對應分析(CCA排序)探究林下植物群落組成與環境因子之間的關系,確定華北半干旱區影響油松天然林林下物種分布的主要因子,以期為其林下生物多樣性保護提供有力的理論依據。
1 材料與方法
1.1 研究地概況
研究區位于山西省中部呂梁山脈,地理坐標為東經110°0′~111°37′,北緯34°34′~37°43′,海拔為770~2850 m。研究區屬溫帶大陸性季風氣候,冬季寒冷干燥,夏季炎熱多雨,降水主要集中在6—8月,約占全年的60%以上,降水量在400~500 mm之間,蒸發量在1600 m以上,無霜期平均145 d。山地成土母巖主要有石灰巖和礫巖等,土壤為風積黃土母質,表層土壤類型主要為褐土,土層10~60 cm厚薄不等,腐殖質層0~10 cm,一般取決于植被覆蓋狀況。
1.2 樣地調查
在研究區采用完全隨機設計法設置樣地,共15個,樣地面積至少1 hm2,所選樣地均為完全郁閉純林林分(郁閉度≥0.75,優勢樹種斷面積比例≥70%)。于2015年夏季進行野外調查,在所選樣地內設置調查樣方,樣方面積為20 m×20 m,對樣方內林冠層活立木(DBH≥10 cm)進行每木檢尺;在每個樣方內采用生長錐法估測其優勢樹種年齡,以樣地內優勢樹種平均年齡作為林分年齡的保守估計;選用標準木建立樹高-胸徑曲線模型計算林分平均高。
高度低于4 m的木本植物視為灌木層(Griffiths et al.,2007;Bartels et al.,2013),包括下木、未進入林冠層的幼苗和幼樹;高度低于1.3 m的維管植物被視為草本層(Bartels et al.,2013)。將每個樣方劃分為25個4 m×4 m的網格和400個1 m×1 m的網格,隨機選取能代表此樣方灌木和草本群落結構的調查小樣方各5個,各層調查小樣方共計75個。在每一小樣方內,記錄灌木(幼樹)、草本植物種類,統計其多度,估算其蓋度(Légaré et al.,2001)。樣地概況見表1。
1.3 土壤樣品采集與測定
在每個樣方內,按照“S”形取樣法設5個土壤采樣點,分別對腐殖質層、0~20 cm表層土壤進行取樣,混合后采用四分法均勻取樣后置于冰盒中帶回實驗室,冷凍待測(Brosofske et al.,2001)。采用烘干法測定土壤含水量(Riegel et al.,1992),復合電極測定土壤pH值(Riegel et al.,1992),灼燒法測定腐殖質層和表層土壤總C(Qian et al.,2003),凱氏定氮法測定總N,氟化銨-鹽酸浸提法測定有效P(Chen et al.,2004)。記錄樣方的地形因子及土壤類型。樣方環境特征見表2。
1.4 數據分析及處理
本研究采用物種重要值(IV)反映林下灌木層和草本層物種在其相對應群落中的地位:
重要值IV=(相對密度+相對頻度+相對基蓋度)/3
林下灌木層和草本層物種多樣性采用Shannon-Weiner指數進行評價:

式中,H′為物種多樣性指數,Pi為種i的相對重要值IV(Alatalo,1981;孟祥楠等,2012)。
