盆底器官脫垂患者子宮骶韌帶組織LC3Ⅱ、COLⅠ和COL Ⅲ的表達變化
2017-06-05 15:01:05王紅南方
山東醫藥 2017年19期
王紅,南方
(鄂州市中心醫院,湖北鄂州436099)
盆底器官脫垂患者子宮骶韌帶組織LC3Ⅱ、COLⅠ和COL Ⅲ的表達變化
王紅,南方
(鄂州市中心醫院,湖北鄂州436099)
目的 觀察盆底器官脫垂(POP)患者子宮骶韌帶組織中自噬相關蛋白微管相關蛋白1輕鏈3Ⅱ(LC3Ⅱ)、Ⅰ型膠原(COLⅠ)和Ⅲ型膠原(COLⅢ)的表達變化,并探討自噬與POP的關系。方法 輕度POP(POPⅠ~Ⅱ度)14例(A組)、重度POP(POPⅢ~Ⅳ級)17例(B組)、無POP者17例(C組),在子宮全切術術中均留取子宮骶韌帶組織,采用Western blotting法檢測各組子宮骶韌帶組織LC3、COLⅠ和COL Ⅲ。采用Pearson相關性分析方法分析LC3、COLⅠ和COL Ⅲ的相關性。結果 A、B、C組LC3蛋白相對表達量分別為30.41±2.76、24.28±2.61、38.15±2.19,COLⅠ蛋白相對表達量分別為30.41±2.76、24.28±2.61、38.15±2.19,COLⅢ蛋白相對表達量分別為31.50±2.15、26.00±2.10、37.29±2.13,組間兩兩比較,P均<0.05。Pearson相關性分析顯示,輕度POP患者子宮骶韌帶組織中LC3Ⅱ與COLⅠ和COLⅢ均呈正相關(r分別為0.736 2,0.781 9;P均<0.01);重度POP患者子宮骶韌帶組織中LC3Ⅱ與COLⅠ和COLⅢ均呈正相關(r分別為-0.865,-0.709 4;P均<0.01)。結論 POP患者子宮骶韌帶組織LC3、COLⅠ和COL Ⅲ均呈低表達。LC3、COLⅠ和COL Ⅲ可能與POP的發生有關。
盆底器官脫垂;子宮骶韌帶;微管相關蛋白1輕鏈3Ⅱ;Ⅰ型膠原;Ⅲ型膠原
盆底器官脫垂(POP)主要發生在于經產婦[1],發病率較高[2,3]。流行病學調查發現,POP的發病主要與分娩、年齡、肥胖、慢性腹壓增加及絕經等因素相關[4]。但目前關于POP發病的具體分子機制仍不明確。自噬是生物體在長期進化中形成的一種自我保護機制,本質上是一種細胞促存活機制。微管相關蛋白1輕鏈3Ⅱ(LC3Ⅱ)是自噬體膜上的標志性蛋白,Ⅰ型膠原(COLⅠ)和Ⅲ型膠原(COLⅢ)是自噬標志蛋白。目前關于自噬在POP中的作用機制相關報道較少。我們觀察了POP患者子宮骶韌帶組織中LC3Ⅱ、COLⅠ和COLⅢ的表達變化,并探討自噬與POP的關系。現將結果報告如下。
1 資料與方法
1.1 臨床資料 選擇2015年1月~2016年1月間在鄂州市中心醫院行子宮全切術的住院患者48例,其中31例因POP行子宮全切術,因子宮腺肌癥、子宮肌瘤或功能性子宮出血等良性疾病行子宮全切術者17例;48例患者均為陰道分娩,均已絕經,既往無激素使用史,無明顯盆腔炎癥,無糖尿病或高血壓等疾病,無各種慢性疾病。根據國際尿控協會(ICS)1996年公布的POP-Q分類法將48例患者分為輕度POP(POPⅠ~Ⅱ度)者14例(A組)、重度POP(POPⅢ~Ⅳ級)者17例(B組)、無POP者17例(C組)。A組年齡(52.50±5.49)歲,BMI(22.57±3.35)kg/m2,分娩次數(2.40±1.18)次;B組年齡(51.88±3.90)歲,BMI(22.94±3.29) kg/m2,分娩次數(2.59±1.33)次,C組年齡(51.35±5.57)歲,BMI(22.71±3.18) kg/m2,分娩次數(2.41±1.18)次。三組年齡、陰道分娩次數、BMI等資料具有可比性。
1.2 各組子宮骶韌帶組織LC3、COLⅠ和COL Ⅲ檢測 三組均行子宮全切術,術中留取距離宮頸約0.5 cm處宮骶韌帶組織,長約0.5 cm。采用Western blotting法檢測各組子宮骶韌帶組織LC3、COLⅠ和COL Ⅲ。取各組組織,加入細胞裂解液充分裂解,離心后取上清,采用BCA試劑盒檢測蛋白濃度。配膠(分離膠10%)、上樣、電泳、轉膜(PVDF膜)、封閉、一抗孵育、二抗孵育。洗膜后ECL顯色液顯色。方正掃描儀掃描條帶,運用Quantity One軟件對條帶進行灰度分析,以LC3、COLⅠ和COL Ⅲ與GAPDH條帶的相對灰度值表示LC3、COLⅠ和COL Ⅲ蛋白的相對表達量。
