烏梁素海濕地蘆葦最大羧化速率的高光譜遙感
2017-04-10 12:28:09衛亞星王莉雯
生態學報 2017年3期
衛亞星,王莉雯
1 遼寧師范大學海洋經濟與可持續發展研究中心,大連 116029 2 遼寧師范大學自然地理與空間信息科學遼寧省重點實驗室,大連 116029 3 遼寧師范大學城市與環境學院,大連 116029
烏梁素海濕地蘆葦最大羧化速率的高光譜遙感
衛亞星1,2,3,*,王莉雯1,2,3
1 遼寧師范大學海洋經濟與可持續發展研究中心,大連 116029 2 遼寧師范大學自然地理與空間信息科學遼寧省重點實驗室,大連 116029 3 遼寧師范大學城市與環境學院,大連 116029
濕地植被生產力和固碳潛力的研究是全球碳循環和全球變化的熱點研究問題。濕地植被的光合能力能夠指示其生長的健康狀態。最大羧化速率是重要的植被光合參數之一,對精確模擬濕地植被光合作用和氣體交換模型中的固碳過程具有重要的作用。以內蒙古烏梁素海湖泊濕地為研究區,進行了蘆葦葉片光合參數和光譜的測量。蘆葦葉片最大羧化速率(Vcmax)數值是基于Farquhar光合作用模型,從光合測量獲取的A-Ci曲線計算并校正到25 ℃得到的。分別基于bootstrap PLSR模型、單波段和高光譜植被指數(包括簡單比值指數SR和歸一化差值指數ND),構建濕地蘆葦葉片最大羧化速率(Vcmax)估算模型。基于高光譜遙感圖像HJ- 1A HSI,采用ND高光譜指數中具有較高Vcmax估算精度的入選波段702和756 nm,獲取研究區濕地蘆葦最大羧化速率空間分布圖。研究結果表明,濕地植被光譜特征和高光譜植被指數,可用于估算濕地蘆葦Vcmax,其中最高精度產生于基于bootstrap PLSR模型的建模方法(R2=0.87,RMSECV=3.90,RPD=2.72),ND高光譜指數的Vcmax估算精度高于SR高光譜指數的估算精度;從獲取的Vcmax空間分布圖上提取估算值,其與測量值對比,存在較好的相關性(R2=0.80,RMSE=4.74)。
最大羧化速率;高光譜遙感;濕地植被光合;bootstrap PLSR;烏梁素海濕地
陸地生態系統碳循環過程與碳源、碳匯問題的研究已成為20世紀90年代以來科技界的最大熱點之一。近20年來國內外已經發展了大量的陸地生態系統模型,其中光合作用生化模型是陸地生態機理模型的核心。這些模型多以植被葉片最大羧化速率作為關鍵參數進行光合作用的模擬[1]。一些經典生化模型在模擬植被光合作用時,通常將最大羧化速率設置為常數,或僅考慮了溫度的影響,但沒有考慮其它環境影響因素,以及由此導致的植被最大羧化速率的時空變化。植物最大羧化速率的空間差異和季相變化的準確模擬,為精確評估陸地生態系統生產力、碳收支及對氣候變化的響應提供依據[2]。
最大羧化速率(Vcmax)是重要的植被光合生理參數,是植被光合作用過程中重要限速反應——羧化反應的速率,對光合速率起著決定性的作用,它決定了植物的最大凈光合速率、光下線粒體呼吸、光呼吸等過程[3]。植被葉片最大羧化速率是植被光合作用過程中由核酮糖- 1,5-二磷酸羧化酶/加氧酶(Rubisco)催化的最大羧化反應速率,即植被葉片在單位時間單位面積固定的最大CO2摩爾數。不同類型植被葉片的Vcmax差異較大,其值在6—194μmol m-2s-1之間變化。同一類型植被葉片的Vcmax在不同生長環境下也不相同,并且呈現年際和季相變化特征[4]。
基于植被葉片或冠層尺度的光譜特征,采用遙感分析技術可以估算植被生化、生理和結構特性參數[5- 8]。采用遙感反演技術,有助于在不同尺度上獲取植被特定光譜特征與最大羧化速率的響應關系,由此準確估算植被光合作用關鍵參數的時空變化。
隨著對植被光合作用和生產力的深入研究,一些研究對影響植物光合和呼吸作用的Vcmax及其影響因子進行了觀測和模擬。其中,采用光譜分析技術模擬Vcmax空間和時相變化的研究也開始出現。Dillen等觀測了兩個樹種在生長季葉片反射率與光合參數(包括Vcmax等參數)的季相變化[9]。研究了紅邊位置指數與光合參數的相關關系,認為光譜植被指數具有指示關鍵光合參數生長季變化的潛力。Jin等的研究表明,不同尺度獲取的植被指數與Vcmax存在緊密相關性,所建立的相關關系可以用于反演光合參數的季相變化[10]。Serbin等基于葉片反射光譜對Vcmax估算進行了研究,發現光譜反射率數據可以有效捕捉短期山楊樹葉片Vcmax對溫度變化的響應[11]。Gamon等對PRI指數進行了研究,他們從研究結果推論出,光合參數會受到同等的調節作用,PRI指數可用于探測植被Vcmax和氣孔導度的變化[12]。雖然有關植被Vcmax的光譜反演研究還較少,但Vcmax對植被生理生態的重要性,會促使植被Vcmax光譜研究成為一個值得探索的研究方向。
