利用CRISPR/CAS9技術編輯水稻香味基因Badh2
2017-04-05 06:26:47邵高能謝黎虹焦桂愛魏祥進圣忠華唐紹清胡培松
中國水稻科學 2017年2期
關鍵詞:水稻
邵高能 謝黎虹 焦桂愛 魏祥進 圣忠華 唐紹清胡培松
實驗技術
利用CRISPR/CAS9技術編輯水稻香味基因Badh2
邵高能 謝黎虹 焦桂愛 魏祥進 圣忠華 唐紹清*胡培松*
(中國水稻研究所/水稻生物學國家重點實驗室/農業部水稻生物學與遺傳育種重點實驗室,杭州310006;*通訊聯系人, E-mail: sqtang@126.com, hupeisong@caas.cn)
【目的】香稻作為一類特殊的水稻群體,以其清香可口的品質特性備受消費者的歡迎。到目前為止,水稻中的香味主要受第8染色體上編碼甜菜堿醛脫氫酶基因Badh2控制。【方法】通過CRISPR/CAS9技術對中花11的香味基因Badh2進行編輯。【結果】獲得轉基因T0代植株并對其所衍生的T1代20個單株進行了鑒定分析,獲得了一個剔除了載體骨架且第1外顯子上插入一個堿基T 的突變體材料。該材料中Badh2 RNA水平顯著下調;利用GC-MS技術測定野生型及突變體材料籽粒2-乙酰-1-吡咯啉含量,結果表明突變體材料中的香味物質顯著增加;此外,我們還對野生型及T2代香型植株水稻產量及稻米蒸煮食味品質進行了考查及測定分析,發現除分蘗數及結實率呈現出顯著差異外(P<0.05),其余各項指標在兩組材料間都無顯著差異。【結論】通過CRISPR/CAS9技術成功地對水稻香味基因進行了編輯,可為香稻育種提供豐富的理論指導,加快香稻的育種進程。
水稻;Badh2;CRISPR/CAS9;香味
近些年來,大量研究報道了水稻香味基因的研究,主要包括香味基因的遺傳分析,基因的克隆以及香味基因的育種利用等[2-4]。早期研究表明香味基因主要由1~3對顯性或隱性基因控制[2,4],隨著研究的深入,遺傳分析結果表明水稻中香味主要受位于第8染色體上的一個單隱形基因Badh2控制,該基因編碼甜菜堿脫氫酶[5],包含15個外顯子和14個內含子,共編碼503個氨基酸,研究表明該基因與香味密切相關并成功克隆[5-6]。香味物質的合成也是香稻研究的重要方面,前人報道了脯氨酸和鳥氨酸可能為香味物質2-乙酰-1-吡咯啉(2-AP)的前體物質[7],當Badh2突變后導致香味物質2-AP含量的大量積累,非香稻材料中該基因所編碼的蛋白甜菜堿脫氫酶可能通過催化其底物4-氨基丁醛進而轉化為4-氨基丁酸,而香稻材料中則可能促進4-氨基丁酸轉化為1-吡咯啉,進而與乙酰基團結合,從而形成香味物質2-AP,但具體的Badh2蛋白功能及2-AP代謝途徑還沒有徹底研究清楚[6]。
分子標記輔助選擇被認為是一種有效的育種手段,特別適合于一些性狀考查比較困難的基因。香味性狀的鑒定相對比較困難,容易受主觀因素的影響,因此,香味基因分子標記輔助選擇可以大大提高其選擇的效率。Badh2第7外顯子8 bp缺失的突變體基因badh2首先被報道。此外,Chen等[5]還發現了Badh2另外一個等位基因,即第2外顯子存在7 bp的缺失,并將有功能的Badh2基因轉化到第2外顯子缺失的香稻材料中,發現轉基因植株的2-AP含量顯著下降,該研究結果表明第2外顯子上7個堿基的缺失也可引起水稻香味的產生。