野外調查樣方、林下物種多樣性及環境因子數據采用Excel建庫,對數據進行初步整理。基于野外測定和分析,共得到12個生物及環境因子,包括林分因子2個:林齡(SA)、林分密度(Den);主要的土壤養分因子6個:表層土壤總碳含量(TC)、土壤總氮含量(TN)、土壤有效磷含量(SP)、腐殖質層總碳含量(HC)、腐殖質層總氮含量(HN)、腐殖質層有效磷含量(HP);地形因子4個:海拔(E)、坡位(P)、坡向(A)、坡度(S)。為便于建立環境數據矩陣,將坡位和坡向數據以等級制表示,將坡位劃分為:0.2代表下坡,0.6代表中坡,1代表上坡;根據實地樣地調查的結果,油松林天然林多分布在東北坡、北坡、西北坡、西坡4個坡向,因此將坡向劃分為:0.2代表東北坡(以正北為0°,順時針旋轉22.5°~67.5°),0.4代表北坡(0°~22.5°,337.5°~360°),0.6代表西北坡(292.5°~337.5°),0.8代表西坡(247.5°~292.5°)。
首先,對所選的6個土壤養分因子進行Pearson相關分析,根據分析結果采用PCA進行降維處理以確定土壤環境因子。然后分別以15個樣方林下灌木層、草本層物種重要值矩陣(A28×15,B44×15)和樣地生物及環境矩陣(Zx×15)為基礎進行典范對應分析(CCA)和偏分析(Partial methods),分析生物及環境因子對物種分布的影響,并采用蒙特卡羅置換檢驗(Monte Carlo permutation test)分析環境因子與物種分布之間相關關系的顯著性,在此基礎上以物種多樣性指數為響應變量、具有顯著影響作用的環境因子為解釋變量,分析環境因子對林下物種多樣性的影響。數據分析在R3.3.1下完成。

表1 油松天然林樣地概況Table 1 Summary of site characteristics and sites sampled (Range)

表2 油松天然林樣地環境特征概況Table 2 Environmental characteristics of the study stands (Range)
2 結果與分析
2.1 油松林林下植物組成
在所調查的15個樣地中,共記錄林下植物72種,分屬33科65屬(見表3),其中薔薇科、菊科、豆科合計23屬28種,占全部種數38.8%,表明這3大科植物在油松天然林林下植物多樣性中占有重要地位。
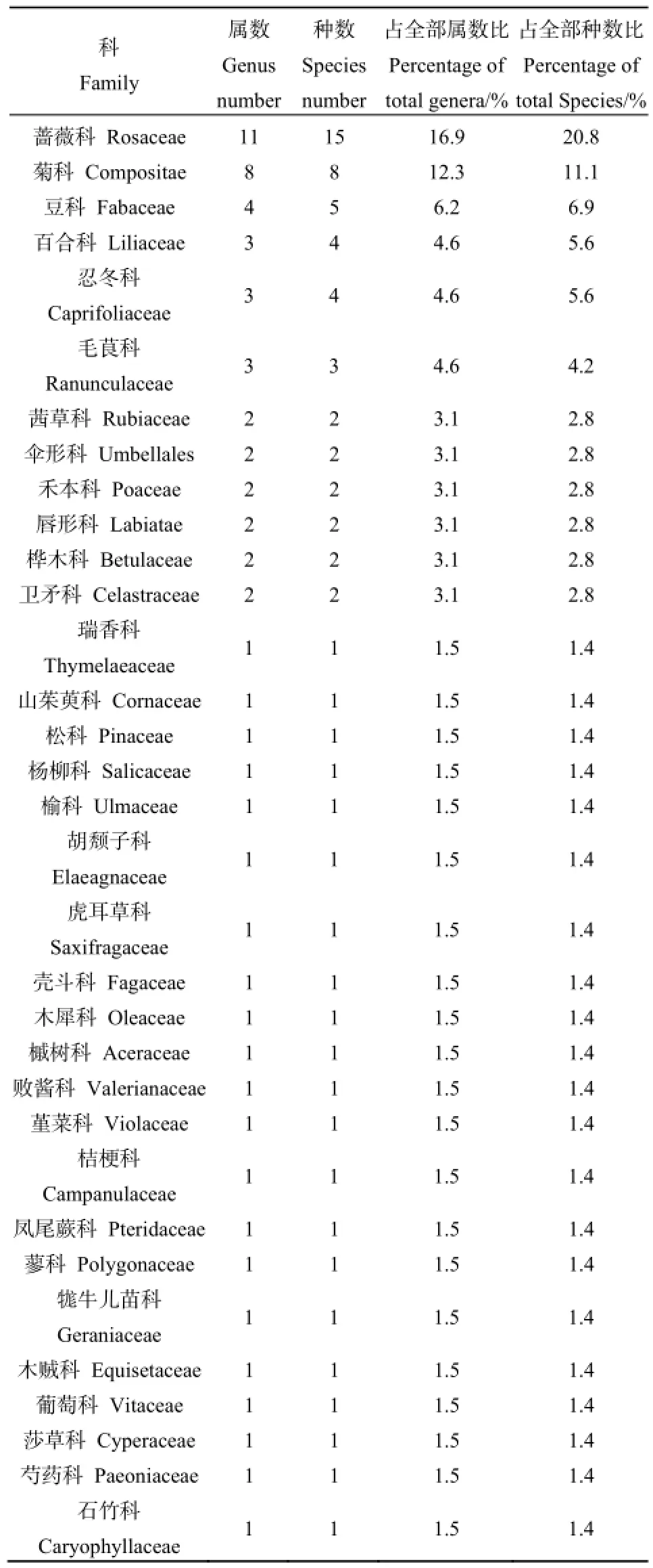
表3 油松林林下植物科屬Table 3 Family and genus distribution of species
