木荷皂苷對稻瘟病病原細胞的致毒作用
2017-04-15 10:32:32陳緒濤柴兆元霍光華彭玉萌陳明輝
江蘇農業科學 2017年5期
陳緒濤+柴兆元++霍光華+彭玉萌+陳明輝+黃建平+李時貴

摘要:為了探索木荷皂苷抗稻瘟病菌的作用方式,通過顯微觀察、血球計數板計數、活體侵染等方法,測定木荷5種皂苷對稻瘟病菌孢子和附著胞的致毒作用。結果顯示,木荷皂苷對稻瘟病菌孢子及附著胞的抑制作用均強于陽性對照稻瘟靈,R2-A、R2-1、R2-2、R2-3、R3-4、稻瘟靈對稻瘟病病菌的孢子萌發抑制EC50分別為2.73、2.27、0.61、0.95、4.68、13.94 μg/mL;在濃度為10 μg/mL時,它們對孢子產附著胞的抑制率分別為78.68%、93.27%、97.23%、91.93%、36.34%、9.11%;抑制稻瘟病病菌附著胞產黑色素的MIC分別為7.0、6.0、4.0、6.5、17.0、18.0 μg/mL。受處理的稻瘟病病菌孢子侵染活體水稻葉片能力會不同程度下降,同時抑制附著胞黑色素的積聚成為空胞,難以形成高壓滲透釘,阻斷了稻瘟病病菌的侵染傳播。研究表明,木荷皂苷抗稻瘟病病菌對稻瘟病病原不同類型細胞不具選擇性,除了作用于菌絲細胞膜外,還可通過抑制孢子萌發、附著胞黑色素積累等方式產生作用;R2-1、R2-2、R2-3是木荷抗稻瘟病病菌的主要活性先導物。
關鍵詞:木荷皂苷;稻瘟病病菌;孢子;附著胞;作用方式
中圖分類號: S435.111.4+1文獻標志碼: A
文章編號:1002-1302(2017)05-0086-05
木荷(Schima superba Gardn. et Champ)為茶科木荷屬植物,是我國中部到南部常綠闊葉林、常綠闊葉-落葉混交林的優勢樹種或群落上層的共建樹種,在我國除自然分布外,還被廣泛種植。木荷可作為優質用材、美麗觀賞樹、防火植被帶樹種,根皮和葉還可入藥,具攻毒消腫、殺蟲催吐的作用,主治疔瘡、無名腫毒等,樹皮甲醇提取物對小菜蛾、菜青蟲均具有極強的拒食活性[1]。
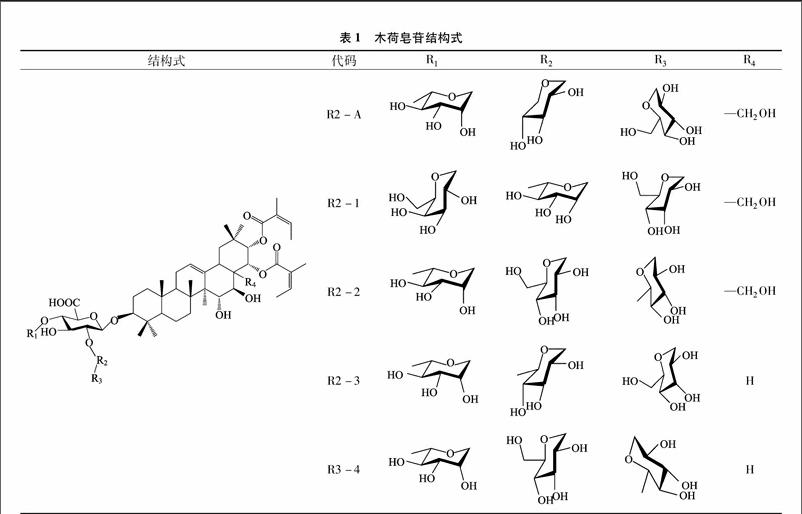
在記載的木荷屬(Schima Reinw.)植物(世界上約有30余種,我國分布21種)中,木荷和紅木荷(Schima wallichii)被認為是有毒植物。已報道的木荷屬植物皂苷有5種,它們均屬齊墩果烷型三萜皂苷,其苷元有玉蕊醇(barrigenol)、高根二醇(erythrodiol)、山茶皂苷元A(camelliagenin A)。某些皂苷在3位連接1個由鼠李糖、半乳糖、葡萄糖和葡萄糖醛酸組成的糖基側鏈,在22位連接1個當歸酰酯基[2-5]。某些皂苷顯示出較強的腫瘤細胞生長抑制活性[2,5-6]。
