乙型肝炎病毒剪接特異性蛋白HBSP與TGFβ1誘導(dǎo)蛋白1相互作用促進(jìn)TGFβ1誘導(dǎo)的肝癌細(xì)胞上皮間質(zhì)轉(zhuǎn)化
2017-04-26 08:54:06陳婉南黃俊高梁菲菲閆小利軒丹丹
中國人獸共患病學(xué)報(bào) 2017年4期
關(guān)鍵詞:肝癌
陳婉南,黃俊高,梁菲菲,閆小利,軒丹丹,林 旭
?
乙型肝炎病毒剪接特異性蛋白HBSP與TGFβ1誘導(dǎo)蛋白1相互作用促進(jìn)TGFβ1誘導(dǎo)的肝癌細(xì)胞上皮間質(zhì)轉(zhuǎn)化
陳婉南,黃俊高,梁菲菲,閆小利,軒丹丹,林 旭
目的 探討乙型肝炎病毒剪接特異性蛋白(Hepatitis B spliced protein, HBSP)與轉(zhuǎn)化生長因子1誘導(dǎo)蛋白1(transforming growth factor-β1-induced transcript 1,TGFβ1I1)相互作用對TGFβ1誘導(dǎo)的肝癌細(xì)胞上皮間質(zhì)轉(zhuǎn)化(epithelial-mesenchymal transition,EMT)的影響。方法 構(gòu)建HBSP慢病毒表達(dá)載體,利用293T細(xì)胞包裝慢病毒顆粒,感染Huh7肝癌細(xì)胞株。以5 ng/mLTGFβ1分別誘導(dǎo)穩(wěn)定表達(dá)HBSP的慢病毒細(xì)胞株及其對照細(xì)胞株,觀察細(xì)胞形態(tài)的變化,并提取細(xì)胞蛋白,Westernblot檢測上皮間質(zhì)轉(zhuǎn)化標(biāo)志物E-鈣黏素(E-cadherin,E-cad)、緊密連接蛋白(Claudin-1)、β-鏈蛋白(β-catenin)、N-鈣黏素(N-cadherin,N-cad)的變化。進(jìn)而以TGFβ1I1特異性siRNA轉(zhuǎn)染上述細(xì)胞,Westernblot觀察以上指標(biāo)變化情況。最后以侵襲小室實(shí)驗(yàn)和劃痕實(shí)驗(yàn)分別檢測TGFβ1誘導(dǎo)的細(xì)胞侵襲與遷移能力的變化。結(jié)果 篩選獲得穩(wěn)定表達(dá)HBSP的慢病毒細(xì)胞株Huh7-HBSP-flag-HIV及其對照細(xì)胞株Huh7-flag-HIV。以TGFβ1誘導(dǎo)后在顯微鏡下觀察到細(xì)胞形態(tài)由緊密的上皮形態(tài)變?yōu)樗缮⒌拈g質(zhì)形態(tài);特異性抗體檢測表明上皮標(biāo)志物E-cad、Claudin-1、β-catenin表達(dá)量下降,而間質(zhì)標(biāo)志物N-cad表達(dá)上升。侵襲小室實(shí)驗(yàn)和劃痕實(shí)驗(yàn)表明TGFβ1誘導(dǎo)的HBSP表達(dá)株侵襲及遷移能力增強(qiáng)。而轉(zhuǎn)染TGFβ1I1特異性siRNA可逆轉(zhuǎn)以上現(xiàn)象。結(jié)論 HBSP與TGFβ1I1相互作用可促進(jìn)TGFβ1誘導(dǎo)的肝癌細(xì)胞上皮間質(zhì)轉(zhuǎn)化并增強(qiáng)其侵襲遷移能力,提示HBSP在HBV相關(guān)性肝細(xì)胞肝癌發(fā)生發(fā)展中具有重要的致病意義。
乙型肝炎病毒;RNA剪接;轉(zhuǎn)化生長因子β1誘導(dǎo)蛋白1;上皮間質(zhì)轉(zhuǎn)化;侵襲
乙型肝炎病毒剪接特異蛋白HBSP由HBV前基因組RNA (pregenomic RNA, pgRNA)在2 447 nt-489 nt發(fā)生RNA剪接產(chǎn)生的長度為2.2 kb的剪接變異體編碼產(chǎn)生,已有研究表明該蛋白在HBV感染不同病程均可檢出,與病毒的持續(xù)性感染及致病性相關(guān)[1-4]。HBSP定位于細(xì)胞漿[5],為了深入闡明HBSP在HBV導(dǎo)致肝癌發(fā)生發(fā)展的過程中具體的致病機(jī)制,本實(shí)驗(yàn)室在前期工作中采用細(xì)胞質(zhì)酵母雙雜交(CytoTrapTMtwo-hybrid system,Stratagene)篩選的方法,發(fā)現(xiàn)HBSP可以和轉(zhuǎn)化生長因子1誘導(dǎo)蛋白1相互作用[6]。已有研究表明,TGFβ1I1可以促進(jìn)鼠和人的上皮細(xì)胞發(fā)生上皮間質(zhì)轉(zhuǎn)化,對腫瘤的發(fā)生發(fā)展有著重要意義[7]。本文旨在通過研究HBSP與TGFβ1I1相互作用對肝癌細(xì)胞株上皮間質(zhì)轉(zhuǎn)化以及侵襲性的影響,更好地闡明HBV導(dǎo)致肝癌發(fā)生發(fā)展的機(jī)制。
1 材料和方法
1.1 材料
1.1.1 質(zhì)粒與細(xì)胞 Huh7肝癌細(xì)胞株由本實(shí)驗(yàn)室保存。人胚腎293T細(xì)胞株、慢病毒表達(dá)載體pCDH-EF1-MCS-T2A-Puro與包裝質(zhì)粒pMDL、pVSVG、pREV由廈門大學(xué)吳喬教授惠贈。
1.1.2 主要試劑 高保真Taq DNA、轉(zhuǎn)染試劑Lipofectamine 3000聚合酶購自Invitrogen公司,限制性內(nèi)切酶和T4 DNA連接酶購自NEB公司,質(zhì)粒提取試劑盒購自Qiagen公司。DMEM高糖培養(yǎng)基、胰酶、Puromycin抗生素購自Gibco公司,胎牛血清購自PAN公司,磷酸鈣轉(zhuǎn)染試劑盒、anti-TGFβ1I1抗體和Boyden Transwell侵襲小室購自BD公司。TGFβ1購自Sigma-Aldrich公司,TGFβ1I1特異性siRNA混合物及無關(guān)序列對照NC-siRNA、兔二抗購自Santa Cruz公司。RIPA裂解液、BCA蛋白定量試劑盒購自碧云天生物技術(shù)公司,PVDF膜購自Millipore公司,CDP-STAR化學(xué)發(fā)光檢測底物購自Roche公司。anti-E-cadherin,anti-N-cadherin,anti-β-catenin,anti-Claudin-1抗體、anti-Flag抗體購自Cell Signaling Technology公司,anti-β-actin抗體購自康為世紀(jì),anti-alpha Smooth Actin抗體、anti-Fibronectin抗體購自Abcam公司,鼠二抗購自Calbiochem公司。