調查樣方中林下灌木植物28種,分屬于15科24屬,其中36%的樹種生長于林緣或疏林中,18%的樹種自然分布于林下或密林中,而46%的樹種為兼性樹種。灌木層常見的植物種類有:土莊繡線菊(Spiraea pubescens Turcz.)、二色胡枝子(Lespedeza bicolor Turcz.)、虎榛子(Ostryopsis davidiana Decaisne)、水栒子(Cotoneaster multiflorus Bge.)、美薔薇(Rosa bella Rehd. et Wils)、山刺玫(Rosa davurica Pall.)等。林下草本植物44種,分屬于22科41屬,其中1、2年生的草本植物有4種,分別為早熟禾(Poa annua L.)、泥胡菜(Hemistepta lyrata Bunge)、苦苣菜(Sonchus oleraceus L.)和豬殃殃(Galium aparine Linn. var. tenerum (Gren. et Godr.) Rchb.);多年生的草本植物有40種,草本層常見的植物種類有:披針葉苔草(Carex lanceolata Boott)、早熟禾(Poa annua L.)、唐松草(Thalictrum aquilegifolium Linn. var. sibiricum Regel)、小紅菊(Dendranthema chanetii)、茜草(Rubia cordifolia L.)、柴胡(Bupleurum chinense)等。
2.2 土壤因子相關性及PCA分析
對所選的6個土壤因子進行Pearson相關性分析,結果見表4。
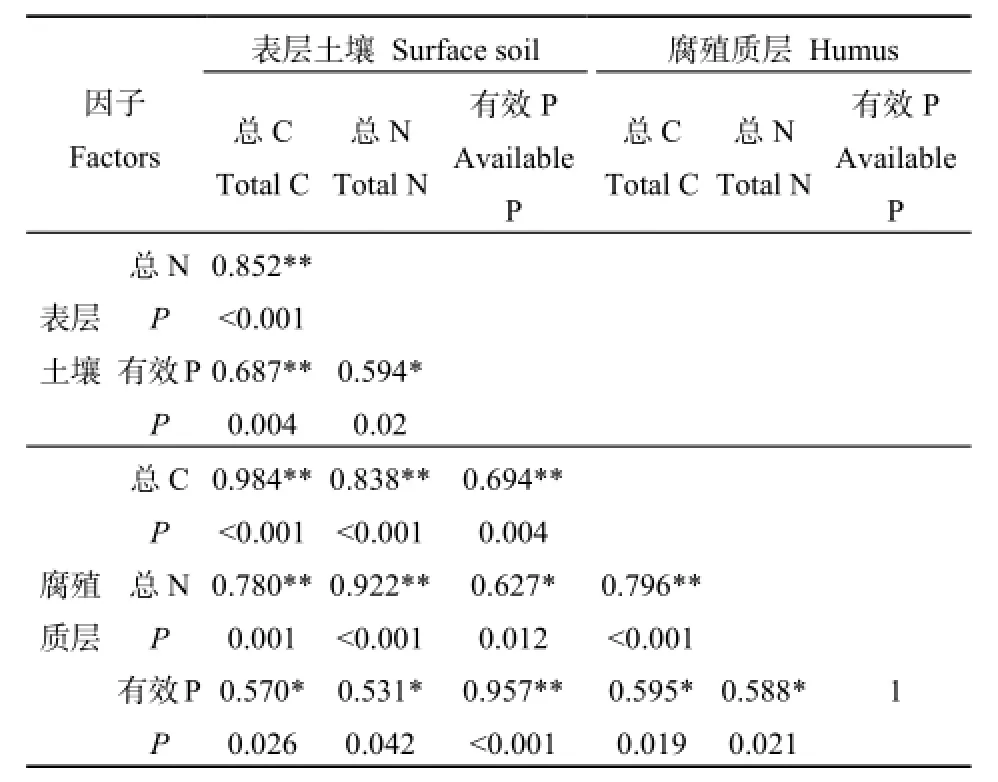
表4 土壤環境因子Pearson相關分析Table 4 Pearson correlation analysis of soil environmental factors
由Pearson相關分析可知,6個養分因子之間均呈不同水平的顯著正相關關系,進一步對這6個養分因子進行主成分分析,結果見表5和圖1。PCA分析結果顯示,前2個主分量的累積貢獻率已經達到了92%以上,其中,第一主分量的累積貢獻率為78.087%,以表層土壤總C、總N、腐殖質層總C、總N含量為主,主要反映土壤C、N含量狀況,重新命名為土壤碳氮(CN);第二主成分中主要因素為表層土壤有效P含量、腐殖質層有效P含量,主要反映土壤P含量狀況,重新命名為土壤有效磷(AP)。因此,根據PCA結果,以2個主成分值土壤碳氮(CN)、土壤有效磷(AP)代表土壤環境因子進行后續分析。
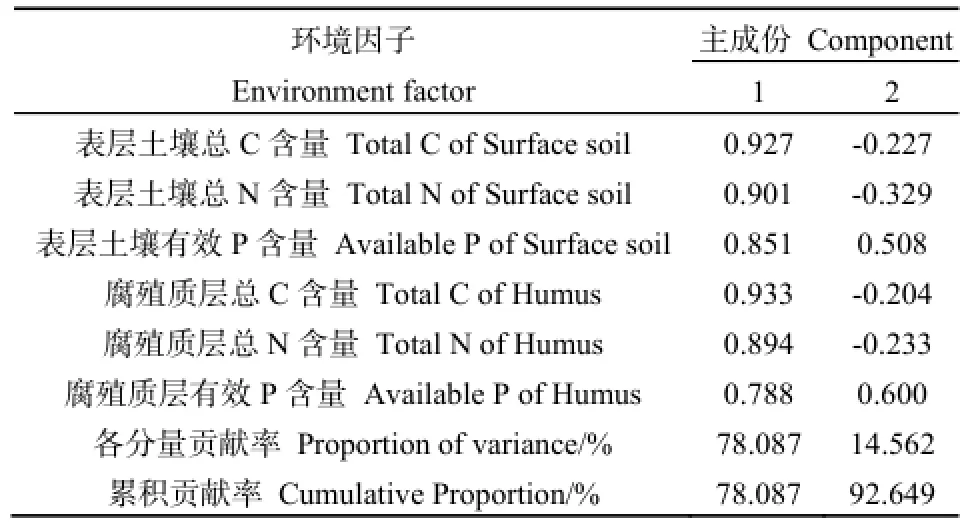
表5 不同土壤環境因子的貢獻率和主分量值Table 5 The contribution rate and principal component values of different soil environmental factors
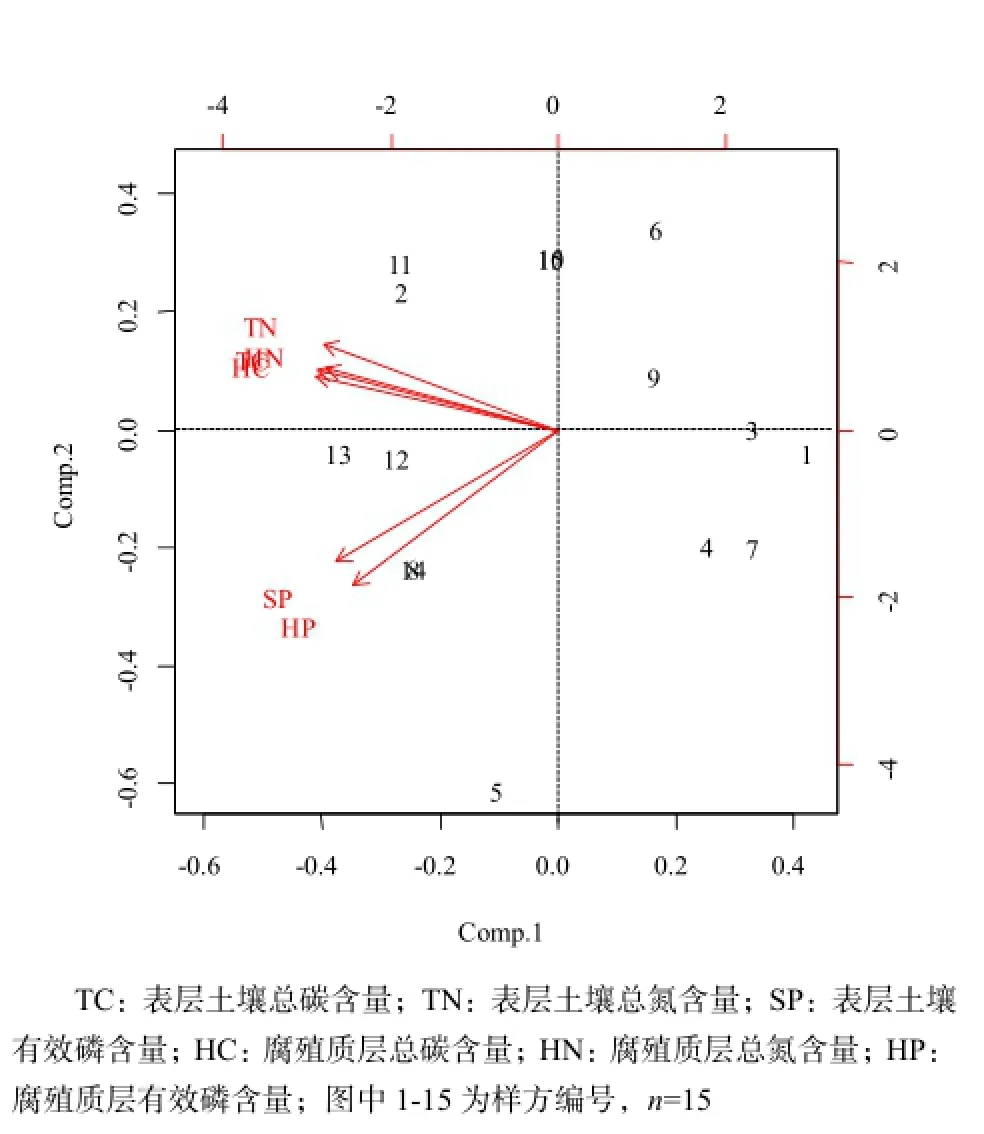
圖1 主成分圖Fig. 1 Principal component diagram
2.3 油松林林下植物與環境因子的CCA排序