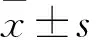
2 結果
三組子宮骶韌帶組織LC3、COLⅠ和COL Ⅲ蛋白的相對表達量比較見表1。
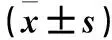
表1 三組子宮骶韌帶組織LC3、COLⅠ和COL Ⅲ蛋白的相對表達量比較
注:與C組比較,#P<0.05;與B組比較,*P<0.05。
Pearson相關性分析顯示,輕度POP患者子宮骶韌帶組織中LC3Ⅱ與COLⅠ和COLⅢ均呈正相關(r分別為0.736 2、0.781 9,P均<0.01);重度POP患者子宮骶韌帶組織中LC3Ⅱ與COLⅠ和COLⅢ均呈正相關(r分別為-0.865、-0.709 4,P均<0.01)。
3 討論
女性盆底器官主要是韌帶和肌肉固定于相應的位置,以至于不隨應力改變位置而影響功能,而子宮主骶韌帶是提供力學抗性的關鍵結構其主要成分是肌肉和膠原纖維。POP患者往往是這些支持結構發生了病變,其內的膠原蛋白等細胞外基質(ECM)成分發生了改變,導致結締組織松弛,最終引起POP。
自噬作為一種防御機制,在細胞缺氧、饑餓和生長因子缺乏等條件下,以及一些病理狀態下,通過對胞內物質的降解和循環利用來改變新陳代謝,使細胞得以生存,對維持細胞的存活有積極作用,是細胞在低氧環境下生存的可能機制,在本質上是一種細胞促存活機制[5],且具有雙向作用[6],適度的自噬有利于細胞的生存,過度自噬則會導致細胞死亡。Liu等[7]研究證實,提高自噬水平可抑制ECM的過度聚集,保護心肌細胞,促進膠原降解來抑制纖維化的進程[8]。而另一項風濕性關節炎疾病研究[9, 10]發現,自噬可促進滑膜內纖維母細胞的存活,抑制其凋亡,從而使得關節腔內的ECM過度分泌聚集,對受累關節造成永久性的損傷。自噬過程由一系列自噬相關蛋白(Atg蛋白)介導完成,這些蛋白質在自噬體形成的不同階段發揮作用。
LC3是自噬體膜上的標記蛋白,細胞內存在兩種形式的LC3蛋白:LC3Ⅰ和LC3Ⅱ。LC3蛋白在合成后其C端即被Atg4蛋白酶切割變成LC3Ⅰ,LC3Ⅰ散在分布于細胞漿內,當自噬體形成后,LC3Ⅰ和磷脂酰乙醇胺(PE)偶聯形成LC3 Ⅱ并定位于自噬體內膜和外膜。與其他一些定位于自噬性結構膜上的Atg蛋白不同(僅在自噬過程的某一階段發揮作用),LC3Ⅱ始終穩定地保留在自噬體膜上直到與溶酶體融合,因此被用來作為自噬體的標記,且LC3Ⅱ的水平在某種程度上反映了自噬體的數量[11],是目前最明確貫穿整個自噬過程,并能夠出現在晚期自噬溶酶體中的自噬相關蛋白,所以LC3-Ⅱ作為自噬的特異性標志物受到廣泛的肯定。本研究發現, A、B組患者子宮骶韌帶組織中LC3Ⅱ、COLⅠ和COLⅢ的蛋白的相對表達量低于C組,與Mao等[12]研究成果一致。COLⅠ和COLⅢ是成纖維細胞主要的細胞外基質成分之一,多數學者認為膠原合成的減少,分解的增加,膠原之間交聯減少,形態異常,COLⅠ和COLⅢ比例下降,這些都在POP的發生發展中具有重要的作用[13]。
相關性分析發現,輕度POP患者子宮骶韌帶組織中LC3Ⅱ與COLⅠ和COLⅢ均呈正相關;重度POP患者子宮骶韌帶組織中LC3Ⅱ與COLⅠ和COLⅢ均呈正相關。那么膠原的合成代謝是否受自噬的影響,目前還需更深層次的研究,但本研究結果發現的正相關提示了這種可能。膠原的合成代謝與多種細胞生長因子有關,其中轉化生長因子TGF-β1在調節膠原代謝過程中起關鍵作用,可通過多條途徑調節膠原合成和代謝[14]。Ghavami等[15]用TGF-β1誘導原代人心前房肌成纖維細胞發生纖維化,發現TGF-β1能夠通過激活細胞發生自噬,促進COLⅠ、纖連蛋白高表達,自噬抑制劑和敲除自噬相關基因均可抑制TGF-β1的促纖維化作用。有學者[16,17]用小鼠胚胎成纖維細胞建立細胞饑餓模型,TGF-β1可緩解細胞饑餓導致的細胞生長受限、線粒體損傷和細胞凋亡,進一步細胞嫁接模型實驗發現自噬可增強TGF-β1介導的成纖維細胞保護作用,說明自噬參與了TGF-β1介導的對腫瘤相關成纖維細胞的保護和促增殖過程。
綜上所述,POP患者子宮骶韌帶組織LC3、COLⅠ和COL Ⅲ均呈低表達。LC3、COLⅠ和COL Ⅲ在POP的發生發展中可能存在相互作用。但其具體作用機制尚有待于進一步研究。
[1] 郎景和,朱蘭. 女性盆底學[M]. 人民衛生出版社,2008:315-319.