此外,隨著香味基因Badh2的克隆[6],一系列Badh2等位變異基因被成功鑒定,相關的功能標記也被開發利用[8-11]。近些年來研究人員希望通過生物技術手段對水稻品種中的香味基因進行編輯,從而加快育種進程。Niu等[12]通過RNAi技術,對日本晴中的香味基因BADH2進行敲除,結果發現,轉基因后代植株中的香味物質2-乙酰-1-吡咯啉(2-AP)含量明顯增加;Chen等[13]采用人工小RNA轉基因技術,對日本晴中的香味基因BADH2進行敲除,結果表明轉基因后代籽粒中2-AP含量明顯提高。轉錄激活因子效應物核酸酶(TALEN)是基因組編輯核酸酶,可以有效地對靶向基因組進行編輯,Shan等[14]通過TALEN技術,對水稻香味基因BADH2進行基因組編輯,獲得了大約30%的雜合BADH2基因型的遺傳材料,通過后代分離鑒定,獲得剔除了轉基因克隆載體的香型突變體材料,突變體材料的2-AP含量明顯提高。
本研究利用CRISPR-CAS9技術,以中花11為研究對象,對水稻香味基因Badh2進行編輯,獲得不攜帶轉基因克隆載體及表現出香味的植株,以期為加快水稻香味育種進程提供一定的理論指導。
1 材料與方法
1.1試驗材料
本研究以中花11為遺傳轉化受體,開展常規的轉基因試驗,轉基因材料種植于中國水稻研究所富陽試驗基地,常規田間種植及管理。
1.2載體構建及轉基因研究
利用試劑盒快速方便地將gRNA靶點序列插入到Cas9/gRNA質粒中。構建好的Cas9/gRNA質粒能夠同時表達植物密碼子優化的Cas9蛋白及gRNA,應用CRISPR技術進行目標基因的敲除和編輯,構建載體及具體插入位置信息如圖1所示(北京唯尚立德生物科技有限公司)。
在油脂化學領域,與脂肪酸羧基相關的反應是非常基礎和重要的,其中很多工藝都被大量的運用于日常生活當中,比如酯化、酰胺化等反應;也有一些幾乎不為人所知。所以有必要全面的加以了解,尤其是對其可發生的化學反應應該具有一定的宏觀認知,這樣有利于在創新上開闊思維,擴展相關反應在不同領域中的運用。
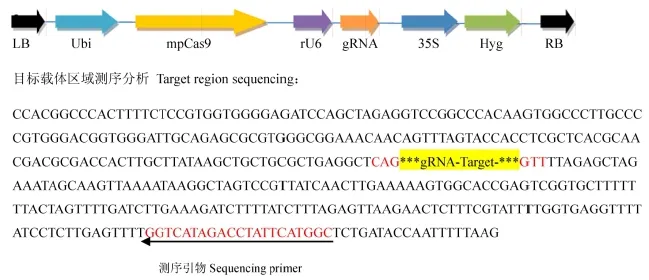
圖1 Cas9/gRNA載體圖Fig. 1. Vector map of Cas9/gRNA.
1.3香味物質測定
香味物質2-乙酰-1-吡咯啉(2-AP)含量利用氣相色譜-質譜聯用儀測定。測定過程中采用2,4,6-三甲基吡啶作為內標,將氣體成分以氦氣作為傳送媒介連續地注射到配置有光離子探測器的氣相色譜儀,氣相色譜儀與5975質量選擇檢測器連接,其中采用一個HP-5MS色譜毛細管柱氣體樣品直接作為離子源,通過電子碰撞的模式起作用,最后經由NIST2005檢測香氣物質的峰值,2-AP的出峰時間為6.324 min。該方法樣品處理簡單、快速、靈敏,樣本和試劑消耗少,能較容易地區分香稻和非香稻品種,降低了人為判斷香味的主觀性,提高了香味鑒定的準確性,特別適合大批量香稻育種材料的鑒定,具體方法參考Shan等[14],并作了相應的改進。

表1 本研究所使用的引物Table 1. List of primers used in this study.
1.4突變體表型分析