1.2 方法
1.2.1 慢病毒載體pCDH-HBSP的構(gòu)建 以pSos-HBSP為模板[6],PCR擴(kuò)增獲得C末端帶有FLAG標(biāo)簽的HBSP基因,插入慢病毒表達(dá)載體pCDH-EF1-MCS-T2A-Puro,獲得重組載體pCDH-HBSP-FLAG。空載體對照pCDH-FLAG為將8個(gè)氨基酸大小的FLAG編碼基因插入上述慢病毒表達(dá)載體獲得。
1.2.2 慢病毒包裝與感染、篩選穩(wěn)定表達(dá)HBSP的肝癌細(xì)胞株 使用以下質(zhì)粒混合物、按照試劑盒說明書以磷酸鈣轉(zhuǎn)染法轉(zhuǎn)染293T細(xì)胞:pCDH-HBSP-FLAG或pCDH-FLAG 2 μg、pMDL 1 μg、pVSVG 0.6 μg、pREV 0.4 μg, 48 h后收集含有病毒的細(xì)胞培養(yǎng)上清液,離心過濾后均勻滴加至Huh7肝癌細(xì)胞中,以1 μg/mL Puromycin抗生素篩選培養(yǎng)2周,取106個(gè)細(xì)胞接種于60 mm培養(yǎng)皿。次日以RIPA裂解液提取細(xì)胞總蛋白,以anti-FLAG抗體(1∶2 000稀釋)檢測目的蛋白HBSP的表達(dá)。其余細(xì)胞凍存至-80 ℃?zhèn)溆谩?/p>
1.2.3 TGFβ1處理穩(wěn)定表達(dá)HBSP的肝癌細(xì)胞株
將經(jīng)過鑒定穩(wěn)定表達(dá)HBSP的Huh7細(xì)胞株于加藥處理前一天接種5×105個(gè)細(xì)胞于35 mm板。次日用PBS清洗后換成不含血清的DMEM培養(yǎng)液,加入TGFβ1(終濃度為5 ng/mL)作用24 h后觀察細(xì)胞形態(tài)的變化。
1.2.4 Westernblot檢測TGFβ1處理后EMT相關(guān)標(biāo)志物 同上以5 ng/mL的TGFβ1處理HBSP表達(dá)細(xì)胞株,24 h后提取細(xì)胞總蛋白,以EMT標(biāo)志物E-cad,N-cad,β-catenin,Claudin-1抗體(均為1∶1 000稀釋)檢測、Quantity One軟件(Biorad)分析計(jì)算蛋白條帶的灰度值。以各組蛋白條帶的灰度值相對于內(nèi)參β-actin條帶的灰度值進(jìn)行標(biāo)準(zhǔn)化。實(shí)驗(yàn)重復(fù)5次取平均值作圖分析。
1.2.5 TGFβ1I1特異性siRNA轉(zhuǎn)染,Westernblot檢測EMT指標(biāo) 同上以5 ng/mL的TGFβ1處理HBSP表達(dá)細(xì)胞株,24 h后采用Lipofectamine 3000轉(zhuǎn)染TGFβ1I1特異性siRNA混合物,使用無關(guān)序列NC-siRNA作為對照。轉(zhuǎn)染方法參照試劑說明書,轉(zhuǎn)染后24 h提取細(xì)胞總蛋白,同上檢測EMT相關(guān)標(biāo)志物。實(shí)驗(yàn)重復(fù)5次取平均值作圖分析。
1.2.6 侵襲小室實(shí)驗(yàn) HBSP表達(dá)細(xì)胞株以5 ng/mL的TGFβ1處理24 h后,用TGFβ1I1特異性siRNA混合物以及對照NC-siRNA轉(zhuǎn)染細(xì)胞24 h,以無血清培養(yǎng)液吹打混勻消化后的細(xì)胞,分別計(jì)數(shù)5 ×104個(gè)細(xì)胞接種到Boyden Transwell小室內(nèi),36 h,后用 0.1 %結(jié)晶紫染液染色,在倒置顯微鏡下隨機(jī)取5個(gè)視野計(jì)數(shù)細(xì)胞取平均值并分別拍照,實(shí)驗(yàn)重復(fù)5次。
1.2.7 劃痕實(shí)驗(yàn) 同上獲得TGFβ1處理并使用TGFβ1I1特異性siRNA混合物轉(zhuǎn)染的HBSP表達(dá)細(xì)胞株,接種于6孔板次日用滅菌槍頭在培養(yǎng)皿底部垂直劃出一道直線,在倒置顯微鏡下隨機(jī)取5個(gè)視野拍照并測量劃痕寬度并取平均值,繼續(xù)無血清培養(yǎng)48 h后,拍照測量劃痕寬度取平均值,實(shí)驗(yàn)重復(fù)5次。
2 結(jié) 果
2.1 慢病毒感染Huh7細(xì)胞,篩選獲得穩(wěn)定表達(dá)HBSP的肝癌細(xì)胞株 PCR擴(kuò)增HBSP基因,經(jīng)XbaI和BamH I雙酶切位點(diǎn)克隆入慢病毒表達(dá)載體pCDH-EF1-MCS-T2A-Puro,重組載體經(jīng)酶切(圖1A)及測序證實(shí)插入的HBSP DNA片段正確無誤,命名為pCDH-HBSP-FLAG。包裝載體pMDL、pVSVG、pREV,以及pCDH-HBSP-FLAG或空載體對照pCDH-FLAG共轉(zhuǎn)染293T細(xì)胞;獲得慢病毒液,感染并以Puromycin篩選Huh7細(xì)胞,以anti-FLAG抗體檢驗(yàn)?zāi)康牡鞍椎谋磉_(dá)(圖1B),獲得的穩(wěn)定表達(dá)HBSP的細(xì)胞株命名為Huh7-HBSP-flag-HIV,對照細(xì)胞株命名為Huh7-flag-HIV。
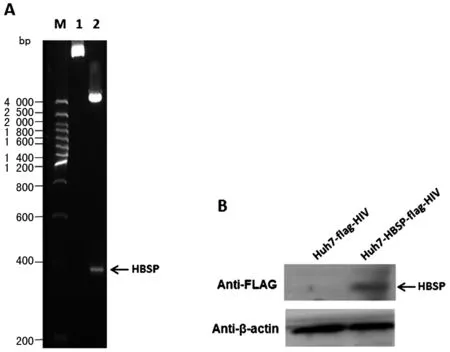