本文以干旱區湖泊濕地蘆葦作為研究對象,采用高光譜遙感技術,探索濕地植被主要光合參數最大羧化速率高光譜估算技術,并且模擬其空間分布狀況,為準確模擬濕地生態系統碳同化和碳收支,評估濕地植被群落健康狀態和濕地凈化能力提供科學依據。
1 研究區概況
烏梁素海濕地位于內蒙古自治區西部巴彥淖爾市烏拉特前旗境內,地理坐標為東經108°43′—108°57′,北緯40°36′—41°03′,是河套地區最大的湖泊濕地。烏梁素海系黃河改道后形成的牛軛湖,湖區水域面積約為293 km2,是干旱區典型的大型草型淡水湖泊。烏梁素海濕地區域屬于溫帶大陸性干旱氣候,年平均氣溫7.3℃,年平均降水量220mm。烏梁素海濕地植物以蘆葦、香蒲、蓖齒眼子菜為優勢種。蘆葦生長茂密,成大片或帶狀分布于湖中。近年來,隨著人類活動干擾強度的不斷增加,烏梁素海濕地出現了水量逐年減少和水體污染較為嚴重的問題,各項富營養化指標也呈現逐年上升的趨勢[13]。
2 數據來源和預處理
2014年7月在烏梁素海濕地進行了野外實驗。根據研究區蘆葦群落的分布特征,選定了73個采樣區域(圖1),在選定的73個采樣區域內設定單位面積為0.5 m×0.5 m的蘆葦生長樣方。野外實驗選擇晴朗無云的天氣進行,測量時間控制在10:00—12:00之間,以避開可能發生的“光合午休”。在每個樣方內隨機選擇5株長勢良好且基本一致的蘆葦植株,待測葉片選擇完全展開、長勢一致、對應葉位一致的3片成熟葉片。每片葉片分別進行3次光合參數測量和10次光譜測量。計算每個樣方所測葉片光合參數的平均值作為該樣方蘆葦葉片光合參數測量結果,計算每個樣方所測葉片光譜的平均值作為該樣方葉片反射光譜曲線,以此消除偶然誤差。
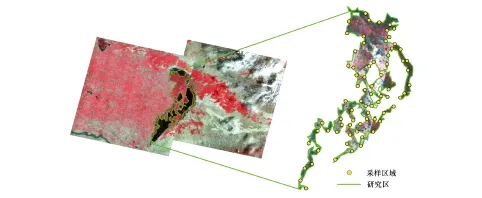
圖1 烏梁素海濕地研究區和采樣區域位置圖Fig.1 Locations of the study area and experiment sites in the Wuliangsuhai wetland
2.1 蘆葦光合參數測量和計算
使用LI- 6400便攜式光合系統分析儀(Li-cor,Lincoln,NE,USA),進行了蘆葦葉片光合參數、氣體交換參數和環境參數的測量,包括葉片凈光合速率、氣孔導度、蒸騰速率、胞間CO2濃度、葉面溫度、大氣溫度、相對空氣濕度、空氣CO2濃度和光合有效輻射等。
測定光響應曲線時,控制CO2的濃度為400 μmolCO2mol-1,設定光合強度梯度為0、50、100、150、200、400、650、800、1000、1200、1400、1600、1800、2000 μmol m-2s-1。測定CO2響應曲線時,控制光合強度為650 μmol m-2s-1,設定CO2濃度梯度為0、50、100、150、200、400、600、800、1000、1200、1400、1600、1800、2000 μmol mol-1。采用二次曲線方程,分別進行光響應曲線擬合和CO2響應曲線擬合,繪制光合(A-PAR)響應曲線和光合-CO2(A-Ci)響應曲線(A為測量葉片的凈光合速率,PAR為光合有效輻射,Ci為CO2濃度),計算表1所列其它光合參數。
基于Farquhar等提出的生化模型和A-Ci響應曲線[14],采用非線性回歸方法,擬合Vcmax數值[10]:
式中,Γ*為缺乏暗呼吸下的CO2補償點;KC和KO分別為羧化作用和加氧作用的Michaelis-Menten常數。在A-Ci響應曲線初始部分,即Ci低值區間,A主要受Vcmax的限制。因此,Vcmax和光下的暗呼吸速率(Rd*),基于Ci<250 μmol mol-1的部分A-Ci曲線進行估算。在缺乏Rd*(Γ*)下的CO2補償點和Michaelis-Menten常數,采用Brooks等[15]提出的葉溫轉換方法和Von Caemmerer等[16]的算法進行估算:
式中,O和Ci分別是胞間O2分壓和胞間CO2分壓;τ是Rubisco特異性因子;Rd為在光下的線粒體呼吸速率。隨后,基于Harley等提出的溫度響應算法[17],將計算獲得的Vcmax校正到25 ℃:
式中,Ha為活化能;Hd為鈍化能;c為尺度轉換常數;ΔS為熵期,取值為0.65kJ K-1mol-1;R為氣體常數;Tk為葉溫。
2.2 蘆葦葉片光譜測量