基于野外調查和PCA分析,共得到8個樣地生物及環境因子,包括林分因子(2個)、土壤環境因子(2個)、地形因子(4個),建立矩陣(Z8×15),采用CCA分析確定影響該區域油松天然林下物種分布格局的主要因子。8個生物及環境因子對林下灌木層物種分布的解釋量為59.92%,采用偏分析法(Partial methods)得出林分因子即林齡、林分密度對灌木層物種分布的解釋量為16.92%,土壤環境因子即土壤碳氮含量、土壤有效磷含量對灌木層物種分布的解釋量為13.82%,地形因子即海拔、坡度、坡向、坡位的解釋量為32.44%,尚有不能解釋的物種分布變量為40.08%。根據蒙特卡羅置換檢驗,坡位(P=0.009,R2=0.57)和坡向(P=0.028,R2=0.42)是影響林下灌木層物種分布的最主要生境因子(圖2)。
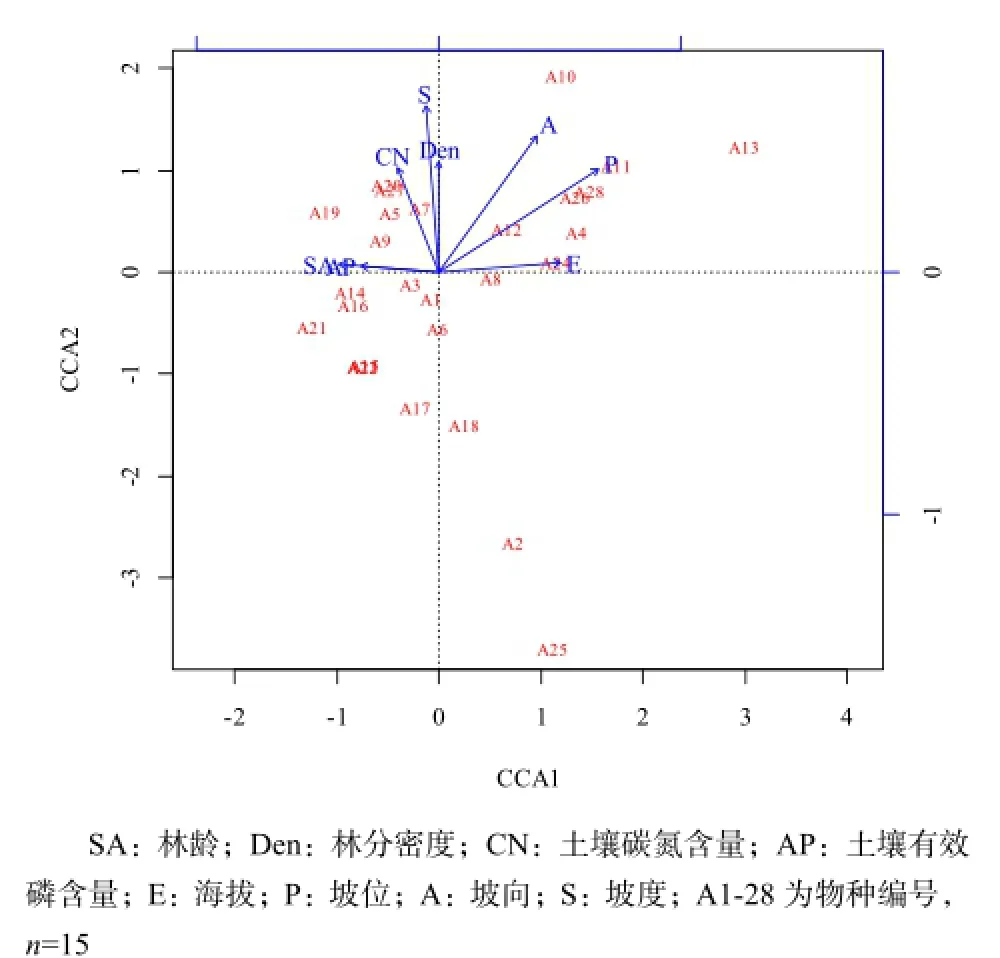
圖2 林下灌木層與環境因子的CCA二維排序Fig. 2 CCA ordination diagram of shrub layer species composition and dominant environmental factors
草本層物種組成與生物及環境因子的CCA分析結果表明,第一主成分軸和第二主成分軸總共能解釋草本層物種組成和生境因子關系變異的66.16%,其中林分因子對草本層物種分布的解釋量為16.35%,土壤因子對草本層物種分布的解釋量為13.87%,地形因子的解釋量為34.38%,3組環境變量的共同解釋量為1.56%,尚有不能解釋的物種分布變量為33.84%。根據蒙特卡羅置換檢驗,影響林下草本層物種分布的最主要生境因子是海拔(P=0.030,R2=0.43)和坡位(P=0.013,R2=0.53)(圖3)。
2.4 環境因子對物種多樣性的影響
物種重要值和多樣性相關性較為顯著,分別對坡位和坡向對林下灌木層物種多樣性、海拔和坡位對草本層物種多樣性的影響關系進行作圖(圖4)。
圖4中,林下灌木層物種多樣性在下坡最高,其次為中坡,上坡最低,隨坡向的變化表現為西北坡>東北坡>北坡>西坡;林下草本層物種多樣性隨坡位的變化與灌木層正好相反,在上坡最高、下坡最低,在所選樣方范圍內,草本層物種多樣性隨海拔的升高而升高。
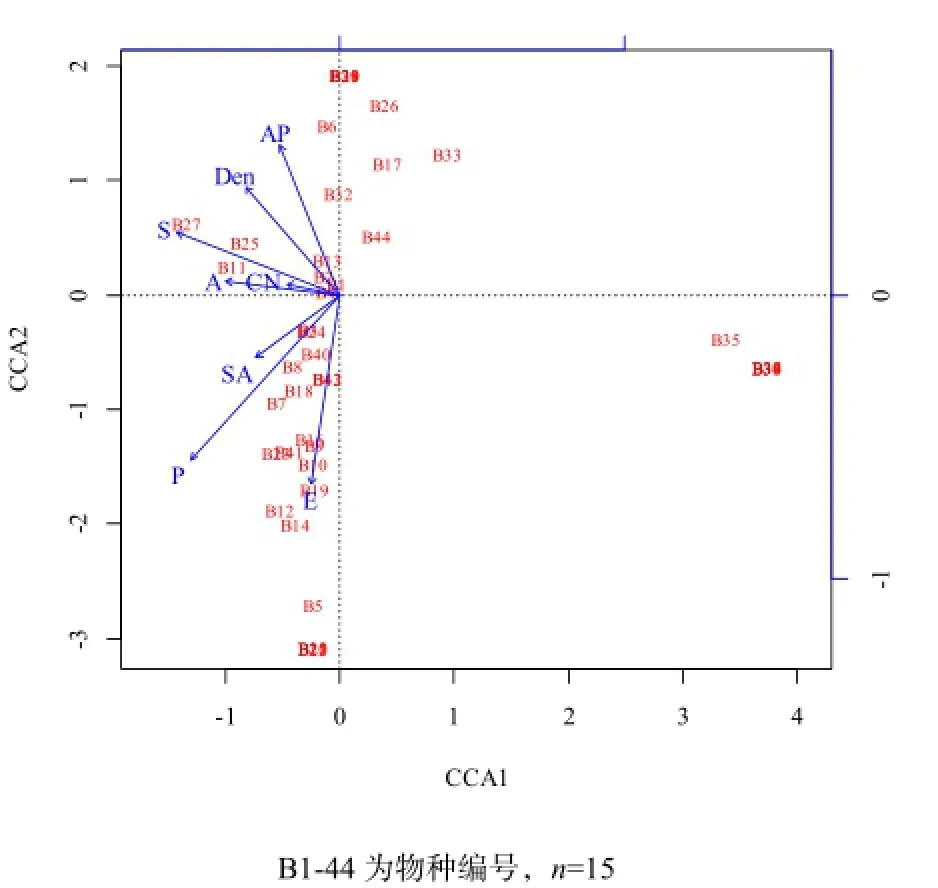
圖3 林下草本層與環境因子的CCA二維排序Fig. 3 CCA ordination diagram of herb layer species composition and dominant environmental factors
3 討論與結論
3.1 討論
在該油松天然林林下植物中,薔薇科、菊科、豆科所占比例較大,3大科植物在該地區的物種分布中占據優勢,且在油松天然林林分中也較為重要。生態型反映植物的生物學特性,36%的林下灌木生長在林緣或疏林中,而46%的樹種為兼性樹種,僅18%的樹種自然分布于林下或密林中,這和油松林的特性相關,油松林針葉直立、冠形特殊且均勻性較低(郭華等,2005;劉建等,2005),即使在郁閉度較高的林分中,光輻射也會透過林冠到達林下,這說明光照并不是油松林下灌木分布的決定性因素。而林下草本中干旱、半干旱的多年生草本占優勢,是對該區域干旱環境的高度響應,由此可以得出水分仍然是該地區草本層物種分布的限制性因素。