A.慢病毒重組載體雙酶切鑒定 B.1. 未經(jīng)酶切的重組載體;2. XbaI和BamH I雙酶切重組載體 B.Westernblot檢測Huh7細(xì)胞株HBSP蛋白的表達(dá)A.Restriction enzyme analysis of recombinant lentiviral expression vector 1.uncut recombinant plasmids; 2. recombinant plasmids digested with XbaI and BamH I B.Westernblot detection of HBSP expression in the established Huh7 cell lines圖1 慢病毒表達(dá)載體構(gòu)建及Westernblot檢測HBSP蛋白表達(dá)的Huh7慢病毒細(xì)胞株Fig.1 Lentiviral vector construction and HBSP protein detection in the established Huh7 cell lines
2.2 HBSP 促進(jìn)TGFβ1誘導(dǎo)的肝癌細(xì)胞上皮間質(zhì)轉(zhuǎn)化 以5 ng/mL的TGFβ1誘導(dǎo)Huh7-HBSP-flag-HIV與對照細(xì)胞株Huh7-flag-HIV24 h,鏡下可見細(xì)胞形態(tài)發(fā)生變化:處理前,對照組Huh7-flag-HIV細(xì)胞呈鋪路石樣,細(xì)胞之間連接緊密,呈現(xiàn)典型的上皮細(xì)胞形態(tài)(圖2A),而Huh7-HBSP-flag-HIV 細(xì)胞(圖2B)為多邊形,部分為梭形;經(jīng)過5 ng/mL TGFβ1處理24 h,Huh7-flag-HIV 細(xì)胞間隙變寬,細(xì)胞間連接變得松散,多呈間質(zhì)細(xì)胞形態(tài)(圖2C),而Huh7-HBSP-flag-HIV細(xì)胞比對照細(xì)胞連接更松散,細(xì)胞變長呈梭形,幾乎全為間質(zhì)細(xì)胞形態(tài)(圖2D)。提示HBSP促進(jìn)TGFβ1誘導(dǎo)的肝癌細(xì)胞發(fā)生上皮間質(zhì)轉(zhuǎn)化。
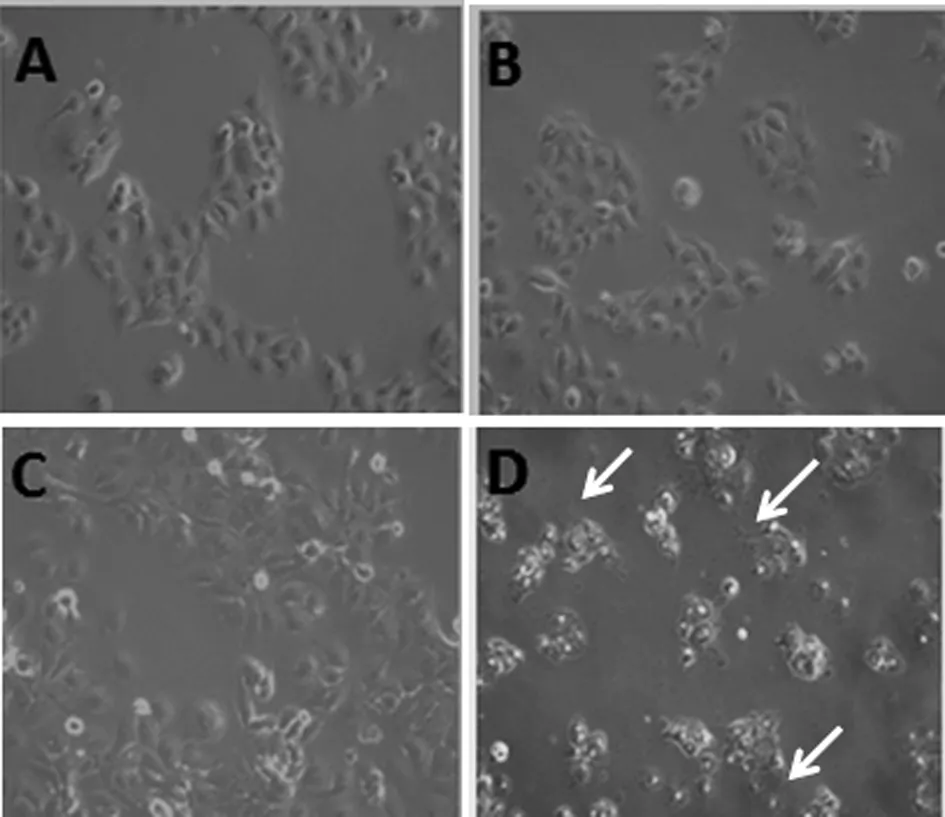
A:Huh7-flag-HIV細(xì)胞; B:Huh7-HBSP-flag-HIV細(xì)胞; C:5 ng/mL TGFβ1處理Huh7-flag-HIV細(xì)胞24 h后;D:5 ng/mL TGFβ1處理Huh7-HBSP-flag-HIV細(xì)胞24 h后A:Huh7-flag-HIVcells; B:Huh7-HBSP-flag-HIV cells; C:Huh7-flag-HIV cells treated with 5 ng/mL TGFβ1 for 24 hours; D:Huh7-HBSP-flag-HIV cells treated with 5 ng/mL TGFβ1 for 24 hours.圖2 TGFβ1處理前后 Huh7-HBSP-flag-HIV 及對照組 Huh7-flag-HIV 細(xì)胞株形態(tài)學(xué)變化(10×)Fig.2 Huh7-HBSP-flag-HIV and Huh7-flag-HIV cells images before and after TGFβ1 treatment (10×)
2.3 HBSP促進(jìn)TGFβ1誘導(dǎo)的肝癌細(xì)胞上皮間
質(zhì)轉(zhuǎn)化相關(guān)指標(biāo)變化 以5 ng/mL TGFβ1誘導(dǎo)細(xì)胞24 h后,提取細(xì)胞總蛋白,用特異性抗體檢測EMT相關(guān)指標(biāo)表達(dá)水平,結(jié)果顯示,Huh7-HBSP-flag-HIV與Huh7-flag-HIV對照細(xì)胞株相比,上皮樣標(biāo)志物E-cadherin、Claudin-1、β-catenin表達(dá)量都出現(xiàn)明顯下降(n=5,P<0.01),而間質(zhì)樣標(biāo)志物N-cadherin表達(dá)量出現(xiàn)明顯升高(n=5,P<0.01)(圖3)。HBSP在肝癌細(xì)胞株內(nèi)過表達(dá)可下調(diào)TGFβ1誘導(dǎo)的肝癌細(xì)胞上皮樣標(biāo)志物表達(dá)水平,并上調(diào)間質(zhì)樣標(biāo)志物表達(dá)水平,進(jìn)一步證明HBSP可促進(jìn)TGFβ1誘導(dǎo)的肝癌細(xì)胞發(fā)生上皮間質(zhì)轉(zhuǎn)化。
2.4 下調(diào)TGFβ1I1可逆轉(zhuǎn)TGFβ1誘導(dǎo)的HBSP表達(dá)細(xì)胞株上皮間質(zhì)轉(zhuǎn)化 以5 ng/mL TGFβ1誘導(dǎo)Huh7-HBSP-flag-HIV以及Huh7-flag-HIV對照株,并分別轉(zhuǎn)染TGFβ1I1 siRNA混合物與無關(guān)對照NC-siRNA,觀察TGFβ1I1表達(dá)水平被特異性siRNA下調(diào)后(圖4A)EMT相關(guān)指標(biāo)的變化情況。結(jié)果顯示:轉(zhuǎn)染無關(guān)對照NC-siRNA組,Huh7-HBSP-flag-HIV細(xì)胞與Huh7-flag-HIV對照細(xì)胞株相比,上皮細(xì)胞相關(guān)指標(biāo)E-cadherin、Claudin-1、β-catenin表達(dá)量都下降,而間質(zhì)細(xì)胞標(biāo)志物N-cadherin表達(dá)量增高(圖4A,第三泳道與第一泳道對比,n=5,P<0.01);而TGFβ1I1 siRNA 混合物轉(zhuǎn)染Huh7-HBSP-flag-HIV細(xì)胞,其上皮樣標(biāo)志物E-cadherin、Claudin-1、β-catenin表達(dá)量逆轉(zhuǎn)上調(diào),間質(zhì)樣標(biāo)志物N-cadherin表達(dá)下降(圖4A,第四泳道與第三泳道對比,n=5,P<0.01),說明下調(diào)TGFβ1I1可逆轉(zhuǎn)TGFβ1誘導(dǎo)的HBSP表達(dá)細(xì)胞株上皮間質(zhì)轉(zhuǎn)化,提示HBSP借由與TGFβ1I1的相互作用來影響TGFβ1誘導(dǎo)的肝癌細(xì)胞上皮間質(zhì)轉(zhuǎn)化。