利用中花11及突變體材料,考查株高、分蘗數、結實率及每穗粒數等產量性狀。考查8個單株,獲得數據通過Excel進行統計分析;千粒重的測定方法為數300粒種子并測定其質量,8個重復。此外,還對直鏈淀粉含量,膠稠度及堿消值等三項蒸煮食味品質指標進行了測定,3次重復,獲得的數據通過Excel進行統計分析。
1.5PCR鑒定分析及qRT-PCR分析
分蘗盛期取參試材料單株葉片,采用CTAB法提取全基因組DNA,之后放置于-20℃冰箱中保存。然后將提取的DNA用于PCR擴增實驗,PCR體系如下:DNA 5 μL,正反向引物(10 μmol/L)各1.5 μL,dNTPs(2 μmol/L)10 μL,2×緩沖液 25 μL,KOD Fx酶1 μL,加ddH2O補足50 μL。PCR產物在2%瓊脂糖凝膠上進行電泳,電泳結束后EB溶液顯色并拍照,需要測序的樣品直接切膠回收送上海博尚生物技術有限公司。
利用RNA 提取試劑盒(Axygen)提取突變體和野生型葉片總RNA。 首先利用DNaseⅠ處理總RNA,接著以消化處理后的RNA為模板,采用cDNA 合成試劑盒(TOYOBO)反轉錄合成第1鏈cDNA,然后利用實時定量PCR (qRT-PCR)方法分析Badh2在野生型和突變體中的表達量,其中Ubiquitin基因作為內參基因。qRT-PCR體系如下: cDNA 模板1 μL,2×SYBR qPCR Mix (TOYOBO) 10 μL,正反引物(10 μmol/L)各1 μL,ddH2O 補足至20 μL。PCR擴增程序如下:95℃下預變性3 min,95℃下10 s,60℃下30 s,72℃下20 s,45個循環。利用公式2-ΔΔCT方法計算基因的相對表達量。
1 結果與分析
2.1CRISPR-CAS9表達載體的構建
通過對中花11編碼甜菜堿脫氫酶基因Badh2進行測序分析,發現該基因開放閱讀框ORF序列與日本晴高度一致,屬于典型的非香型粳稻材料。此外,該材料在轉基因過程中由于其轉化效率高、生育期適中、結實率較高等優點,因此,我們以中花11為轉基因受體,借助CRISPR/CAS9基因敲除技術開展轉基因實驗。首先我們通過已知的香味基因Badh2登錄號LOC_Os08g32870,獲得了其全長cDNA序列,結合http://crispr.dbcls.jp/生物信息學網站,通過序列比對分析,在Badh2外顯子上找到一條長度為20 bp特異性較好的靶點序列Target 1(表1, 圖2-A、B),將其構建到表達載體中(圖1),并開展轉基因實驗。
2.2Badh2的編輯及無標記后代的鑒定分析
首先我們獲得了8個獨立的T0代轉基因植株,我們對每個轉基因苗提取相應的DNA,同時在表達載體上設計特異性引物Vector-identify-F/R對靶點序列進行了測序驗證(表1)。測序結果表明,我們驗證了靶點序列的載體已成功插入到植物基因組,通過利用設計特異性測序引物Seq-F/R對Badh2靶點位置的基因組序列進行了測序分析(表1)。結果表明,其中3個獨立轉基因系材料在PAM附近發生了編輯,由于兩個轉基因系沒有收到足夠的種子,因此僅對其中一個轉基因系進行了遺傳研究(圖2-C)。
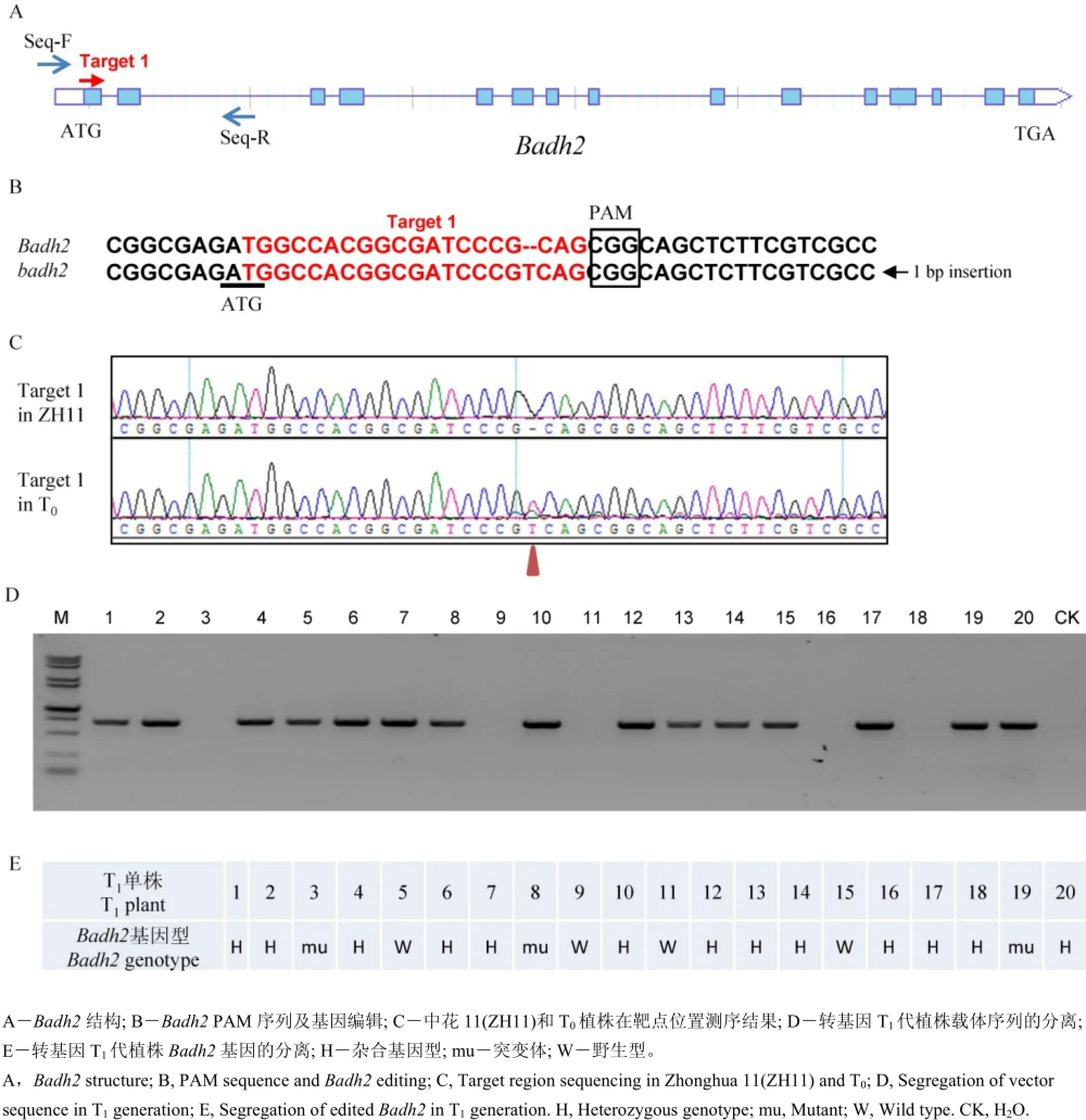
圖2 Badh2及基因編輯Fig.2. Badh2 and its CRISPR-CAS9-mediated editing.