蘆葦葉片光譜測量使用了ASD FieldSpec Pro FR野外光譜輻射儀。光譜測量范圍為350—2500 nm。傳感器探頭采取垂直向下的位置進行觀測,測量視場角為25°,距離所測蘆葦葉片上方約1 cm處。實驗中使用了99%的朗伯體白板作為參考板。
2.3 高光譜圖像
環境與災害監測預報衛星HJ- 1A上搭載有超光譜成像儀(hyperspectral imaging radiometer,HSI)傳感器。其空間分辨率為100 m,工作光譜譜段115個,光譜范圍459—956 nm。相比于EO- 1 Hyperion、MODIS等傳感器,其光譜監測性能有所提高,從而有效提高了對地物信息的提取能力,適用于植被生長狀況、生化組分信息等多種專題研究[18]。
研究采用了2014年7月覆蓋研究區的HSI高光譜圖像,軌道號為11/65和12/65。對高光譜圖像依次進行了格式轉換、絕對輻亮度值轉換、大氣校正和幾何精校正等數據預處理。采用ENVI的FLAASH模型(fast line-of-sight atmospheric analysis of spectral hypercubes)進行圖像的大氣校正。以TM為參考影像,使用二次多項式法進行了圖像的幾何精校正,校正誤差控制在0.5個像元,得到歸一化地表反射率圖像。
3 研究方法
3.1 高光譜參量選取
本文基于單波段和高光譜植被指數,進行濕地蘆葦光合參數最大羧化速率的回歸分析。單波段光譜采用ASD光譜輻射儀測量獲取的高光譜數據(400—2350 nm波譜范圍)。高光譜植被指數采用了兩種基本類型的指數,分別為簡單比值指數(SR)和歸一化差值指數(ND):
(1)
(2)
式中,Rλ1為波段λ1的反射率,Rλ2為波段λ2的反射率,λ1≠λ2。在本研究中,在400—2350nm波譜范圍內(去除波段除外),采用了由此波譜區間內任意兩個波段構建的SR和ND高光譜指數。
3.2 基于bootstrap的PLSR模型
本研究采用了bootstrap技術結合偏最小二乘回歸模型(PLSR)的回歸建模方法。
bootstrap技術是利用有限的樣本進行隨機、等概率、有放回的多次重復抽樣,建立起足以代表母體樣本分布之新樣本,從而實現擴大樣本容量進行數據模擬。因此,基于再抽樣和數據模擬的bootstrap技術,能夠有效提高所建模型的估算精度[19]。
本研究在MATLAB平臺上完成建模的編程計算。具體建模步驟包括:基于原始樣本數據采用PLSR回歸建模方法構建估算模型,獲得初始回歸系數集;在原始數據集中隨機抽取一個樣本點(yi(1),xi1(1),xi2(1),…,xip(1)),記錄其值后再放回原始數據集,重復n次,得到一個bootstrap樣本:
(3)
x是自變量數據,y是因變量數據;基于bootstrap樣本S(1),采用PLSR回歸建模方法,建立估算模型:
(4)
β是回歸系數;重復(3)、(4)步驟,得到估算模型回歸系數組,構建基于bootstrap樣本的回歸系數集;計算基于bootstrap樣本的回歸系數與初始回歸系數差值的絕對值,取絕對值的90%分位點作為拒絕域的臨界值;比較回歸系數的絕對值與臨界值,判斷其是否通過顯著性檢驗;將經過顯著性檢驗的自變量在原數據集上重新構建PLSR估算模型;重復上述步驟,直到所有變量均通過顯著性檢驗。
3.3 模型建立及精度驗證
研究采用4種方法構建濕地蘆葦葉片最大羧化速率估算模型:(1)采用線性回歸建立單波段與蘆葦葉片Vcmax的回歸模型;(2)采用線性回歸建立SR高光譜指數與蘆葦葉片Vcmax的回歸模型;(3)采用線性回歸建立ND高光譜指數與蘆葦葉片Vcmax的回歸模型;(4)采用基于bootstrap PLSR模型建立預處理后光譜與蘆葦葉片Vcmax的回歸模型。
所建模型估算精度評價指標選用了R2(決定系數)、RMSECV(交叉驗證均方根誤差)和RPD。研究基于留一交叉算法(leave-one-out)計算RMSECV。即對有N個樣本的原始數據,每個步驟取出其中的1個樣本,只使用其余N-1個樣本建立預測模型,用以預測被取出的樣本值。這個過程一直重復至每個樣本都被取出1次。本文計算標準差(SD)與RMSECV的比值作為RPD值。
3.4 基于高光譜圖像的濕地蘆葦最大羧化速率空間分布估算
濕地植被最大羧化速率空間分布的獲取,有利于較為精確地描述其空間變化狀況,改進區域或全球尺度濕地生態系統生產力與碳收支模型中光合參數的精度。