筆者所在課題組以稻瘟病病菌為靶標生物,篩選并分離出具有強殺稻瘟病菌活性的木荷皂苷混合物M2IGR5(EC50=16.41 μg/mL),其有效濃度與生產上使用的化學品富士1號防治稻瘟病中使用的有效濃度相當[7]。用它處理稻瘟病病菌,導致菌絲斷裂、細胞腫脹、去壁的原生質體變形黏連、細胞膜通透性增大、完整性喪失,從而引起內含物大量滲漏,甚至還間接引起細胞壁降解,加速了稻瘟病病菌細胞的凋亡[8]。從其級分M2IGR5R1中分離鑒定報道了2種新的齊墩果烷型三萜皂苷:母核屬于含有5個六元環的玉蕊醇,環上有7個甲基,3個羥基,2個當歸酰酯基取代,12、13位有1個雙鍵,3位通過氧原子連接1個寡糖基(由鼠李糖、半乳糖、葡萄糖和葡萄糖醛酸基組成或由鼠李糖、木糖、葡萄糖和葡萄糖醛酸基組成),但抗稻瘟病病菌活性不及混合物[9]。進一步對其級分M2IGR5R2、M2IGR5R3分離又獲得7種活性皂苷,其中5種活性較強[10]。本研究從稻瘟病原細胞水平上,研究5種木荷皂苷對稻瘟病病菌孢子及附著胞的致毒作用及方式,確定抗稻瘟病病菌活性皂苷先導物,為開發新型木荷皂苷殺稻瘟劑提供了分子設計思路。
1材料與方法
1.1材料
供試菌株及水稻:稻瘟病菌(Magnaporthe oryzae)10Z3-5,由江西省農業科學院植物保護研究所提供;汕B稻(Oryza Sativa L.),由江西農業大學作物生理和栽培江西省重點實驗室提供。木荷皂苷R2-A、R2-1、R2-2、R2-3、R3-4,由江西省南昌市生物資源保護和利用重點實驗室分離制得[10](表1)。
藥劑及培養基:用0.1%二甲基亞砜(DMSO)溶液溶解各個皂苷,配制木荷皂苷母液終濃度為100 μg/mL。以市售30%稻瘟靈(isoprothiolane)乳油為陽性對照(四川省川東農藥化工有限公司)。馬鈴薯葡萄糖瓊脂(potato dextrose agar,簡稱PDA)培養基配方:去皮馬鈴薯200 g,葡萄糖20 g,瓊脂20 g,加蒸餾水至 1 L,121 ℃濕熱滅菌30 min;稻稈汁玉米粉(straw juice corn flour agar)培養基配方:水稻稈40 g,玉米粉20 g,瓊脂20 g,加稻稈浸泡液至1 L,121 ℃濕熱滅菌30 min。
1.2方法
1.2.1稻瘟病病菌培養及孢子制備
將稻瘟病病菌菌種接于PDA培養基中,28 ℃黑暗活化培養7 d,轉接于稻稈汁玉米粉培養基中,28 ℃黑暗培養5 d后,交替光照(光—暗周期12 h—12 h)培養2~3 d促其產孢,每皿加滅菌水 10 mL,滅菌毛筆刷洗菌落表面孢子,4層紗布過濾除菌絲,將孢子懸浮液用無菌水調整至濃度為1萬個/mL備用[11-12]。
1.2.2孢子萌發率的測定
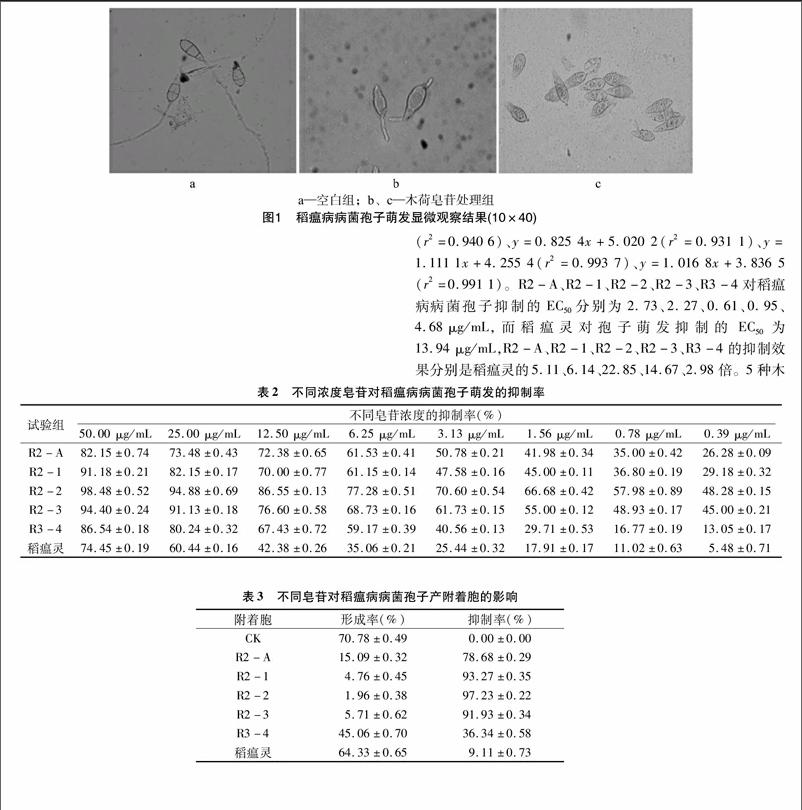
取無菌的96孔板,轉移50 μL上述孢子懸浮液至每個孔,分別吸取50 μL R2-A、R2-1、R2-2、R2-3、R3-4等5種木荷皂苷母液加入第1個孔中,混勻后移50 μL至第2孔,依次進行2倍稀釋,制成50.00、25.00、12.50、6.25、3.13、1.56、0.78、0.39 μg/mL濃度梯度的帶藥孢子懸液,用0.1% DMSO無菌水溶液作空白對照(CK)。28 ℃黑暗保濕培養,血球計數板顯微鏡觀察孢子萌發情況,以芽管長度≥孢子短徑視為萌發,待空白組大部分孢子萌發后,隨機選取視野,觀察約200個分生孢子,統計孢子萌發數量,計算抑制率,相關公式如下:
[JZ]孢子萌發率=萌發的孢子數/鏡檢的孢子總數×100%;
抑制率=(對照孢子萌發率-藥劑處理孢子萌發率)/對照孢子萌發率×100%。
1.2.3孢子產附著胞能力測定
取洋蔥內表皮(從外向內第3、4層),剪為18 mm×18 mm方塊,三氯甲烷中浸泡 20 min,其間輕輕擺動表皮,無菌水沖洗3遍,黏附于無菌載玻片上(2 mol/L 硫酸溶液浸泡30 min,無菌水沖洗至中性),風干。在洋蔥表皮上滴加含10 μg/mL帶藥的孢子懸液 50 μL,用0.1% DMSO無菌水溶液作空白對照(CK)。28 ℃保濕培養,顯微鏡觀察孢子產生附著胞情況,待空白組大部分孢子產生附著胞時,隨機選取視野,觀察約200個分生孢子,統計孢子產生附著胞數量,計算抑制率[13],相關公式如下:
附著胞形成率=產生的附著胞數/鏡檢的孢子總數×100%;
附著胞抑制率=(對照附著胞形成率-藥劑處理附著胞形成率)/對照附著胞形成率×100%。
1.2.4附著胞產黑色素的測定
用乙醇擦拭賽璐玢薄膜,無菌水漂洗晾干,在膜上滴加20 μL濃度為0.5、1.0、1.5、2.0、2.5、3.0、3.5、4.0、4.5、5.0、5.5、6.0 μg/mL的帶藥孢子懸液,28 ℃保濕培養2 d,在顯微鏡下觀察附著胞色素產生情況,不產黑色素或呈現淺黃色的最低濃度即為抑制附著胞黑色素化的MIC。在上述梯度濃度范圍內變色不明顯的,用同樣方法放大濃度進一步測定[14-15]。
1.2.5孢子侵染活體水稻能力測定
將水稻種子用75%乙醇消毒,無菌水沖洗后,平鋪于墊有濾紙片的培養皿(內徑為11 cm)中,加無菌水保濕,28 ℃恒溫培養至發芽,待芽長5 cm左右,移栽至盛有泥土的瓷盆(內徑20 cm)中,自然條件培養,待水稻長至2~3葉期時施少量尿素保綠、鉀肥增寬葉面。用含1%吐溫-40的無菌水配制含各皂苷終濃度為 10 μg/mL 的稻瘟病病菌孢子懸浮液(5×103個/mL),選取水稻葉片長勢基本一致的葉片,在葉片上滴加20 μL上述帶藥孢子懸浮液和對照孢子懸浮液。保濕培養6 d后,觀察發病病情,并拍照。
1.3數據分析
數據處理采用DPS 7.5軟件進行統計分析[16]。
2結果與分析
[HTK]2.1對稻瘟病病菌孢子萌發的致毒作用[HT]
健康的稻瘟病病菌孢子飽滿完整,雙層隔膜明顯,大部分孢子從頂端萌發出芽管(圖1-a)。受木荷皂苷作用后,孢子受到一定程度的損傷,隔膜不明顯,甚至有從隔膜處斷裂成為單隔膜孢子現象,極少數會從2端或側面萌發出芽管(圖1-b),甚至芽管不萌發(圖1-c)。[FL)]