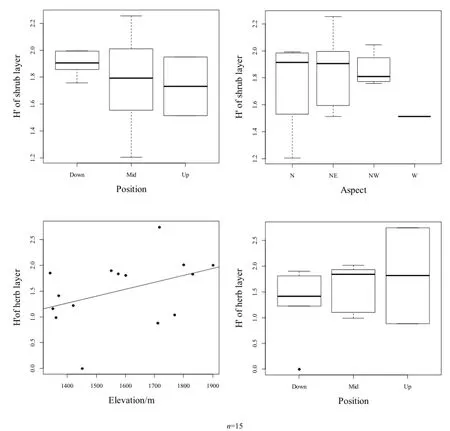
圖4 林下物種多樣性與環境顯著因子關系Fig. 4 Relationship between understory species diversity and environmental significant factors
一般認為林下植物的生物多樣性在大尺度上主要由氣候類型、海拔高度所決定,而在小尺度范圍內,則受到冠層類型、森林發育階段、坡度、坡向、坡位以及光照和土壤養分條件等多種因素的共同影響。本研究的生物及環境因子對林下灌木層和草本層多樣性分布格局的影響均在50%以上,表明所選的8個因子基本能代表林下植物物種分布的所有影響因素,尚未能解釋的因素可能是氣候條件、各種干擾以及各種因素的通過作用。在能夠解釋林下植物多樣性分布格局的環境因子中,以地形因子所占的比例略大,表明地形因子的作用大于冠層喬木(Chipman et al.,2002)和立地條件(Huo et al.,2014),是影響油松林下植物群落組成的重要環境因子,與前人的研究結果一致(Cantón et al.,2004;Maltez-Mouro et al.,2005)。
對林下植物層起主要作用的影響因子分別為坡位、坡向和海拔,坡位和坡向直接影響了太陽輻射和水肥的空間再分配,代表著光照、水分、養分等環境因素的常年、綜合生態梯度變化(劉宏文等,2014)。一般認為下坡較高的水分和養分含量會導致較高的林下植物多樣性(Chipman et al.,2002),本研究灌木物種多樣性在下坡較高說明影響其組成和分布的主要因素是水分和養分條件。在華北山地,下坡往往位于低海拔而上坡位于高海拔,而較短距離內海拔的快速變化會顯著影響地表所受到的太陽輻射量,導致微環境、季節長度、地質狀況以及人為干擾等出現較大差異(K?rner,2007),從而影響了林下植物尤其是速生的草本層的分布格局(Maltez-Mouro et al.,2005),這和前人的研究結論相似(Li et al.,2011),可能是由于高海拔地區具有較高的光照有效性引起的。
3.2 結論
在本研究調查的樣地中,關帝山油松天然林林下共出現植物物種72種,隸屬33科65屬,其中薔薇科、菊科、豆科植物種類豐富,林下灌木多為喜光或兼性樹種,林下草本以多年生草本占優勢。該區生物因子與非生物因子共同對油松天然林林下植物的分布格局產生作用,對林下灌木層和草本層的解釋率分別為59.92%和66.16%,其中地形因子的作用大于林分因子和土壤因子。林下灌木層物種多樣性在下坡最高、上坡最低,并表現為西北坡高于其他坡向;林下草本層物種多樣性隨坡位的變化與灌木層正好相反,在上坡最高、下坡最低,在所選樣方范圍內,草本層物種多樣性隨海拔的升高而升高。