在本文使用的4種方法中,選取一種適宜的濕地蘆葦最大羧化速率估算方法,將該方法應用于HSI高光譜圖像。從經過預處理的HSI圖像上,提取73個采樣區域對應像素的反射光譜曲線。將73組樣本數據(包括從圖像上提取的反射光譜曲線和測量計算獲取的Vcmax),隨機分成2組,包括建模樣本數據(n=50)和驗證樣本數據(n=23)。基于建模樣本數據,構建蘆葦Vcmax估算模型,獲取研究區濕地蘆葦最大羧化速率空間分布圖。基于驗證樣本數據,采用R2和RMSE對Vcmax估算精度進行評價。
4 結果與分析
4.1 研究區濕地蘆葦光合生理參數特征
野外實驗測量在7月進行,研究區蘆葦長勢較好,種群密度92株/m2,平均株高2.68 m,葉面積指數3.64 m2/m2。從表1可以看出,蘆葦光飽和點和光補償點之間的光能利用區間較寬,這導致蘆葦具有較高的生產力。蘆葦葉片較高的光飽和點和光補償點,也從一方面表明了其屬于不耐蔭的陽性植物[20]。從各光合生理參數指標綜合來看,研究區濕地蘆葦呈現較強的光合能力。
4.2 濕地蘆葦葉片光譜特征
圖2為經實驗測量獲取的蘆葦葉片平均反射光譜曲線。曲線中剔除了水分吸收帶、大氣影響以及噪聲干擾較大的部分。曲線呈現出綠色植物葉片的一般光譜特征:在可見光波段,在葉綠素等色素吸收和反射作用影響下,葉片光譜反射率值較低,在542 nm存在明顯的反射峰,在683 nm存在明顯的吸收谷;在近紅外波段,683—760 nm波段范圍呈現“紅邊”特征,760—1335 nm波段之間光譜反射率值較高,在1063 nm達到峰值,這是葉片細胞結構高反射率影響所致;在短波紅外(SWIR)波段,受葉片含水量的影響,光譜反射率值降低,以1450、1950 nm為中心形成了水分吸收帶。

表1 烏梁素海濕地蘆葦葉片光合生理參數
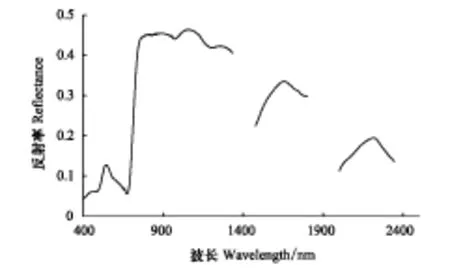
圖2 濕地蘆葦葉片平均反射光譜曲線 Fig.2 The spectral curve of the average reflectance for reed leaves in wetlands
4.3 濕地蘆葦葉片最大羧化速率估算精度對比
表2為本文中采用的4種方法估算蘆葦葉片Vcmax的結果及精度評價指標對比。圖3為采用上述4種方法構建蘆葦葉片Vcmax估算模型,入選較高估算精度的波段或波段組合,進行Vcmax估算值與測量值比較。總體來看,最高估算精度產生于基于bootstrap PLSR模型的Vcmax估算建模方法(R2=0.87,RMSECV=3.90,RPD=2.72)。ND高光譜指數的Vcmax估算精度(R2介于0.85—0.79),高于SR高光譜指數的估算精度(R2介于0.80—0.74)。基于單波段構建的Vcmax線性估算模型精度最低(R2介于0.55—0.48)。
RPD常被用來評價預測模型的精度。一些研究根據估算結果總結出,RPD值越大預測模型的估算能力越好。當RPD>2時表示模型具有較好的預測能力;2>RPD>1.4時表示模型的估算結果可以被接受,存在改進空間;RPD<1.4時說明模型預測的可靠性在減小;RPD<1時表示模型預測能力較差,不能應用于樣本估算[21]。在本研究中,基于bootstrap PLSR模型、ND高光譜指數、SR高光譜指數中的657和794波段組合(表2),其RPD值均超過2,說明這些估算方法可以獲得精度較高的蘆葦葉片Vcmax預測值。
本研究中所采用的bootstrap技術結合PLSR回歸建模方法,獲取了蘆葦葉片Vcmax的最高估算精度。 在本文中,采用了基于bootstrap的PLSR回歸建模方法,利用bootstrap技術可以有效提高估算模型精度,證明bootstrap和PLSR結合的建模方法,適用于基于高光譜數據的蘆葦葉片Vcmax估算。但是,存在的問題是,在類似生態和地理參數光譜估算研究中,通常使用整個光譜測量波段數據(包括上千個波段),這導致在PLSR模型中,部分光譜變量對估算變量不具有重要性影響(例如,影響系數接近于0)或者負影響[11]。當空間尺度由局地測量上升至寬尺度遙感應用時,可利用的波段數量較為有限,需要較多成分變量的PLSR建模方法將導致較大估算誤差。因此,基于寬波段星載光譜數據時,PLSR并不是理想的建模方法,其應用會受到該問題的限制,而光譜指數的應用卻更為普遍。
表2 4種方法估算烏梁素海濕地蘆葦葉片最大羧化速率結果和評價指標對比
Table 2 The results of estimating maximum carboxylation rate of reed leaves in Wuliangsuhai wetland using 4 methods and the comparisons of evaluation indicators (n=73)