空白組孢子產生的附著胞圓潤飽滿,黑色素積累量多,成熟的附著胞會從附著胞小孔產生粗壯的侵染栓(圖2-a);皂苷作用后的附著胞形態有所不同,孢子尖端只產生芽管或略膨大,但不能正常形成附著胞(圖2-b);圖2-c所示為受皂苷作用后稻瘟病病菌孢子產生中空附著胞,且附著胞內沒有黑色素;受藥后孢子產生的附著胞不能積聚黑色素,且相比正
2.4對孢子侵染活體水稻葉片的致病力影響
各處理組濃度為10 μg/mL,經過6 d的發病培養后,對孢子侵染水稻葉片組織的抑制效果不同。CK組孢子侵染力強,發病嚴重,單位面積上菌斑數量多,病斑連接成片;稻瘟靈處理組出現6個明顯的病斑,病斑擴大趨勢明顯;R2-A組見4個小的侵染斑點,R2-1組僅有2個明顯病斑出現,R2-2 對稻瘟病病菌的孢子有強烈的抑制作用,葉面上完全沒有病斑出現,R2-3對稻瘟病病菌孢子也具有很強的抑制作用,只出現1個病斑,R3-4組發病嚴重,抗病效果較弱,對稻瘟病病菌孢子在活體水稻葉片上的侵染能力影響較小。5種木荷皂苷抗稻瘟病病菌侵染水稻能力大小排序為R2-2>R2-3>R2-1>R2-A>R3-4>稻瘟靈。
3結論與討論
水稻是我國乃至世界的主要糧食作物之一,稻瘟病是引起水稻減產的主要病害。稻瘟病是由稻瘟病原(Magnaporthe oryzae)引起的分布廣、發病部位多、潛伏期長、危害嚴重的全球性病害,一般可導致水稻減產11%~30%[17]。稻瘟病病菌主要以分生孢子在適宜條件下對水稻進行侵染和病害傳播,其侵染過程有以下幾步[18]:(1)稻瘟病病菌孢子接觸寄主表皮后,孢子尖端釋放黏膠使孢子緊密黏附于細胞表面;(2)孢子萌發出芽管后繼而分化為附著胞;(3)附著胞產生侵染釘穿透角質層及表皮細胞壁;(4)病菌分化為次生菌絲在寄主細胞中生長并繼續侵染鄰近細胞,且能深入葉肉細胞;(5)5~7 d開始出現發病癥狀,分生孢子梗上分化出大量的新孢子并脫落釋放出來;(6)新形成的分生孢子可重新侵染寄主細胞,后期以菌絲體和分生孢子在病稻草、病稻谷等上越冬,完成侵染循環。由此可見,孢子及附著胞在整個侵染過程中起著關鍵作用。稻瘟病病菌附著胞的形成不僅受遺傳因子調控,還受外界因素的誘導[19-20]。
R2-A、R2-1、R2-2、R2-3、R3-4的EC50分別為 2.73、2.27、0.61、0.95、4.68 μg/mL,而陽性對照藥物稻瘟靈對抑制稻瘟病病菌孢子萌發的EC50為13.94 μg/mL。5種皂苷的抑菌活性分別是稻瘟靈的5.11、6.14、22.85、14.67、298倍;各皂苷濃度為10 μg/mL時,抑制稻瘟病病菌孢子產附著胞率分別為 78.68%、93.27%、97.23%、91.93%、36.34%,而稻瘟靈的僅為9.11%,各皂苷對抑制稻瘟病病菌孢子產附著胞的能力分別是稻瘟靈的8.64、10.24、10.67、10.09、3.99倍;各皂苷對附著胞黑色素化的MIC分別是7.0、6.0、4.0、65、17.0 μg/mL,稻瘟靈的MIC為18.0 μg/mL,其抑制附著胞黑色素化能力分別是稻瘟靈的2.57、3.00、4.50、2.77、106倍;5種木荷皂苷在稻瘟病病菌孢子侵染活體水稻葉片試驗中表現不同,其抗稻瘟病病菌侵染能力大小排序為 R2-2>R2-3>R2-1>R2-A>R3-4>稻瘟靈。