ALATALO R V. 1981. Problems in the measurement of evenness in ecology [J]. Oikos, 37(2): 199-204.
BARTELS S F, CHEN H. 2013. Interactions between overstorey and understorey vegetation along an overstorey compositional gradient [J]. Journal of Vegetation Science, 24(24): 543-552.
BROSOFSKE K D, CHEN J Q, CROW T R. 2001. Understory vegetation and site factors: Implications for a managed Wisconsin landscape [J]. Forest Ecology and Management, 146(1-3): 75-87.
CANTóN Y, BARRIO G D, SOLé-BENET A, et al. 2004. Topographic controls on the spatial distribution of ground cover in the Tabernas badlands of SE Spain [J]. Catena, 55(3): 341-365.
CHASTAIN JR R A, CURRIE W S, TOWNSEND P A. 2006. Carbon sequestration and nutrient cycling implications of the evergreen understory layer in Appalachian forests [J]. Forest Ecology and Management, 231(1-3): 63-77.
CHEN H, LéGARéS, BERGERON Y. 2004. Variation of the understory composition and diversity along a gradient of productivity in Populus tremuloides stands of northern British Columbia [J]. Canadian Journal of Botany, 82(9): 1314-1323.
CHIPMAN S J, JOHNSON E A. 2002. Understory vascular plant species diversity in the mixed wood boreal forest of Western Canada [J]. Ecological Applications, 12(2): 588-601.