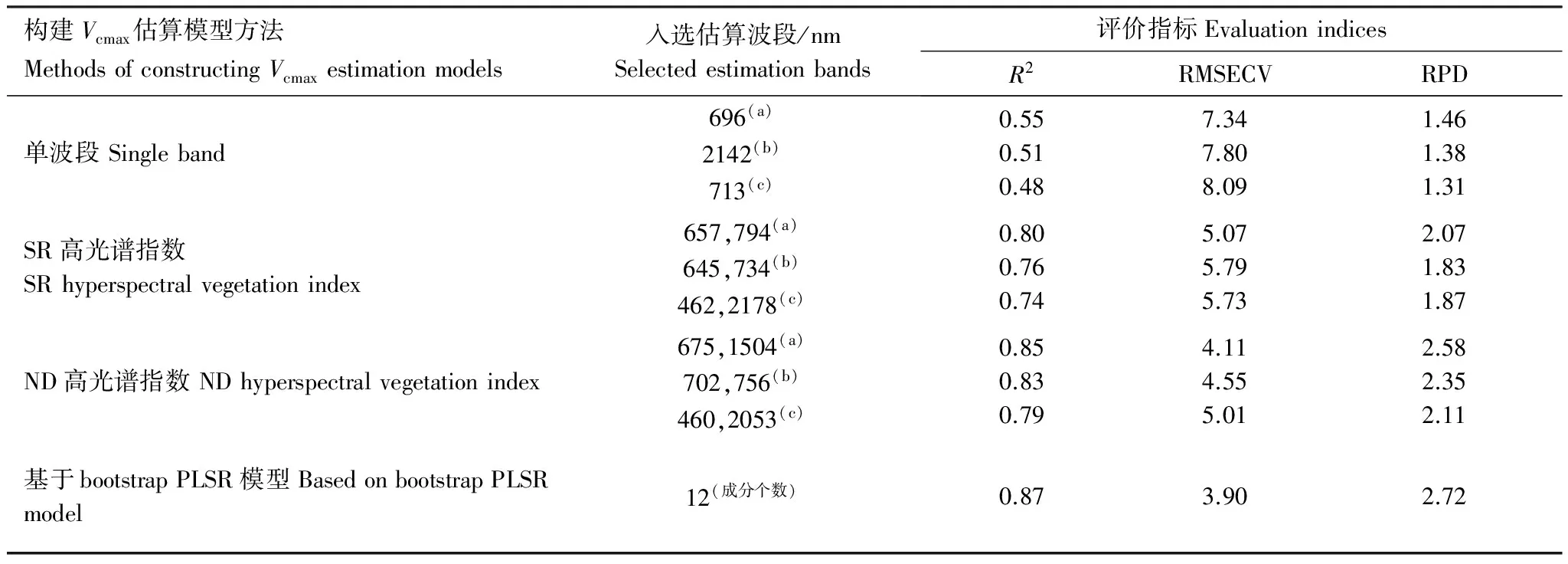
構建Vcmax估算模型方法MethodsofconstructingVcmaxestimationmodels入選估算波段/nmSelectedestimationbands評價指標EvaluationindicesR2RMSECVRPD單波段Singleband696(a)2142(b)713(c)0.550.510.487.347.808.091.461.381.31SR高光譜指數SRhyperspectralvegetationindex657,794(a)645,734(b)462,2178(c)0.800.760.745.075.795.732.071.831.87ND高光譜指數NDhyperspectralvegetationindex675,1504(a)702,756(b)460,2053(c)0.850.830.794.114.555.012.582.352.11基于bootstrapPLSR模型BasedonbootstrapPLSRmodel12(成分個數)0.873.902.72
入選估算波段一列,下標a、b、c分別代表在同一種估算蘆葦葉片Vcmax的方法中,綜合各估算精度評價指標,并且考慮實際應用中特征波段經常發生偏移,以及星載波段的設置,a為估算精度最高的入選波段,b和c為估算精度較高的入選波段;R2: 決定系數 determination coefficient; RMSECV: 交叉驗證均方根誤差 root mean square error of cross-validation; RPD: the ratio of performance to deviation
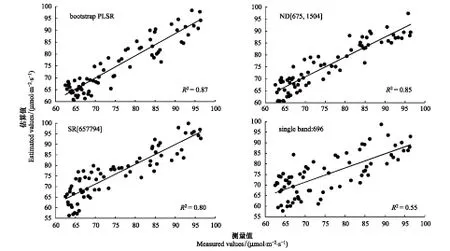
圖3 4種方法對蘆葦葉片Vcmax最高精度的估算值與測量值比較Fig.3 Measured versus estimated Vcmax with the highest accuracy for reed leaves based on four kinds of estimation methods (n=73)
歸一化植被指數(NDVI[705, 750])、光化學植被指數(PRI[531, 570])等,在已有研究中證明了其估算植被光合參數的可行性[22]。本研究中,采用了ND形式和SR形式的高光譜指數,估算濕地蘆葦葉片Vcmax。值得注意的是,ND形式中標記為b的高光譜指數,利用了702和756 nm波段,這與上文提及的705和750 nm波段較為接近,這兩個波段被認為與植被葉片葉綠素含量相關性較高,而葉綠素含量在很大程度上決定了葉片的光合能力[23]。尤其是在星載光譜數據中,相近的這兩對波段可能各自位于同一衛星波段中。探索高光譜指數對植被光合參數(例如,最大羧化速率、最大光合電子傳遞速率等)的估算研究,有利于在不同尺度上開展其時空變化監測及對環境影響因素的響應。
表2顯示,入選估算波段分別位于藍光(460、462 nm)、紅光(645、657、675、696 nm)、近紅外(702、713、734、756、794 nm)、SWIR波段(1504、2053、2142、2178 nm)。其中,696、702、713、734、756 nm位于近紅外波段的“紅邊”區域。可以看出,主要入選估算波段位于紅外、近紅外、SWIR光譜范圍。一些研究證明,許多中心位于紅光和近紅外的波段在植被葉片和冠層光合參數的指示方面具有潛能[24]。表2中,ND[702, 756]、SR[657, 794]和SR[645, 734]即為此類高光譜指數,它們表現出了對蘆葦葉片Vcmax較好的估算能力。在本研究中,一些SWIR波段也顯示出能夠指示蘆葦葉片的Vcmax。Rubisco在SWIR波段存在幾個較寬的光譜吸收特征波段,主要位于1500、1680、1740、1940、2050、2170、2290、2470 nm波段[25]。另一方面,葉片中氮素的30%—50%分配于蛋白質核酮糖- 1,5-二磷酸羧化酶/加氧酶(Rubisco)中,葉片氮素狀態與凈光合速率顯著相關[25]。而氮素在SWIR波段的吸收波段中心位置,主要位于1510、1940、2060、2180、2300、2350 nm[5]。將Rubisco、氮素吸收特征波段和表2中入選估算波段相對比,在SWIR波段,入選的1504、2053、2178 nm與前兩者相應的吸收特征波段較為接近。一些研究表明,SWIR波段的入選有利于提高預測模型的估算精度,主要源于其獨特的光譜特性[5],這在本文中也有所體現。