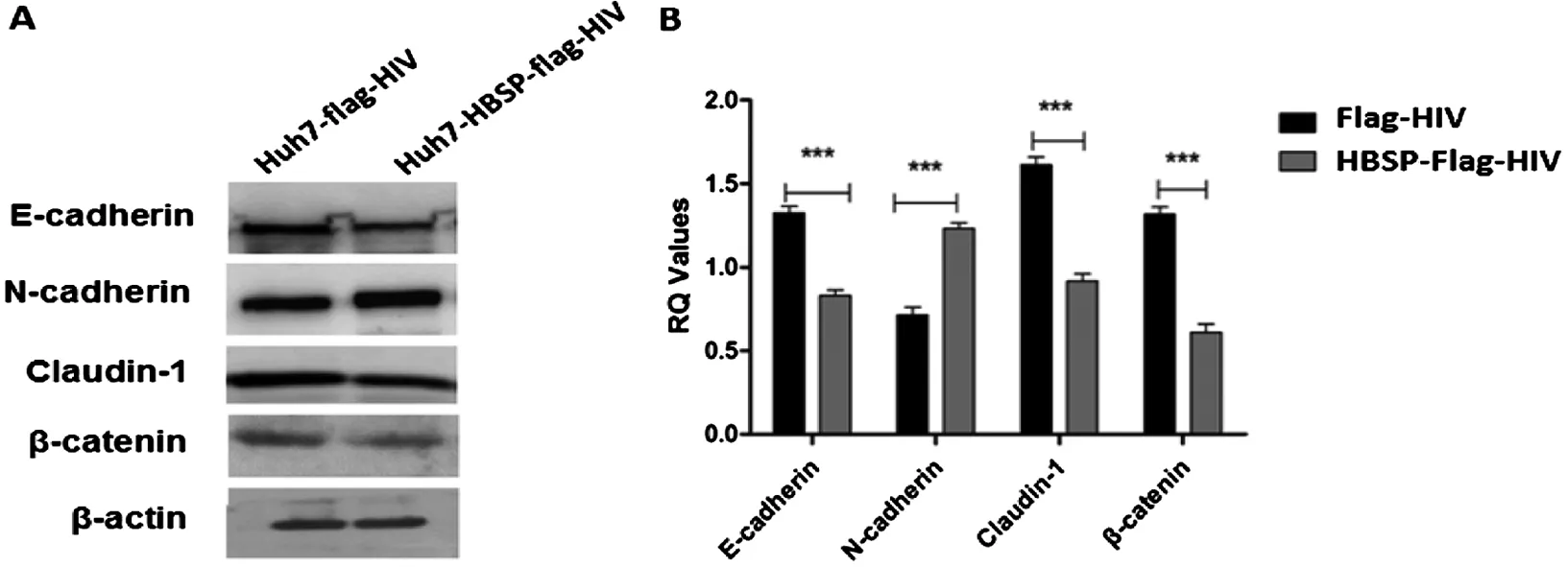
A:Westernblot檢測Huh7 細(xì)胞株EMT標(biāo)志物 B:蛋白條帶灰度值比較A:Westernblot analysis of EMT markers; B:Relative intensities of plotted proteins.*** n=5, P<0.0001圖3 TGFβ1誘導(dǎo)的肝癌細(xì)胞上皮間質(zhì)轉(zhuǎn)化相關(guān)指標(biāo)變化Fig.3 TGFβ1-induced EMT markers detection in Huh7 cell lines
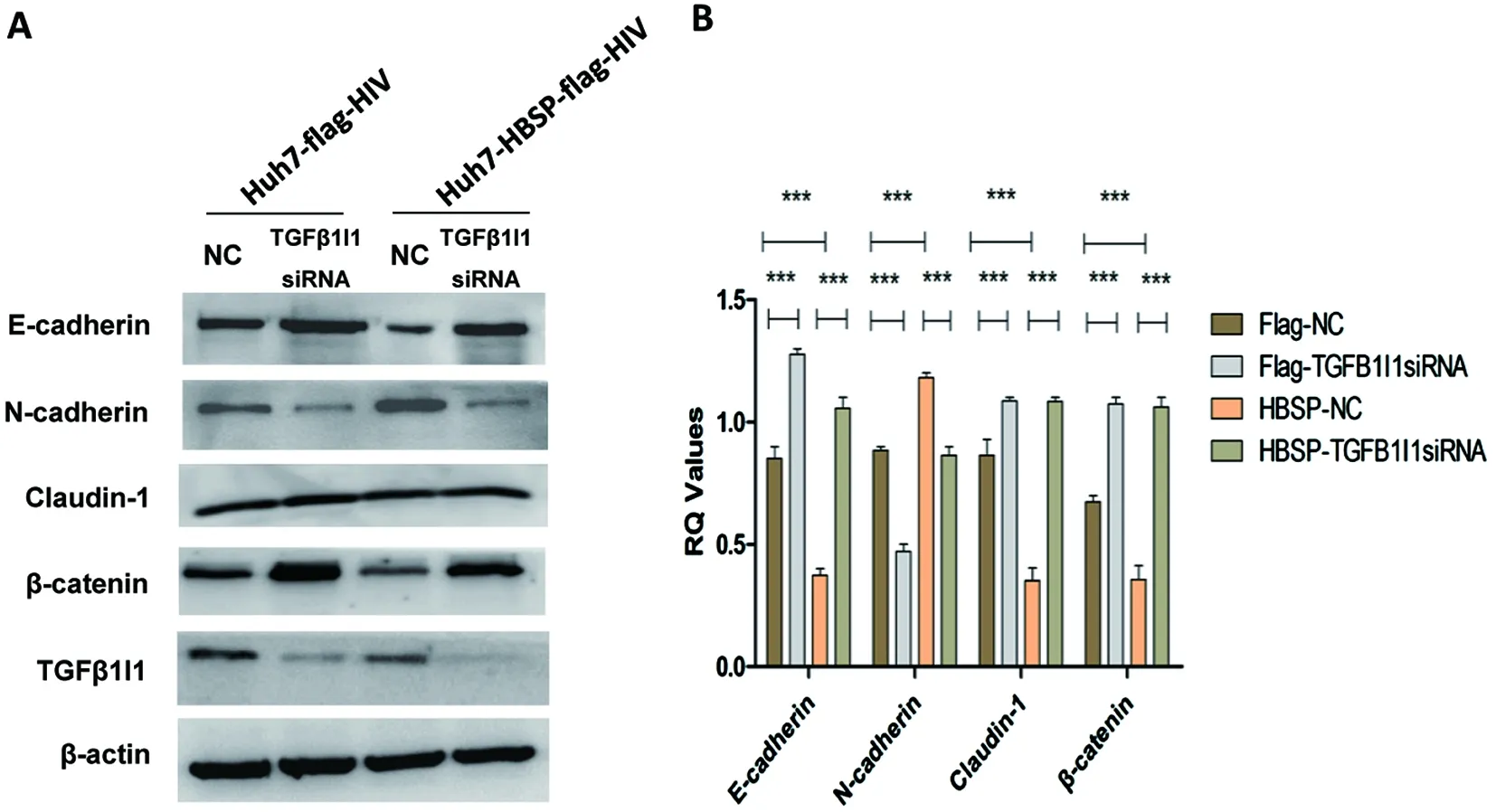
A:Westernblot檢測 EMT標(biāo)志物 B:蛋白條帶灰度值比較A:Western blot detection of EMT markers; B:Relative intensities of plotted proteins.*** n=5, P<0.0001.圖4 siRNA下調(diào)TGFβ1I1可逆轉(zhuǎn)TGFβ1誘導(dǎo)的HBSP表達(dá)細(xì)胞株上皮間質(zhì)轉(zhuǎn)化Fig.4 TGFβ1I1 knockdown by siRNA could reverse epithelial-mesenchymal transition in HBSP expressed Huh7 hepatoma cells
2.5 HBSP與TGFβ1I1的相互作用促進(jìn)TGFβ1誘導(dǎo)的肝癌細(xì)胞侵襲與遷移能力 Huh7-HBSP-flag-HIV以及Huh7-flag-HIV對照株分別以5 ng/mL TGFβ1處理24 h后分別轉(zhuǎn)染TGFβ1I1 siRNA混合物與NC-siRNA,作用24 h后接種于侵襲小室,36 h后觀察到:轉(zhuǎn)染無關(guān)對照NC-siRNA,Huh7-HBSP-flag-HIV細(xì)胞與Huh7-flag-HIV細(xì)胞株相比(見圖5A,第三列與第一列相比),細(xì)胞數(shù)明顯增加(94±3 vs. 29±3,P<0.05),說明HBSP能夠促進(jìn)TGFβ1誘導(dǎo)的肝癌細(xì)胞侵襲能力。Huh7-HBSP-flag-HIV細(xì)胞株轉(zhuǎn)染TGFβ1I1特異性siRNA后(見圖5A,第四列與第三列相比)細(xì)胞數(shù)明顯下降(29±3 vs. 9±2,P<0.05)。
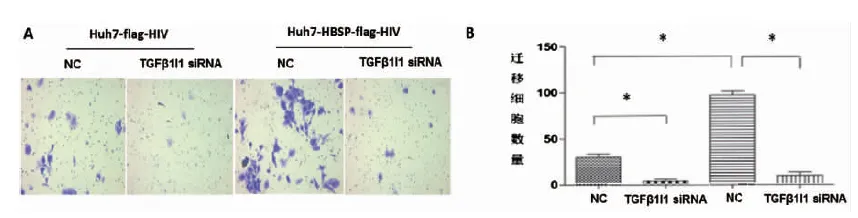
A:鏡下觀察遷移穿過transwell小室的細(xì)胞 B:遷移細(xì)胞數(shù)量比較A:Images of Huh7 invaded through membrane into the bottom of transwell chamber; B:Comparison of invaded cells quantities.* n=5, P<0.05.圖5 HBSP與TGFβ1I1相互作用促進(jìn)TGFβ1誘導(dǎo)的肝癌細(xì)胞侵襲能力Fig.5 HBSP-TGFβ1I1 interaction promotes invasion of TGFβ1-induced Huh7 hepatoma cells
同樣地,對5 ng/mL TGFβ1處理Huh7-HBSP-flag-HIV以及Huh7-flag-HIV對照株轉(zhuǎn)染TGFβ1I1 siRNA混合物與NC-siRNA 24 h后,進(jìn)行劃痕實(shí)驗(yàn),48 h后觀察細(xì)胞修復(fù)情況。結(jié)果與侵襲實(shí)驗(yàn)類似,轉(zhuǎn)染無關(guān)對照NC-siRNA,Huh7-HBSP-flag-HIV細(xì)胞與Huh7-flag-HIV細(xì)胞株相比,細(xì)胞遷移能力明顯加強(qiáng)(見圖6A,第三列與第一列相比),說明HBSP促進(jìn)TGFβ1誘導(dǎo)的肝癌細(xì)胞遷移能力。當(dāng)TGFβ1I1被siRNA下調(diào)后,Huh7-HBSP-flag-HIV細(xì)胞株遷移能力下降(見圖6A,第四列與第三列相比)。上述結(jié)果提示HBSP與TGFβ1I1的相互作用促進(jìn)TGFβ1誘導(dǎo)的肝癌細(xì)胞侵襲與遷移能力。
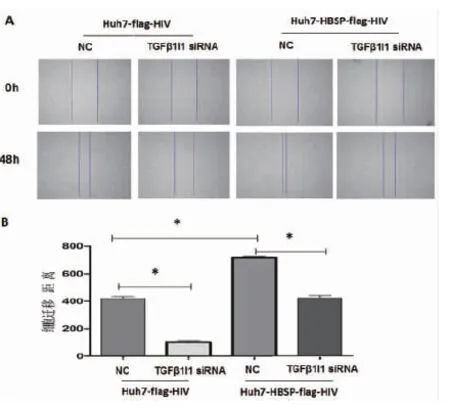
A:鏡下觀察劃痕實(shí)驗(yàn)前后的細(xì)胞 B:劃痕修復(fù)寬度比較A:Images of Huh7 with wound healing assay captured at 0 h and 48 h; B:Quantify the total distance of cells moved from the edge of the scratch toward the center.* n=5,P<0.05.圖6 HBSP與TGFβ1I1相互作用促進(jìn)TGFβ1誘導(dǎo)的肝癌細(xì)胞遷移能力Fig.6 HBSP-TGFβ1I1 interaction promotes migration of TGFβ1-induced Huh7 hepatoma cells
3 討 論