我們將T1代材料種植到轉基因實驗田,通過DNA提取及載體序列擴增,發現載體在后代中發生了分離(圖2-D)。此外,我們進一步對T1代植株Badh2進行了測序分析,發現在Badh2第一外顯子上距離起始密碼子ATG下游18 bp位置插入一個堿基T,導致了蛋白質的移碼,影響了Badh2蛋白的結構和功能(圖2-B)。通過對T120個單株Badh2編輯位點進行測序分析,發現了該基因突變位點也發生了分離(圖2-E)。通過CRISPR-CAS9 技術對水稻中的香味基因進行編輯,希望獲得剔除了載體骨架,同時還有香味的遺傳材料,結合載體序列PCR鑒定和Badh2靶點測序結果,發現單株3中的載體骨架已經分離出去,而且還攜帶有突變形式的香味基因badh2(圖2-D、E)。
2.3轉基因后代Badh2 RNA水平檢測及香味物質2-AP含量的測定
前期我們獲得了一個轉基因材料badh2,該材料中不包含載體序列且Badh2已被成功編輯,野生型與badh2表現出相似的植株表型(圖3-A)。此外,為了進一步研究香味基因在轉基因后代中的功能,我們對中花11和突變體材料中Badh2 RNA表達水平進行了檢測。前人已報道水稻苗期葉片適合于Badh2 RNA 檢測[5],因此我們提取了兩組材料苗期14 d左右葉片總RNA,然后通過qRT-PCR技術,對Badh2進行了定量分析,我們發現突變體材料中Badh2表達水平顯著降低(圖3-B)。該結果表明香味基因第1外顯子一個核苷酸的插入,影響了該基因的轉錄水平,導致該基因在突變體材料中表達顯著下調。
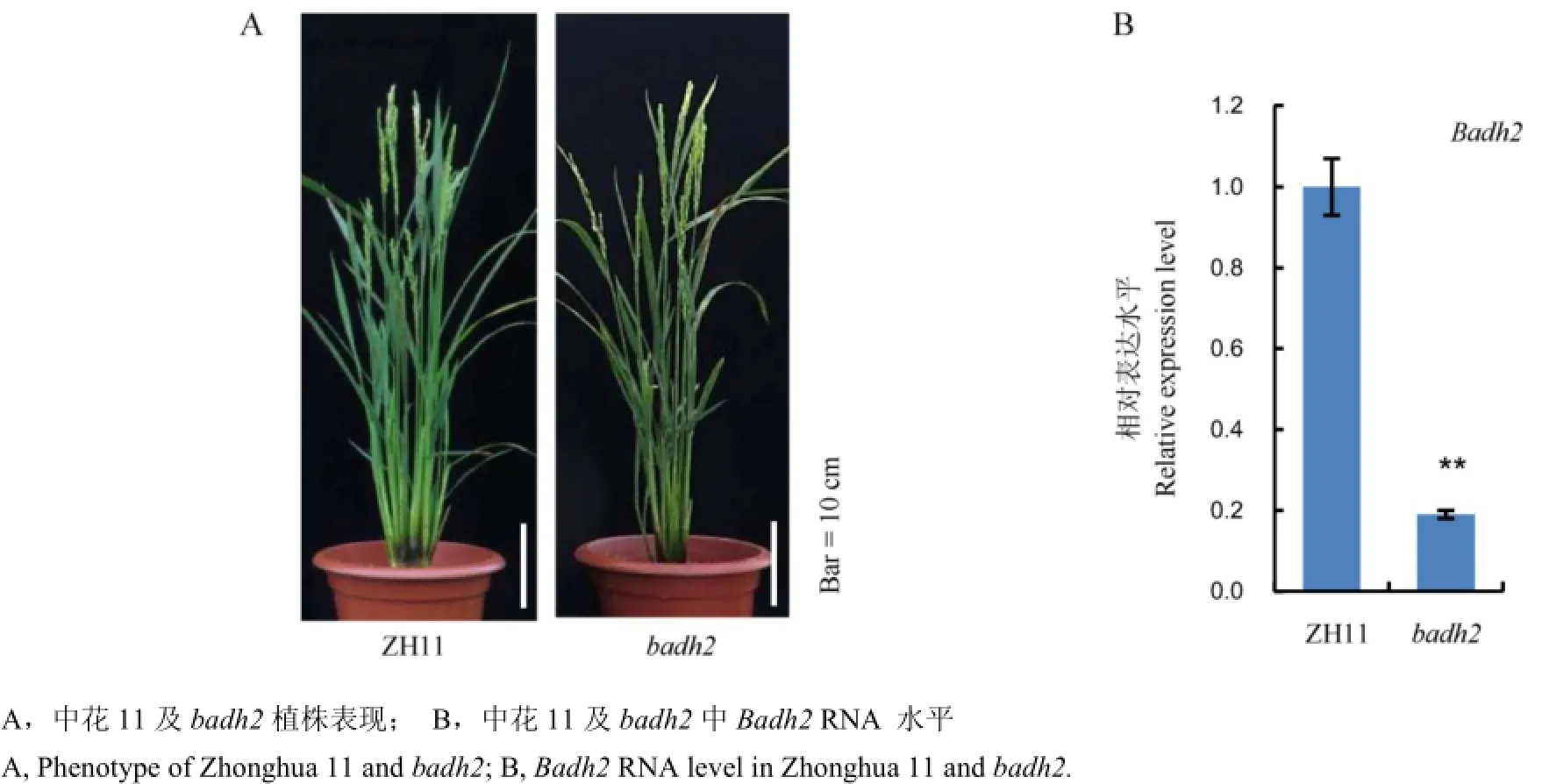
圖3 Badh2基因編輯后代表型及RNA表達水平的檢測Fig. 3. Phenotype and Badh2 RNA level of ZH11(Zhonghua 11) and badh2.