CHIPMAN S J, JOHNSON E A. 2002. Understory vascular plant species diversity in the mixedwood boreal forest of Western Canada [J]. Ecological Applications, 12(2): 588-601.
GAMFELDT L, SN?LL T, BAGCHI R, et al. 2013. Higher levels of multiple ecosystem services are found in forests with more tree species [J]. Nature Communications, 4: 1-8.
GRIFFITHS M E, LAWES M J, TSVUURA Z. 2007. Understorey gaps influence regeneration dynamics in subtropical coastal dune forest [J]. Plant Ecology, 189(2): 227-236.
HUO H, FENG Q, SU Y H. 2014. The Influences of Canopy Species and Topographic Variables on Understory Species Diversity and Composition in Coniferous Forests [J]. The Scientif i c World Journal, (2): 252489.
K?RNER C. 2007. The use of “altitude” in ecological research [J]. Trends in Ecology & Evolution, 22(11): 569-574.
LéGARé S, BERGERON Y, LEDUC A, et al. 2001. Comparison of the understory vegetation in boreal forest types of southwest Quebec [J]. Canadian Journal of Botany, 79(9): 1019-1027.
LI L P, WANG X P, ZERBE S, et al. 2011. Altitudinal patterns of stand structure and herb layer diversity of Picea schrenkiana forests in the central Tianshan Mountains, Northwest China [J]. Journal of Arid Land, 3(4): 254-260.