乙型肝炎病毒是一種在其復(fù)制周期中存在逆轉(zhuǎn)錄過程的DNA病毒,由前基因組RNA剪接所產(chǎn)生的剪接變異體可編碼產(chǎn)生一些剪接變異體特異性新蛋白。HBSP是HBV感染者中常見的一種剪接蛋白[8],已有研究認(rèn)為,HBSP可誘導(dǎo)肝細(xì)胞凋亡,加重肝炎病情,還與肝纖維化有關(guān)[2];并通過誘導(dǎo)特異性細(xì)胞毒性T淋巴細(xì)胞(cytotoxic lymphocyte,CTL)活性,導(dǎo)致肝臟損傷[3]。我們先前的研究也顯示HBSP與組織蛋白酶B(cathepsin B, CTSB)相互作用,通過PI3K/Akt通路及MAPK通路增強(qiáng)肝癌細(xì)胞株的侵襲能力[4]; HBSP與纖維蛋白原(fibrinogen gamma chain,F(xiàn)GG)相互作用,抑制其交聯(lián),能抑制血小板凝集、活化,造成凝血障礙[5]; HBSP與微粒體環(huán)氧化物酶(microsomal epoxidehydrolase, mEH)相互作用,通過促進(jìn)后者活性,導(dǎo)致苯丙芘(Benzo(a)pyrene, B[a]P)的致癌產(chǎn)物增加,協(xié)同促進(jìn)肝癌細(xì)胞生長[9]。為了更好地了解定位于細(xì)胞漿內(nèi)的HBSP與肝細(xì)胞蛋白相互作用的情況,本實(shí)驗(yàn)室在前期工作中采用細(xì)胞質(zhì)酵母雙雜交研究與HBSP相互作用的蛋白,篩選出轉(zhuǎn)化生長因子1誘導(dǎo)蛋白1,并利用哺乳動物細(xì)胞雙雜交、免疫共沉淀等方法證實(shí)了HBSP可與其相互作用[6]。
轉(zhuǎn)化生長因子β1誘導(dǎo)蛋白1受TGFβ1誘導(dǎo)[10],因其又受H2O2誘導(dǎo),也稱為Hic-5(hydrogen peroxide-inducible clone-5)[11]。TGFβ1I1對TGFβ1存在正反饋調(diào)節(jié)機(jī)制,即TGFβ1I1具有維持局部組織TGFβ1不斷自分泌的作用[12]。TGFβ1I1可促進(jìn)細(xì)胞發(fā)生上皮間質(zhì)轉(zhuǎn)化[13],且這類由TGFβ1誘導(dǎo)的EMT可促進(jìn)細(xì)胞間基質(zhì)的降解和提高細(xì)胞的侵襲轉(zhuǎn)移能力[14],有研究表明,TGFβ1誘導(dǎo)的EMT與肝細(xì)胞性肝癌發(fā)生發(fā)展密切相關(guān)[15-16]。EMT是腫瘤轉(zhuǎn)移的重要機(jī)制之一,主要特征是上皮細(xì)胞極性消失,轉(zhuǎn)換成運(yùn)動更為靈活、易于侵襲的間質(zhì)細(xì)胞,最直觀的表現(xiàn)就是其細(xì)胞形態(tài)常常發(fā)生改變[17]。本研究觀察到乙型肝炎病毒剪接特異性蛋白HBSP在肝癌細(xì)胞株Huh7中過表達(dá)可促進(jìn)TGFβ1誘導(dǎo)的EMT現(xiàn)象,經(jīng)過5 ng/mL TGFβ1刺激24 h后,表達(dá)HBSP的慢病毒細(xì)胞株大部分發(fā)生明顯形態(tài)改變,表現(xiàn)為細(xì)胞之間連接變得松散,細(xì)胞間隙變寬,細(xì)胞變長呈梭形,即上皮源性的肝細(xì)胞呈現(xiàn)出典型的間質(zhì)細(xì)胞形態(tài)。為深入探討其機(jī)制,我們進(jìn)一步利用特異性抗體檢測同樣處理?xiàng)l件下EMT的一些標(biāo)志分子,可見相應(yīng)細(xì)胞上皮標(biāo)志物E-cad、Claudin-1、β-catenin表達(dá)量下降,而間質(zhì)標(biāo)志物N-cad表達(dá)量升高,且該現(xiàn)象可隨TGFβ1I1被特異性siRNA下調(diào)而逆轉(zhuǎn)。提示HBSP過表達(dá)Huh7細(xì)胞株受TGFβ1誘導(dǎo)發(fā)生EMT,可能與HBSP-TGFβ1I蛋白相互作用有關(guān)。E-cad是一種跨膜類型的糖蛋白,在胞漿內(nèi)通過α、β連接素(α、β-catenin)間接的與肌動蛋白骨架連接,對于穩(wěn)定細(xì)胞骨架和保持上皮細(xì)胞間連接起重要作用,E-cad水平的下降往往同時(shí)伴隨有細(xì)胞間連接分子Claudin-1和β-catenin的下調(diào)[18]。N-cad是間質(zhì)細(xì)胞的主要標(biāo)志物,在很多腫瘤細(xì)胞發(fā)生中都出現(xiàn)N-cad的上調(diào)[19]。
EMT發(fā)生往往導(dǎo)致腫瘤細(xì)胞黏附能力下降,遷移運(yùn)動能力增加,使得腫瘤細(xì)胞更易于離開原有位置,發(fā)生原位浸潤或者隨血行、淋巴等途徑轉(zhuǎn)移[17]。為了深入揭示HBSP-TGFβ1I1相互作用導(dǎo)致EMT如何影響肝癌細(xì)胞惡性生物學(xué)行為,我們進(jìn)一步做了侵襲小室實(shí)驗(yàn),發(fā)現(xiàn)在TGFβ1誘導(dǎo)下,HBSP過表達(dá)組與對照組相比,侵襲能力顯著增強(qiáng),而當(dāng)TGFβ1I1被特異性siRNA下調(diào),侵襲能力得以減弱,說明HBSP能通過與TGFβ1I1相互作用加強(qiáng)Huh7細(xì)胞的侵襲能力。劃痕實(shí)驗(yàn)也同樣說明了HBSP蛋白與TGFβ1I1相互作用加強(qiáng)了Huh7細(xì)胞的遷移能力。
綜上所述,我們認(rèn)為乙型肝炎病毒編碼的剪接特異性蛋白HBSP可借由與TGFβ1I1的相互作用,促進(jìn)肝癌細(xì)胞發(fā)生EMT,繼而促進(jìn)腫瘤侵襲轉(zhuǎn)移。本研究所獲得的結(jié)果有助于闡明HBV病毒成分導(dǎo)致肝癌轉(zhuǎn)移的具體機(jī)制,并為抑制HBV相關(guān)肝癌的侵襲轉(zhuǎn)移提供新思路。
[1] Wu HL, Chen PJ, Tu SJ, et al. Characterization and genetic analysis of alternatively spliced transcripts of hepatitis B virus in infected human liver tissues and transfected HepG2 cells[J]. J Virol, 1991, 65(4):1680-1686.
[2] Soussan P, Tuveri R, Nalpas B, et al. The expression of hepatitis B spliced protein (HBSP) encoded by a spliced hepatitis B virus RNA is associated with viral replication and liver fibrosis[J]. J Hepatol, 2003, 38(3):343-348. DOI:10.1016/S0168-8278(02)00422-1