4.4 研究區濕地蘆葦最大羧化速率空間分布
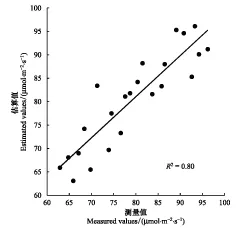
圖4 基于高光譜圖像HSI的濕地蘆葦最大羧化速率估算值與測量值比較(n=23)Fig.4 Measured versus estimated maximum carboxylation rate of wetland reeds based on HSI hyperspectral image
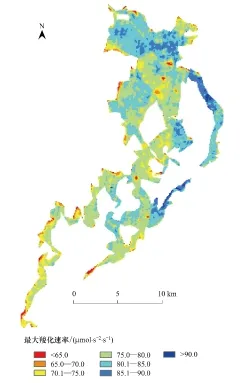
圖5 研究區濕地蘆葦最大羧化速率空間分布 Fig.5 The spatial distribution of reed maximum carboxylation rate in the wetland of study area
本文采用的高光譜遙感圖像為HJ- 1A HSI,其光譜范圍為459—956 nm。根據該高光譜圖像的波譜覆蓋范圍,以及實際推廣應用的可行性和實用性,選擇ND高光譜指數中具有較高Vcmax估算精度的入選波段702和756 nm,即采用ND[702, 756]高光譜指數估算蘆葦Vcmax。
圖4為根據23組驗證樣本數據的空間位置,從研究區濕地蘆葦最大羧化速率空間分布圖(圖5)上提取的估算值與測量值對比。圖4顯示,估算值與測量值之間存在較好的相關性(R2=0.80,RMSE=4.74)。結果表明,高光譜指數ND[702, 756]以較好的精度估算了濕地蘆葦最大羧化速率的變化,即基于光譜特征的植被指數可以較為精確地估算濕地植被最大羧化速率。
圖5為基于高光譜指數ND[702, 756]、50組建模樣本數據和線性回歸模型,獲取的研究區濕地蘆葦最大羧化速率空間分布圖。濕地蘆葦最大羧化速率估算值介于62.3—94.8之間,平均值為76.7 μmol m-2s-1。在烏梁素海濕地的研究區,北部和東部沿岸地區呈現蘆葦最大羧化速率的高值區。在這些區域,蘆葦生長密集度大,而且單個植株長勢旺盛。
5 結論
本文主要研究結論為:
(1)基于蘆葦葉片測量光譜和高光譜遙感圖像HSI,所獲取的植被光譜特征和高光譜植被指數,可用于以較高精度估算濕地蘆葦最大羧化速率,從而獲得濕地植被最大羧化速率的空間分布。
(2)用于估算濕地蘆葦葉片Vcmax的4種方法中,最高估算精度產生于基于bootstrap PLSR模型的Vcmax估算建模方法(R2=0.87,RMSECV=3.90,RPD=2.72)。ND高光譜指數的Vcmax估算精度高于SR高光譜指數的估算精度。單波段的Vcmax估算精度最低。
(3)基于高光譜遙感圖像HSI,采用ND[702, 756]高光譜指數估算蘆葦Vcmax,獲取了研究區濕地蘆葦最大羧化速率空間分布圖。經驗證,估算值與測量值之間存在較好的相關性(R2=0.80,RMSE=4.74)。
(4)估算濕地蘆葦葉片Vcmax的入選估算波段,主要位于藍光(460、462 nm)、紅光(645、657、675、696 nm)、近紅外(702、713、734、756、794 nm)、SWIR波段(1504、2053、2142、2178 nm)。其中,696、702、713、734、756 nm位于近紅外波段的“紅邊”區域。
(5)對比Rubisco、氮素吸收特征波段和本文獲得的Vcmax入選估算波段,在SWIR波段,入選的1504、2053、2178 nm與前兩者相應的吸收特征波段較為接近。
植被最大羧化速率是表征植被光合能力的重要光合參數。植被光合作用過程不僅與植物自身的生理生態特性(包括葉片構造、葉齡、葉位等)有密切關系,還受到環境因素的影響(包括光照、溫度、水分、CO2濃度、土壤養分等)。隨著遙感空間技術的發展,已存在的多種傳感器可以在不同空間和時間尺度上持續提供海量的地物光譜信息,如此大數據的光譜數據有助于精確捕捉植被光合參數的時空變化。
[1] Houborg R, Cescatti A, Migliavacca M, Kustas W P. Satellite retrievals of leaf chlorophyll and photosynthetic capacity for improved modeling of GPP. Agricultural and Forest Meteorology, 2013, 177: 10- 23.