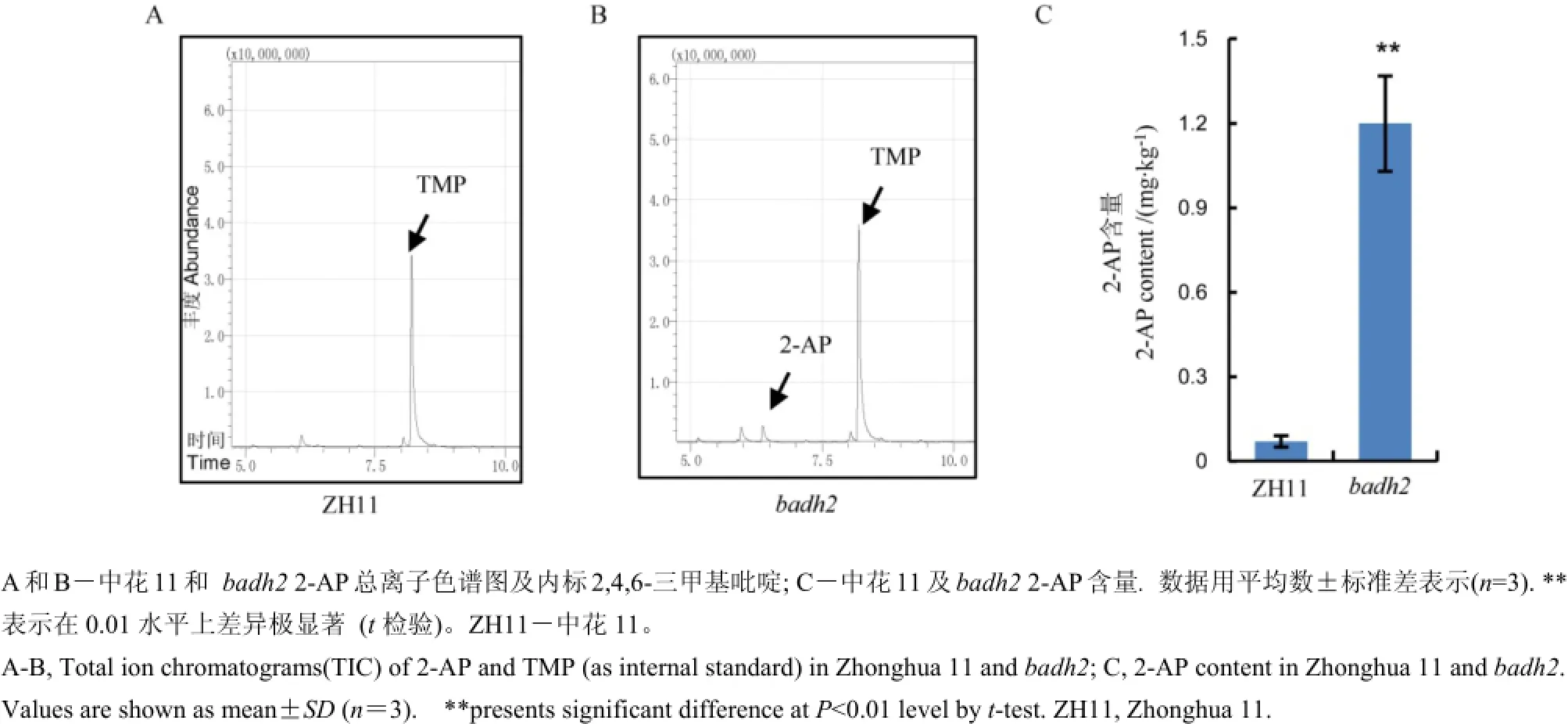
圖4 中花11及badh2中總離子色譜分析及香味物質2-AP的含量Fig. 4. Total ion chromatograms(TIC) and 2-acetyl-1-pyrroline(2-AP)content of Zhonghua 11 and badh2.