[3] Mancini-Bourgine M, Bayard F, Soussan P, et al. Hepatitis B virus splice-generated protein induces T-cell responses in HLA-transgenic mice and hepatitis B virus-infected patients[J]. J Virol, 2007, 81(10):4963-4972. DOI:10.1128/JVI.02619-06
[4] Chen WN, Chen JY, Jiao BY, et al. Interaction of hepatitis B spliced protein (HBSP) with cathepsin B promotes hepatoma cell migration and invasion[J]. J Virol, 2012, 86(24):13533-13541. DOI:10.1128/JVI.02095-12
[5] Chen JY, Chen WN, Liu LL, et al. Hepatitis B spliced protein (HBSP) generated by a spliced hepatitis B virus RNA participates in abnormality of fibrin formation and functions by binding to fibrinogen γ chain[J]. J Med Virol, 2010, 82(12):2019-2026. DOI:10.1002/jmv.21918
[6] Sciencepaper Online.Interaction of the hepatitis B spliced protein with transforming growth factor beta 1 induced transcript 1[EB/OL].[2015-7-13].http://www.paper.edu.cn/html/releasepaper/201507-118.
[7] Tumbarello DA, Turner CE. Hic-5 contributes to epithelial-mesenchymal transformation through a RhoA/ROCK-dependent pathway[J]. J Cellular Physiol, 2007, 211(3):736-47. DOI:10.1002/jcp.20991
[8] Soussan P, Garreau F, Zylberberg H, et al.Invivoexpression of a new hepatitis B virus protein encoded by a spliced RNA[J]. J Clin Investigat, 2000, 105(1):55-60. DOI:10.1172/JCI8098
[9] Chen JY, Chen WN, Jiao BY, et al. Hepatitis B spliced protein (HBSP) promotes the carcinogenic effects of benzo [alpha] pyrene by interacting with microsomal epoxide hydrolase and enhancing its hydrolysis activity[J]. BMC Cancer, 2014, 14:282. DOI:10.1186/1471-2407-14-282
[10] Shibanuma M, Mashimo J, Mita A, et al. Cloning from a mouse osteoblastic cell line of a set of transforming-growth-factor-beta 1-regulated genes, one of which seems to encode a follistatin-related polypeptide[J]. Eur J Biochem, 1993, 217(1):13-19. DOI:10.1111/j.1432-1033.1993.tb18212.x
[11] Shibanuma M, Mashimo J, Kuroki T, et al. Characterization of the TGF beta 1-inducible hic-5 gene that encodes a putative novel zinc finger protein and its possible involvement in cellular senescence[J]. J Biol Chem, 1994, 269(43):26767-26774. DOI:10.1007/s00109-010-0608-3
[12] Dabiri G, Tumbarello DA, Turner CE, et al. Hic-5 promotes the hypertrophic scar myofibroblast phenotype by regulating the TGF-beta1 autocrine loop[J]. J Invest Dermatol, 2008, 128(10):2518-2525. DOI:10.1038/jid.2008.90
[13] Tumbarello DA, Turner CE. Hic-5 contributes to epithelial-mesenchymal transformation through a RhoA/ROCK-dependent pathway[J]. J Cellular Physiol, 2007, 211(3):736-747. DOI:10.1002/jcp.20991