其中 R2-1、R2-2、R2-3等3種皂苷對稻瘟病病菌細胞的致毒作用強烈,受藥后大部分孢子萌發活力下降;少數孢子尖端只產生芽管而不能進一步萌發為附著胞,且附著胞數量明顯減少、直徑縮小,附著胞內的色素積累減少,有的甚至萎縮、中空、透明,從而使附著胞難以形成高壓滲透釘,不能產生足夠的膨壓侵染穿透組織。由此可見,R2-1、R2-2、R2-3是木荷抗稻瘟病病菌的主要活性先導物。
目前,對稻瘟病病菌的防治藥物多種多樣,不同藥物對稻瘟病病菌的抑菌機制各不相同。茶多酚對稻瘟病菌作用機制主要是破壞菌體的細胞膜結構,抑制過氧化氫酶(CAT)、過氧化物酶(POD)活性,使其喪失細胞膜的屏障和酶系的保護功能,最終導致菌體生長受到抑制或死亡[21]。皂苷抗真菌的主要活性機制是與病原菌生物膜上的甾醇形成復合物,導致微孔形成,使生物膜失去完整性[22]。茶皂素屬三萜類皂苷,對稻瘟菌的菌絲生長和孢子萌發有明顯的抑制作用,對三環唑防治稻瘟病具有明顯的增效作用[23]。細胞自噬在稻瘟病病菌的孢子萌發和附著胞形成過程中起著很重要的作用,自噬缺陷的稻瘟病病菌菌株缺乏足夠的膨壓而不能侵入水稻表皮,而自噬缺陷菌株的單個孢子中萌發的細胞數和形成的附著胞數與野生型沒有差異[24-26]。稻瘟病病菌侵入主要是靠機械作用,但酶或其他作用也可能輔助或加速這一過程的進行。三環唑等農藥可以抑制[WTBX][STBX]BUF1[WTBZ][STBZ]基因所編碼的酶活性,從而能夠有效阻斷黑色素的合成,令稻瘟病病菌附著胞難以形成高壓滲透釘而失去侵染性[18],但在受傷的水稻表面,稻瘟病病菌菌絲可直接侵染而失去防治效果。木荷皂苷對稻瘟病病菌的致毒作用,主要作用于病原細胞膜上,不僅導致菌絲細胞完整性和通透性損傷、內含物外泄[8],而且還可抑制孢子萌發、孢子產附著胞及附著胞積聚黑色素等多個環節,阻斷稻瘟病病菌侵染過程,有效控制稻瘟病病菌的發生和傳播,不像三環唑、木荷皂苷對不同類型稻瘟病病原細胞不具備選擇性。
參考文獻:
[1]鄧志勇,鄧業成,劉艷華. 木荷提取物對小菜蛾和菜青蟲的拒食活性[J]. 農藥,2007,46(12):854-856.
[2]Ohtsuki T,Miyagawa T,Koyano T,et al. Acylated triterpenoid saponins from Schima noronhae and their cell growth inhibitory activity[J]. Journal of Natural Products,2008,71(5):918-921.
[3]陳昌祥,葉海亞,申永樂,等. 紅木荷樹皮的化學成分[J]. 云南植物研究,1997,19(2):201-206.