與此同時,我們進一步分析了該基因的突變及表達水平的降低是否與香味的產生相關。采用氣相色譜-質譜聯用技術對兩份材料中花11及突變體材料籽粒中的2-AP含量進行了分析(圖4-A、B),與野生型中花11對照相比,突變體材料中的香味物質2-AP顯著提高,中花11中2-AP含量約0.07mg/kg,而突變體中則增加到1.2 mg/kg(圖4-C),因此,我們的結果表明Badh2突變是引起香味的一個關鍵基因。
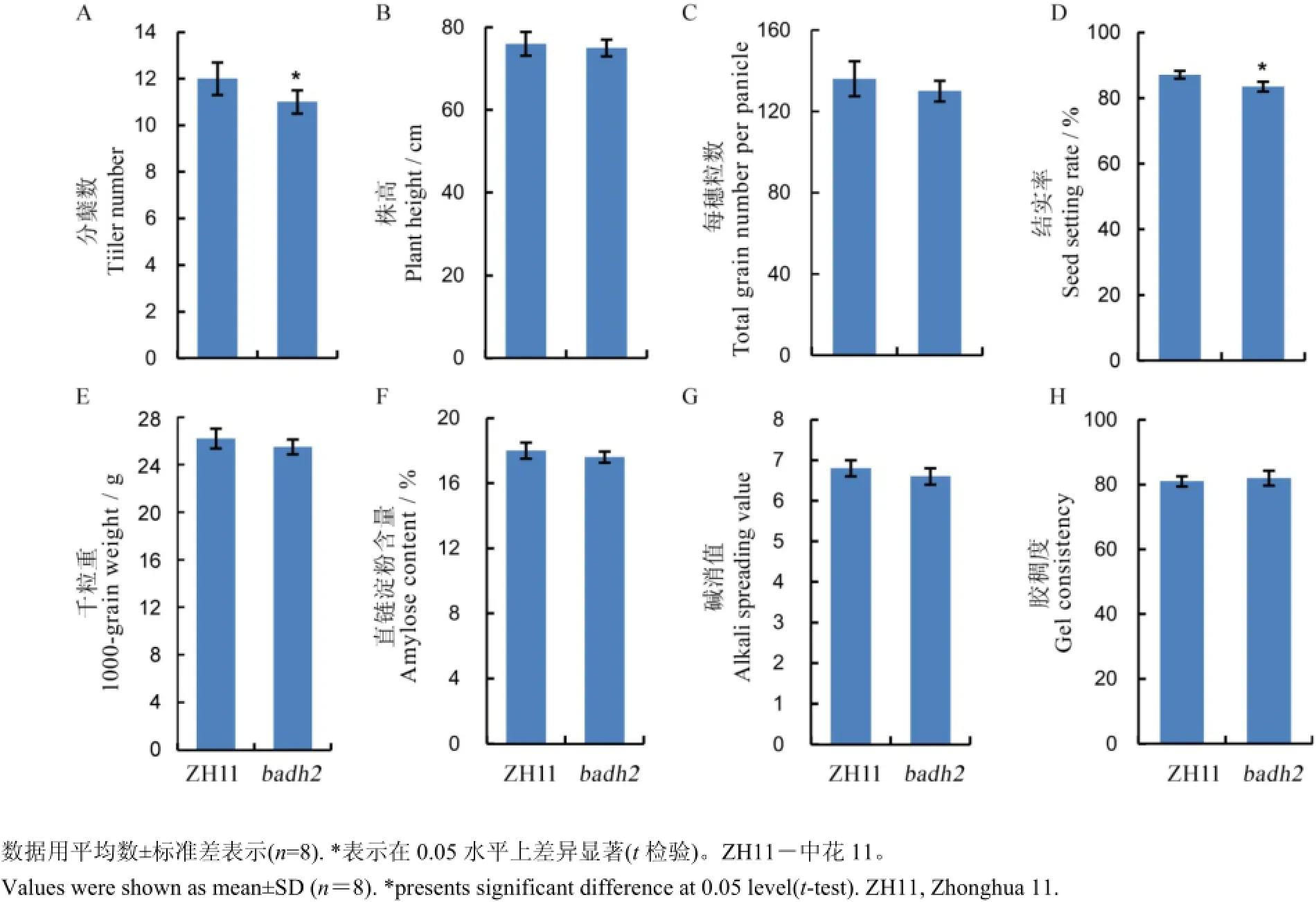
圖5 水稻產量及蒸煮食味品質在野生型及突變體材料中的表現Fig. 5. Performance of rice yield and cooking and eating quality in ZH11 and badh2.