[14] Pignatelli J, Tumbarello DA,Schmidt RP, et al. Hic-5 promotes invadopodia formation and invasion during TGF-β-induced epithelial-mesenchymal transition[J]. J Cell Biol, 2012, 197(3):421-437. DOI:10.1083/jcb.201108143
[15] Xu Z, Shen MX, Ma DZ, et al. TGF-beta1-promoted epithelial-to-mesenchymal transformation and cell adhesion contribute to TGF-beta1-enhanced cell migration in SMMC-7721 cells [J]. Cell Res, 2003, 13(5):343-350. DOI:10.1038/sj.cr.7290179
[16] Calvisi DF, Pascale RM, Feo F. Epidermal growth factor-like repeats and discoidin I-like domains 3:a multifaceted oncoprotein at the crossroad of MAPK and TGF-beta pathways in human hepatocellular carcinoma[J]. Transl Cancer Res, 2016, 5(2):103-109. DOI:10.21037/tcr.2016.03.09
[17] Polyak K, Weinberg RA. Transitions between epithelial and mesenchymal states:acquisition of malignant and stem cell traits[J]. Nat Rev Cancer, 2009, 9(4):265-273. DOI:10.1038/nrc2620
[18] Tepass U, Truong K, Godt D, et al. Cadherins in embryonic and neural morphogenesis[J]. Nat Rev Mol Cell Biol, 2000, 1(2):91-100. DOI:10.1038/35040042
[19] An HT, Yoo S, Ko J.α-Actinin-4 induces the epithelial-to-mesenchymal transition and tumorigenesis via regulation of Snail expression and β-catenin stabilization in cervical cancer[J]. Oncogene, 2016, 35(45):5893-5904.DOI:10.1038/onc.2016.117
Lin Xu, Email:linxu@mail.fjmu.edu.cn
Hepatitis B spliced protein triggers TGFβ1-induced epithelial-mesenchymal transition via interaction with transforming growth factor beta-1-induced transcript 1 protein
CHEN Wan-nan, HUANG Jun-gao, LIANG Fei-fei, YAN Xiao-li, XUAN Dan-dan, LIN Xu
(KeyLaboratoryofMinistryofEducationforGastrointestinalCancer,KeyLaboratoryofFujianProvinceforTumorMicrobiology,SchoolofBasicMedicalSciences,FujianMedicalUniversity,Fuzhou350122,China)
To investigate the TGFβ1-induced epithelial-mesenchymal transition (EMT) of Huh7 hepatoma cells caused by interaction of hepatitis B spliced protein (HBSP) with transforming growth factor beta-1-induced transcript 1 protein (TGFβ1I1), coding region of HBSP was cloned into lentiviral expression vector. Huh7 hepatoma cells were infected by recombinant lentivirus packaged in 293T cells. Stable cell lines expressing HBSP or control cells were selected by puromycin. Cells were incubated with 5 ng/mL TGFβ1 for 24 h, and observed under contrast-phase microspcope. Then the whole cell lysates were collected for western blot analysis using specific antibodies against EMT markers including E-cadherin, N-cadherin, Claudin-1 and β-catenin. To evaluate the effects of HBSP-TGFβ1I1 interaction on EMT, TGFβ1-induced EMT marker