[2] Weber AM, Richter HE. Pelvic organ prolapse[J]. Obstet Gynecol, 2005,106(3):615-634.
[3] 王靜怡,朱蘭. 成年女性盆腔臟器脫垂流行病學研究進展[J]. 實用婦產科雜志,2011,27(2):95-98.
[4] Medvedev AE, Piao W, Shoenfelt J, et al. Role of TLR4 tyrosine phosphorylation in signal transduction and endotoxin tolerance[J]. J Biol Chem, 2007,282(22):16042-16053.
[5] Mizushima N. Autophagy: process and function[J]. Genes Dev, 2007,21(22):2861-2873.
[6] Shintani T, Klionsky DJ. Autophagy in health and disease: a double-edged sword[J]. Science, 2004,306(5698):990-995.
[7] Liu S, Chen S, Li M, et al. Autophagy activation attenuates angiotensin II-induced cardiac fibrosis[J]. Arch Biochem Biophys, 2016,2016(590):37-47.
[8] Aranguiz-Urroz P, Canales J, Copaja M, et al. Beta(2)-adrenergic receptor regulates cardiac fibroblast autophagy and collagen degradation[J]. Biochim Biophys Acta, 2011,1812(1):23-31.
[9] Connor AM, Mahomed N, Gandhi R, et al. TNFalpha modulates protein degradation pathways in rheumatoid arthritis synovial fibroblasts[J]. Arthritis Res Ther, 2012,14(2):62.
[10] Shin YJ, Han SH, Kim DS, et al. Autophagy induction and CHOP under-expression promotes survival of fibroblasts from rheumatoid arthritis patients under endoplasmic reticulum stress[J]. Arthritis Res Ther, 2010,12(1):19.
[11] Kimura S, Fujita N, Noda T, et al. Monitoring autophagy in mammalian cultured cells through the dynamics of LC3[J]. Methods Enzymol, 2009,2009(452):1-12.
[12] Tomasek JJ, Gabbiani G, Hinz B, et al. Myofibroblasts and mechano-regulation of connective tissue remodelling[J]. Nat Rev Mol Cell Biol, 2002,3(5):349-363.
[13] Edwall L, Carlstrom K, Fianu JA. Markers of collagen synthesis and degradation in urogenital tissue and serum from women with and without uterovaginal prolapse[J]. Mol Hum Reprod, 2008,14(3):193-197.
[14] Sun Y, Zhu F, Yu X, et al. Treatment of established peritoneal fibrosis by gene transfer of Smad7 in a rat model of peritoneal dialysis[J]. Am J Nephrol, 2009,30(1):84-94.
[15] Ghavami S, Cunnington RH, Gupta S, et al. Autophagy is a regulator of TGF-beta1-induced fibrogenesis in primary human atrial myofibroblasts[J]. Cell Death Dis, 2015,2015(6):1696.
[16] Liu FL, Mo EP, Yang L, et al. Autophagy is involved in TGF-beta1-induced protective mechanisms and formation of cancer-associated fibroblasts phenotype in tumor microenvironment[J]. Oncotarget, 2016,7(4):4122-4141.
[17] 邰海鳳,陳姝彤,童曉文.自噬基因 Beclin1在宮頸癌中的研究進展[J].山東醫藥,2014,2014(9):97-99.
南方(E-mail: 847285973@qq.com)
10.3969/j.issn.1002-266X.2017.19.022
R711.2
B
1002-266X(2017)19-0072-03
2016-11-12)