[4]陳維新,吳大剛. 銀木荷苷元研究[J]. 化學學報,1978,36(3):229-232.
[5]徐文,周光雄,戴毅,等. 木荷莖的化學成分研究[J]. 中草藥,2010,41(6):863-866.
[6]Takashi O,Masaaki S,Takashi K,et al. Steroidal saponins from Calamus insignis and their cell growth and cell cycle inhibitory activities[J]. Bioorganic & Medicinal Chemistry,2006,14(3):659-665.
[7]馮華根,張長伶,霍光華. 木荷活性成分的分離及其對稻瘟病菌的室內毒力[J]. 江蘇農業科學,2012,40(4):114-117.
[8]易磊,霍光華,韓啟燦,等. 木荷皂甙對稻瘟病菌細胞形態及生理生化指標的影響[J]. 植物保護學報,2013,40(5):450-456.
[9]Huo G H,Zhang C L,Zhang Y J. Structure elucidation of two Triterpenoid saponins from leaves of Schima superba Gardn. et Champ[J]. Lecture Notes in Electrical Engineering,2014,250(2):915-922.[ZK)]
[10]彭玉萌,霍光華,韓啟燦,等. 抗稻瘟病菌活性木荷皂甙類似物的分離條件及其分離[J]. 分析化學,2014,42(1):59-64.
[11]殷麗青,陳全慶. 培養基和光照對稻瘟病菌產孢量的影響[J]. 上海農業科技,1991(5):43-48.
[12]王國平,羅寬. 稻瘟病菌產孢培養基的篩選[J]. 湖南農學院學報,1989(2):58-62.
[13]廖乾生. 抗稻瘟病菌附著胞形成研究[D]. 杭州:浙江大學,2002.
[14]張欣,靳學慧,張亞玲. 三環唑抑制稻瘟病菌MIC的測定及菌株產黑色素能力比較[J]. 黑龍江八一農墾大學學報,2010,22(2):20-24.[HJ1.73mm]
[15]張傳清,周明國,薛娜. 稻瘟病菌對三環唑的敏感性檢測枝術與抗藥性風險評估[J]. 中國水稻科學,2005,19(1):79-84.
[16]唐啟義. DPS數據處理系統[M]. 北京:科學出版社,2007.
[17]Daw B D,Zhang L H,Wang Z Z. Salicylic acid enhances antifungal resistance to Magnaporthe grisea in rice plants[J]. Australasian Plant Pathology,2008,37(6):637-644.
[18]林福呈. 稻瘟病菌附著胞形成的細胞生物學[J]. 植物病理學報,2001,31(2):97-101.
[19]Jelitto T C,Page H A,Read N D. Role of external signals in regulating the prepenetration phase of infection by the rice blast fungus,Magnaporthe grisea[J]. Planta,1994,194(4):471-477.
[20]Lee Y H,Dean R A. Hydrophobicity of contact surface induces appressorium formation in Magnaporthe grisea[J]. FEMS Microbiology Letters,1994,115(1):71-75.
[21]汪金蓮,邱業先,扶教龍,等. 茶多酚對稻瘟病菌的抑制作用及抑菌機理[J]. 天然產物研究與開發,2011,23(5):918-922.
[22]Charles G,Jean L,Karl G L,et al. Haemolytic activity,cytotoxicity and membrane cell permeabilization of semi-synthetic and natural lupane- and oleanane-type saponins[J]. Bioorganic & Medicinal Chemistry,2009,17(5):2002-2008.
[23]盛書強. 茶皂素對殺菌劑的增效作用及機理[D]. 長沙:湖南農業大學,2003.
[24]Claire V F,Madhumita B,Martin E,et al. Autophagic fungal cell death is necessary for infection by the rice blast fungus[J]. Science,2006,312(5773):580-583.
[25]Liu X H,Lu J P,Zhang L,et al. Involvement of a Magnaporthe grisea serine/threonine kinase gene,[WTBX][STBX]MgATG1[WTBZ][STBZ],in appressorium turgor and pathogenesis[J]. Eukaryot Cell,2007,6(6):997-1005.
[26]Lu J P,Liu X H,Feng X X,et al. An autophagy gene,[WTBX][STBX]MgATG5[WTBZ][STBZ],is required for cell differentiation and pathogenesis in Magnaporthe oryzae[J].