2.4Badh2基因編輯后代產量性狀的考查及蒸煮食味品質的測定
為了研究香味基因是否與水稻產量及稻米品質密切相關,我們對中花11及其T2代突變體材料中的產量性狀及蒸煮食味品質等幾項指標進行了考查及測定。產量性狀主要包括株高、分蘗數、千粒重、每穗粒數及結實率,而蒸煮食味品質主要包括直鏈淀粉含量、膠稠度及堿消值。通過生物統計分析,我們發現除了分蘗數及結實率在兩組材料中表現出顯著差異外(P<0.05),其余各項指標間都無顯著差異(圖5)。因此,通過以上數據的分析結果,推斷該基因并不影響稻米品質,但是有可能影響水稻的產量。
3 討論
全球香稻資源豐富、栽培歷史悠久、種植范圍分布廣泛,印度的Basmati系列、泰國的KDML105、日本的宮香、美國的Della都是頗有名氣的香稻品種。我國也是香稻的主要種植區域,種植歷史悠久,各水稻主產區都有適應當地氣候的原始香稻類型,如云南的螃蟹谷、貴州的香禾、廣西有靖西香糯等。實際生產過程中,傳統香稻存在地域性強、生育期長、抗倒伏和抗病能力差、產量低等缺點,使得香稻的推廣和生產應用受到了較大的限制[15]。傳統的香稻育種主要通過香型和非香型品種間雜交,由于后代分離植株香味鑒定的不確定性,以及香味基因可能與一些產量、品質或抗病基因存在連鎖累贅,因此成功選育成香型優良品種比較困難。
眾所周知,傳統香稻育種還是主要通過傳統的育種手段,品種間雜交及后代鑒定篩選,需要花費大量時間和精力,因此,前人已嘗試通過轉基因的技術,直接對我們所培育品種中的香味基因Badh2進行敲除或編輯,從而加快育種進程[12-14]。CRISPR-CAS9系統是繼RNAi技術、鋅指核酸酶和TALEN核酸酶之后最新發展起來的另一個可精確定點編輯基因組DNA的新技術,其具有設計構建簡單快速等優點。該技術首先在動物中廣泛利用,隨后科研人員將該技術應用于植物基因組編輯。我們通過在Badh2基因ORF區域上設計特異性靶位點(圖2-A),進行轉基因特異性敲除實驗,成功獲得了一個攜帶有香味且剔除了載體序列的突變體材料badh2,該基因第一外顯子上發生了一個核苷酸T的插入(圖2-B),不僅導致Badh2 RNA水平顯著降低(圖3-B),而且引起了該蛋白質的移碼,突變后的Badh2不能發揮正常蛋白功能,進而導致了香味的產生(圖4-C),由于該材料未攜帶轉基因元件,可直接應用于育種研究,培育新的香型水稻材料,因此,利用該轉基因技術策略及特異性靶點,可以直接對不同水稻品種的Badh2進行編輯,創制香型遺傳材料,這將大大加速香稻品種的選育進程。
前人已報道香味基因與產量、品質及抗病性存在一定的關系,我們通過對野生型材料中花11及轉基因后代材料的產量及品質指標進行了統計分析,發現香味基因的變異與稻米品質沒有表現出一定的相關性,只有分蘗數和結實率兩項指標在中花11及突變體材料中表現出顯著差異,株高、千粒重、每穗粒數以及三項蒸煮食味品質指標在兩組材料都無顯著差異(圖5)。轉基因植物容易導致植株變矮,分蘗數變少及結實率降低,所以我們推測該差異可能歸因于脫靶或轉基因過程中的組織培養,因此,我們目前將對突變體材料與野生型材料進行回交,進一步研究香味基因與分蘗數及結實率的關系。
[1] 胡培松, 唐紹清, 魏興華. 泰國香米事件及啟示. 中國稻米, 2006, 12(4): 1-2.
Hu P S, Tang S Q, Wei X H. Thailand's aromatic rice incident and its revelation. China Rice, 2006, 12(4): 1-2. (in Chinese)
[2] Ahn S N, Bollich, C N, Tanksley S D. RFLP tagging of a gene for aroma in rice. Theor Appl Genet, 1992, 84:825-828.
[3] Pinson S. Inheritance of aroma in six rice cultivars. Crop Sci, 1994, 34: 1151-1157.
[4] Sakthivel K, Sundaram R M, Shobha Rani N, Balachandran S M, Neeraja C N. Genetic and molecular basis of fragrance in rice. Biotechnol Adv, 2009, 27:468-473.
[5] Chen S H, Yang Y, Shi W W, Ji Q, He F, Zhang Z D, Cheng Z K, Liu X N. Badh2, encoding betaine aldehyde dehydrogenase, inhibits the biosynthesis of 2-acetyl-1-pyrroline, a major component in rice fragrance. Plant Cell, 2008, 20: 1850-1861.