MALTEZ-MOURO S, GARCíA LV, MARA?óN T, et al. 2005. Understory floristic heterogeneity within a Mediterranean oak forest: how much is explained by edaphic, spatial and overstory components? [J]. Revista de Biologia (Lisboa), 23: 121-133.
NILSSON M C, WARDLE D A. 2005. nderstory vegetation as a forest ecosystem driver: evidence from the northern Swedish boreal forest [J]. Frontiers in Ecology and the Environment, 3(8): 421-428.
QIAN H, KLINKA K, ?KLAND R H, et al. 2003. Understorey vegetation in boreal Picea mariana and Populus tremuloides stands in British Columbia [J]. Journal of Vegetation Science, 14(2): 173-184.
RIEGEL G M, MILLER R F, KRUEGER W C. 1992. Competition forresources between understory vegetation and overstory Pinus ponderosa in northern Oregon [J]. Ecological Applications, 2(1): 71-85.
WU Y C, LI Z C, CHENG C F, et al. 2014. Responses of Soil Organic Carbon to Long-Term Understory Removal in Subtropical Cinnamomum camphora Stands [J]. International Journal of Ecology, (4): 1-6.
叢靜, 尹華群, 盧慧, 等. 2013. 神農架保護區典型植被的物種多樣性和環境解釋[J]. 林業科學, 49(5): 30-35.
郭華, 王孝安. 2005. 黃土高原子午嶺人工油松林冠層特性研究[J].西北植物學報, 25(7): 1335-1339.
劉宏文, 程小琴, 康峰峰. 2014. 油松人工林林下植物群落變化及其環境解釋[J]. 生態學雜志, 33(2): 290-295.
劉建, 何維明, 房志玲. 2005. 東靈山油松林和遼東櫟林下土壤資源和光資源的空間特征[J]. 生態學報, 25(11): 2954-2960.
孟祥楠, 趙雨森, 鄭磊, 等. 2012. 嫩江沙地不同年齡樟子松人工林種群結構與林下物種多樣性動態[J]. 應用生態學報, 23(9): 2332-2338.
尤業明, 徐佳玉, 蔡道雄, 等. 2016. 廣西憑祥不同年齡紅椎林林下植物物種多樣性及其環境解釋[J]. 生態學報, 36(1): 164-172.
Species Composition and Environmental Factors of Understory Plant Communities in Pinus tabulaeformis Garr. Natural Forest of Guandi Mountain
WANG Hui, NAN Hongwei, LIU Ning
College of Forestry, Shanxi Agricultural University, Taigu 030801, China
As one of the important components of forest ecosystem, understory plant communities plays an important role in regulation of forest structure and function. This paper studied 15 plots in Guandi mountain, north China, to explored the relationship between understory plant communities and environment factors, and revealed the key controlling factors of distribution pattern of understory plant communities in Pinus tabulaeformis natural forest.The results showed that: (1) 72 understory species belonging to 33 families and 65 genera were recorded on 15 plots, mainly in Rosaceae, Compositae and Leguminosae accounting for 38.8% in total species. Shrubs mostly are heliophile or facultative species, which had close relations with special crown-shape of Pinus tabulaeformis forest, and perennial herbs were dominant living types. (2) The results of CCA clearly reflected the relationship between the distribution of understory plants and environment factors. Biological factors and non-biological factors work together to the distribution pattern of understory plant, explained 59.92% and 66.16% of the variation in the shrub and herb layer, respectively. The effect of topographical factor was greater than stands or soil factors, plays an important role in understory plants’ composition and distribution. The species diversity of shrub layers were highest on down-slope and lowest on up-slope, northwest-wards was higher than other direction; herb layers were highest on up-slope, while lowest on down-slope and increasing along with the altitude.
mountainous regions of north China; Pinus tabulaeformis Garr. natural forest; understory plant; distribution pattern; environmental factors
10.16258/j.cnki.1674-5906.2017.01.003
Q948; X17
A
1674-5906(2017)01-0013-07
王慧, 南宏偉, 劉寧. 2017. 關帝山油松天然林林下植物組成及環境解釋[J]. 生態環境學報, 26(1): 13-19.
WANG Hui, NAN Hongwei, LIU Ning. 2017. Species composition and environmental factors of understory plant communities in Pinus tabulaeformis Garr. natural forest of Guandi Mountain [J]. Ecology and Environmental Sciences, 26(1): 13-19.
國家自然科學基金項目(31470630)
王慧(1985年生),女,講師,博士,研究方向為森林培育。E-mail: sxauwh@163.com *通信作者
2016-11-28
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
中老年保健(2021年12期)2021-08-24 03:30:40
中國傳媒大學學報(自然科學版)(2021年1期)2021-06-09 08:43:00
中國生殖健康(2020年6期)2020-02-01 06:28:50
新世紀智能(英語備考)(2019年12期)2020-01-13 06:07:18
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
中國生殖健康(2019年11期)2019-01-07 01:28:02
中國生殖健康(2018年6期)2018-11-06 07:09:28