[2] 閆霜, 張黎, 景元書, 何洪林, 于貴瑞. 植物葉片最大羧化速率與葉氮含量關系的變異性. 植物生態學報, 2014, 38(6): 640- 652.
[3] 張彥敏, 周廣勝. 植物葉片最大羧化速率對多因子響應的模擬. 科學通報, 2012, 57(13): 1112- 1118.
[4] Wullschleger S D. Biochemical limitations to carbon assimilation in C3plants-a retrospective analysis of theA/Cicurves from 109 species. Journal of Experimental Botany, 1993, 44(5): 907- 920.
[5] 王莉雯, 衛亞星. 植被氮素濃度高光譜遙感反演研究進展. 光譜學與光譜分析, 2013, 33(10): 2823- 2827.
[6] Soudani K, Hmimina G, Dufrêne E, Berveiller D, Delpierre N, Ourcival J M, Rambal S, Joffre R. Relationships between photochemical reflectance index and light-use efficiency in deciduous and evergreen broadleaf forests. Remote Sensing of Environment, 2014, 144: 73- 84.
[7] Wu C Y, Niu Z, Gao S. The potential of the satellite derived green chlorophyll index for estimating midday light use efficiency in maize, coniferous forest and grassland. Ecological Indicators, 2012, 14(1): 66- 73.
[8] Raymond Hunt Jr E, Doraiswamy P C, McMurtrey J E, Daughtry C S T, Perry E M, Akhmedov B. A visible band index for remote sensing leaf chlorophyll content at the canopy scale. International Journal of Applied Earth Observation and Geoinformation, 2013, 21: 103- 112.
[9] Dillen S Y, de Beeck M O, Hufkens K, Buonanduci M, Phillips N G. Seasonal patterns of foliar reflectance in relation to photosynthetic capacity and color index in two co-occurring tree species,QuercusrubraandBetulapapyrifera. Agricultural and Forest Meteorology, 2012, 160: 60- 68.
[10] Jin P B, Wang Q, Iio A, Tenhunen J. Retrieval of seasonal variation in photosynthetic capacity from multi-source vegetation indices. Ecological Informatics, 2012, 7(1): 7- 18.
[11] Serbin S P, Dillaway D N, Kruger E L, Townsend P A. Leaf optical properties reflect variation in photosynthetic metabolism and its sensitivity to temperature. Journal of Experimental Botany, 2012, 63(1): 489- 502.
[12] Gamon J A, Bond B. Effects of irradiance and photosynthetic downregulation on the photochemical reflectance index in Douglas-fir and ponderosa pine. Remote Sensing of Environment, 2013, 135: 141- 149.
[13] 孫惠民, 何江, 呂昌偉, 高興東, 樊慶云, 薛紅喜. 烏梁素海氮污染及其空間分布格局. 地理研究, 2006, 25(6): 1003- 1012.
[14] Farquhar G D, Von Caemmerer S, Berry J A. A biochemical model of photosynthetic CO2assimilation in leaves of C3species. Planta, 1980, 149(1): 78- 90.
[15] Brooks A, Farquhar G D. Effect of temperature on the CO2/O2specificity of ribulose- 1, 5-bisphosphate carboxylase/oxygenase and the rate of respiration in the light. Planta, 1985, 165(3): 397- 406.
[16] Von Caemmerer S, Evans J R, Hudson G S, Andrews T J. The kinetics of ribulose- 1, 5-bisphosphate carboxylase/oxygenase in vivo inferred from measurements of photosynthesis in leaves of transgenic tobacco. Planta, 1994, 195(1): 88- 97.
[17] Harley P C, Tenhunen J D. Modeling the photosynthetic response of C3leaves to environmental factors // Boote K J, Loomis R S, eds. Modeling Crop Photosynthesis-from Biochemistry to Canopy. Geneva: American Society of Agronomy, 1991: 17- 39.