[6] Bradbury L M, Fitzgerald T L, Henry R J, Jin Q S, Waters D L. The gene for fragrance in rice. Plant Biotechnol J, 2005, 3: 363-370.
[7] Bradbury L M, Gillies S A, Brushett D J, Waters D L, Henry R J. Inactivation of an aminoaldehyde dehydrogenase is responsible for fragrance in rice. Plant Mol Biol, 2008, 68: 439-449.
[8] Kovach M J, Calingacion M N, Fitzgerald M A, McCouch S R. The origin and evolution of fragrance in rice (Oryza sativa L.). Proc Natl Acad Sci U S A, 2009, 106: 14444-14449.
[9] Shao G N, Tang S Q, Chen M L, Wei X J, He J W, Luo J, Jiao G A, Hu Y C, Xie L H, Hu P S. Haplotype variation at Badh2, the gene determining fragrance in rice. Genomics, 2013, 101: 157-162.
[10] Shao G N, Tang A, Tang S Q, Luo J, Jiao G A, Wu J L, Hu P S. A new deletion mutation of fragrant gene and the development of three molecular markers for fragrance in rice. Plant Breeding, 2011, 130: 172-176.
[11] Shi W W, Yang Y, Chen S H, Xu M L. Discovery of a new fragrance allele and the development of functional markers for the breeding of fragrant rice varieties. Mol Breed, 2008, 22: 185-192.
[12] Niu X L, Tang W, Huang W Z, Ren G J, Wang Q L, Luo D, Xiao Y Y, Yang S M, Wang F, Lu B R, Gao F Y, Lu T G, Liu Y S. RNAi-directed downregulation of OsBADH2 results in aroma (2-acetyl-1-pyrroline) production in rice (Oryza sativa L.). BMC Plant Biol, 2008, 8: 100.
[13] Chen M L, Wei X J, Shao, G N, Tang S Q, Luo J, Hu P S. Fragrance of the rice grain achieved via artificial microRNA-induced down-regulation of OsBADH2. Plant Breeding, 2012, 131: 584-590.
[14] Shan Q W, Zhang Y, Chen K L, Zhang K, Gao C X. Creation of fragrant rice by targeted knockout of the OsBADH2 gene using TALEN technology. Plant Biotechnol J, 2015, 13: 791-800.
[15] 余亞瑩, 邵高能, 圣忠華, 蔣漢偉,賀記外,孫園園,蔡怡聰,胡培松,唐紹清. 國內外香稻資源遺傳多樣性研究. 植物分類與資源學報, 2015, 37(6): 871-880.
Yu Y Y, Shao G N, Sheng Z H, Jiang H W, He J W, Sun Y Y, Cai Y C, Hu P S, Tang S Q. Genetic diversity of global aromatic rice varieties. Plant Div&Resour, 2015, 37(6):871-880. (in Chinese with English abstract)
CRISPR/CAS9-mediated Editing of the Fragrant Gene Badh2 in Rice
SHAO Gaoneng, XIE Lihong, JIAO Guiai, WEI Xiangjin, SHENG Zhonghua, TANG Shaoqing*, HU Peisong*
(State Key Laboratory of Rice Biology, Key Laboratory of Rice Biology and Breeding of Ministry of Agriculture, China National Rice Research Institute, Hangzhou 310006, China;*Corresponding author, E-mail: sqtang@126.com, hupeisong@caas.cn)
【Objective】Fragrant rice, which is favored because of its quality characteristics of faint scent and tastiness, is a special rice type. Fragrance in rice is mainly controlled by the gene encoding betaine aldehyde dehydrogenase on chromosome 8.【Method】The fragrant gene Badh2 in Zhonghua 11 was edited by CRISPR/CAS9.【Result】Twenty T1individuals derived from T0generation were genotyped. One plant, which contains an additional T base in the first exon of Badh2 without the vector skeleton, was finally produced. qRT-PCR result suggested that Badh2 RNA level was decreased in the mutant. Compared with the wild type, the mutant increased 2-acetyl-1-pyrroline content by the GC-MS method. Furthermore, two rice yield-related traits including tiller numbers and seed-setting rate showed significant difference at the 0.05 level among five traits related to yield and three traits related to cooking and eating quality.【Conclusion】We succeed editing the Badh2 by CRISPR-CAS9 technical in rice, and it would provide abundant theoretical guidance and accelerate the breeding process of fragrant rice.
rice; Badh2; CRISPR/CAS9; fragrance

Q755; S511.032
A
1001-7216(2017)02-0216-07
2016-06-20; 修改稿收到日期:2016-09-10。
國家自然科學基金國際(地區)合作與交流項目(31161140348); 浙江省自然科學基金資助項目(Y14C130040)。
猜你喜歡
幼兒100(2023年39期)2023-10-23 11:36:32
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
中國土壤與肥料(2021年5期)2021-12-12 02:02:11
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-10-14 08:35:40
金橋(2021年7期)2021-07-22 01:55:38
今日農業(2020年20期)2020-11-26 06:09:10
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