transition, as well as cell invasion and migration were explored after knocking down of TGFβ1I1 by siRNA. Results showed that Huh7 cell lines expressing HBSP (Huh7-HBSP-flag-HIV) and control cell lines (Huh7-flag-HIV) were successfully established. Huh7-HBSP-flag-HIV cells lost their pebble-like shape and tight cell-cell adhesion and transformed into the mesenchymal-like cells in the presence of TGFβ1. Decreased expression level of epithelial marker of E-cadherin, Claudin-1, β-catenin, increased expression level of mesenchymal marker of N-cadherin, and enhanced migration and invasion abilities were observed in Huh7-HBSP-flag-HIV cells as compared to the control cells. Moreover, the changes of EMT markers and metastasis abilities of Huh7-HBSP-flag-HIV cells could be reversed when TGFβ1I1 was knocked down by siRNA. In conclusion, HBSP could promote hepatoma cell migration and invasion by triggering EMT via interaction with TGFβ1I1. Our findings highlight new insights for HBSP-induced HCC progression.
hepatitis B virus; RNA splicing; transforming growth factor beta-1-induced transcript 1 protein; epithelial-mesenchymal transition; metastasis
10.3969/j.issn.1002-2694.2017.04.003
國家自然科學(xué)基金青年基金項(xiàng)目 (No. 81201293);福建省衛(wèi)生廳醫(yī)學(xué)創(chuàng)新課題(No.2012-CX-14);福建省高校杰出青年科研人才培育計(jì)劃(No. JA11104);國家衛(wèi)生和計(jì)劃生育委員會共建科學(xué)研究基金-福建省衛(wèi)生教育聯(lián)合攻關(guān)計(jì)劃(No. WKJ-FJ-29);福建省高等學(xué)校新世紀(jì)優(yōu)秀人才支持計(jì)劃(No.JA13129);福建省衛(wèi)生系統(tǒng)中青年骨干人才培養(yǎng)項(xiàng)目(No. 2014-ZQN-ZD-25)聯(lián)合資助
林 旭:Email:linxu@mail.fjmu.edu.cn
福建醫(yī)科大學(xué)基礎(chǔ)醫(yī)學(xué)院,消化道惡性腫瘤省部共建教育部重點(diǎn)實(shí)驗(yàn)室,福建省腫瘤微生物學(xué)重點(diǎn)實(shí)驗(yàn)室,福州 350122
Supported by the National Natural Science Foundation of China (No. 81201293), the Fujian Provincial Medical Innovation Project (No. 2012-CX-14), the Outstanding Young Researchers Training Plan of Fujian Province (No. JA11104), the Joint Research Program of Health and Planning Committee and Education Department of Fujian (No. WKJ-FJ-29), the New Century Talents Supporting Plan of Fujian Education Department (No. JA13129), and the Medical Elite Cultivation Program of Fujian (No. 2014-ZQN-ZD-25)
R373
A
1002-2694(2017)04-0305-07
2016-11-16 編輯:梁小潔
猜你喜歡
天津醫(yī)科大學(xué)學(xué)報(bào)(2019年3期)2019-08-13 06:53:08
中成藥(2016年8期)2016-05-17 06:08:14
癌癥進(jìn)展(2016年12期)2016-03-20 13:16:17
罕少疾病雜志(2016年5期)2016-03-11 16:34:44
吉林大學(xué)學(xué)報(bào)(醫(yī)學(xué)版)(2015年1期)2015-12-17 07:47:28
腫瘤預(yù)防與治療(2015年1期)2015-09-26 07:26:20
中國當(dāng)代醫(yī)藥(2015年16期)2015-03-01 02:03:11
中國醫(yī)藥導(dǎo)報(bào)(2015年26期)2015-02-28 22:07:59
肝膽胰外科雜志(2015年4期)2015-02-27 11:12:34
肝膽胰外科雜志(2015年4期)2015-02-27 11:12:24