[18] 陳雪洋, 蒙繼華, 吳炳方, 朱建軍, 杜鑫. 基于HJ星高光譜數據紅邊參數的冬小麥葉面積指數反演. 中國科學: 信息科學, 2011, 41(增刊): 213- 220.
[19] 王惠文, 吳載斌, 孟潔. 偏最小二乘回歸的線性與非線性方法. 北京: 國防工業出版社, 2006: 267- 274.
[20] 付為國, 李萍萍, 卞新民, 吳沿友, 曹秋玉. 鎮江北固山濕地蘆葦光合日變化的研究. 西北植物學報, 2006, 26(3): 496- 501.
[21] Anne N J P, Abd-Elrahman A H, Lewis D B, Hewitt N A. Modeling soil parameters using hyperspectral image reflectance in subtropical coastal wetlands. International Journal of Applied Earth Observation and Geoinformation, 2014, 33: 47- 56.
[22] 王莉雯, 衛亞星. 植被光能利用率高光譜遙感反演研究進展. 測繪與空間地理信息, 2015, 38(6): 15- 22.
[23] Wu C Y, Chen J M, Desai A R, Hollinger D Y, Altaf Arain M, Margolis H A, Gough C M, Staebler R M. Remote sensing of canopy light use efficiency in temperate and boreal forests of North America using MODIS imagery. Remote Sensing of Environment, 2012, 118: 60- 72.
[24] Inoue Y, Peuelas J, Miyata A, Mano M. Normalized difference spectral indices for estimating photosynthetic efficiency and capacity at a canopy scale derived from hyperspectral and CO2flux measurements in rice. Remote Sensing of Environment, 2008, 112(1): 156- 172.
[25] Elvidge C D. Visible and near infrared reflectance characteristics of dry plant materials. International Journal of Remote Sensing, 1990, 11(10): 1775- 1795.
Maximum carboxylation rates of reed in the Wuliangsuhai wetland using hyperspectral remote sensing
WEI Yaxing1,2,3,*, WANG Liwen1,2,3
1CenterforStudiesofMarineEconomyandSustainableDevelopment,LiaoningNormalUniversity,Dalian116029,China2LiaoningKeyLaboratoryofPhysicalGeographyandGeomatics,LiaoningNormalUniversity,Dalian116029,China3CollegeofUrbanandEnvironmentalScience,LiaoningNormalUniversity,Dalian116029,China
Studies on wetland vegetation productivity and their carbon sequestration potential are becoming an important focus of the global carbon cycle and global climate change research. The photosynthetic capacity of wetland vegetation can indicate the health status of its growth. In addition, an accurate estimate of maximum carboxylation rate (Vcmax) is important for accurately simulating wetland vegetation photosynthesis and carbon sequestration processes with a gas exchange model. Here, the wetland of Wuliangsuhai (Inner Mongolia) was chosen as the study area, and the photosynthetic parameters and spectral reflectance of reed leaves were measured. Based on the Farquhar model of photosynthesis, reed leafVcmaxvalues were calculated fromA-Cicurves, and subsequently standardized to 25 ℃. Estimation models ofVcmaxfor reed leaves in the wetland were constructed with a bootstrap PLSR model and single band and hyperspectral vegetation indices (e.g., simple ratio index (SR) and normalized difference index (ND)). Based on hyperspectral remote sensing images from HJ- 1A HSI, the bands of 702 and 756 nm, which had a higher estimation accuracy forVcmax, were selected from the ND hyperspectral indices. Subsequently, a spatial distribution map ofVcmaxfor wetland reed was acquired for the study area. The results showed that the spectral characteristics of wetland vegetation, combined with hyperspectral vegetation indices, could be used effectively to accurately estimate reedVcmaxin the wetland. The highest accuracy was produced from the modeling method based on a bootstrap PLSR model (R2=0.87,RMSECV=3.90,RPD=2.72). Furthermore, the accuracy ofVcmaxestimations from the ND hyperspectral indices was higher than that from the SR hyperspectral indices. Overall, the estimated values extracted from the spatial distribution map ofVcmaxhad a good correlation with the measured values (R2=0.80,RMSE=4.74).
maximum carboxylation rate; hyperspectral remote sensing; wetland vegetation photosynthesis; bootstrap PLSR; Wuliangsuhai wetland
國家自然科學基金項目(41271421);教育部人文社會科學研究規劃基金項目(14YJA630064)
2015- 09- 11;
日期:2016- 06- 13
10.5846/stxb201509111879
*通訊作者Corresponding author.E-mail: wyx9585@sina.com
衛亞星,王莉雯.烏梁素海濕地蘆葦最大羧化速率的高光譜遙感.生態學報,2017,37(3):841- 850.
Wei Y X, Wang L W.Maximum carboxylation rates of reed in the Wuliangsuhai wetland using hyperspectral remote sensing.Acta Ecologica Sinica,2017,37(3):841- 850.
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50
數學物理學報(2020年2期)2020-06-02 11:29:24
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
汽車工程學報(2017年2期)2017-07-05 08:13:02
光學精密工程(2016年6期)2016-11-07 09:07:19
