兔肝細(xì)胞生長(zhǎng)因子蛋白質(zhì)結(jié)構(gòu)及功能分析*
2017-06-26 03:11:32韓金霞梁兆光
黑龍江醫(yī)藥 2017年10期
韓金霞,梁兆光
(1.大慶油田總院心內(nèi)科,黑龍江 大慶 163000;2.哈爾濱醫(yī)科大學(xué)附屬第一醫(yī)院心內(nèi)七科,黑龍江 哈爾濱 150001)
肝細(xì)胞生長(zhǎng)因子 (Hepatocyte growth factor,HGF),是一種分泌型肝素親和糖蛋白,是心肌營(yíng)養(yǎng)因子和特異的促內(nèi)皮細(xì)胞生長(zhǎng)因子,能夠促進(jìn)新生血管的形成。HGF明顯促進(jìn)VEGF(血管內(nèi)皮生長(zhǎng)因子)的mRNA表達(dá),兩者在促血管生成方面具有協(xié)同作用[1]。文獻(xiàn)報(bào)道,在兔、小鼠缺血模型中,HGF能刺激新生血管生成,在體內(nèi)和體外試驗(yàn)中,HGF的血管生成活性被證實(shí)比血管內(nèi)皮生長(zhǎng)因子及堿性成纖維細(xì)胞生長(zhǎng)因子更為強(qiáng)大[2-3]。有報(bào)道稱[4],在大鼠心肌梗死模型中,HGF過(guò)度表達(dá)能刺激血管新生和側(cè)支血管形成。兔肝細(xì)胞生長(zhǎng)因子(rabbit hepatocyte growth factor,RABHGF) 是首次擴(kuò)增出的兔HGF基因序列,已在genbank中發(fā)表。本實(shí)驗(yàn)獲得了兩種全長(zhǎng)的兔HGF cDNA序列,利用軟件和生物信息學(xué)站點(diǎn)進(jìn)行分析,為今后利用兔HGF進(jìn)行基因研究提供更多的信息。
1 材料與方法
2007年本課題組通過(guò)5′-Race和3′-Race法和重組PCR法成功獲得了未知的兔肝細(xì)胞生長(zhǎng)因子的基因序列,獲得了兩種全長(zhǎng)的兔HGF cDNA序列,命名為兔HGF-1和兔HGF-2,已被Genbank收錄,編號(hào)分別為EU128740和FJ237417。利用計(jì)算機(jī)軟件Gene Runner和DNAStar及生物學(xué)信息站點(diǎn),對(duì)兔HGF cDNA序列進(jìn)行分析和比較,并對(duì)編碼的蛋白質(zhì)氨基酸序列進(jìn)行分析和預(yù)測(cè)。
2 結(jié)果
2.1 兔HGF的cDNA序列及其分析
兔HGF-1由2299個(gè)堿基對(duì)組成,編碼了762個(gè)氨基酸;兔HGF-2由2320堿個(gè)基對(duì)組成,編碼了769個(gè)氨基酸。人HGF基因cDNA序列為2184bp,編碼1條含728個(gè)氨基酸,具有4個(gè)糖基化位點(diǎn)、4個(gè)kringle環(huán),分子量為83KD的多肽鏈。通過(guò)比較不同的HGF的cDNA克隆后發(fā)現(xiàn),存在多種HGF的mRNA的轉(zhuǎn)錄本,經(jīng)過(guò)不同的拼接,導(dǎo)致了不同的HGF變種[5]。例如:人HGF是由1條含有728個(gè)氨基酸的單鏈前體蛋白裂解而來(lái)的異質(zhì)二聚體,單鏈HGF和異質(zhì)二聚體形式具有相等的生物活性[6]。兔HGF-2的cDNA在第854位堿基處較兔HGF-1的cDNA有1個(gè)21堿基片段(GAAGCCGTTATTTTGCAAGAG)的插入,使推導(dǎo)的兔HGF-2氨基酸序列在285位殘基處較兔HGF-1多7個(gè)氨基酸(GSRYFAR)。通過(guò)Gene Runner分析表明,在兔HGF-2氨基酸序列的285~292位置附近并不存在特殊的蛋白質(zhì)功能基序。將兔HGF-1的cDNA與兔HGF-1的cDNA核酸序列用DNAStar的Clustal V方法進(jìn)行比較發(fā)現(xiàn)二者相似性為99.8%;而用Jotun Hein和Clustal W方法比較二者相似性為100%。兔HGF-1與兔HGF-2的氨基酸序列進(jìn)行比較發(fā)現(xiàn)二者相似性為;用Jotun Hein方法比較為100%,用Clustal V方法比較為99.1%,用Clustal W方法比較為99.9%。因此,推斷兔HGF-1和兔HGF-2二者編碼蛋白質(zhì)的活性是相似或者相同的,或者推斷出兔HGF-1和兔HGF-2基因序列是兩種極為相似的變種。
兔HGF-1蛋白和兔HGF-2蛋白都缺失了FLPSS分子。FLPSS分子是pro-HGF的自然變種,因在其編碼區(qū)缺失15bp,故在Kringle 1區(qū)缺失了5個(gè)氨基酸。但這一缺失不影響其分泌后的加工,仍可被酶切為少于5個(gè)氨基酸的雙鏈HGF形式。與全長(zhǎng)的pro-HGF分子比較,F(xiàn)LPSS分子存在數(shù)量少,與肝素的親和力略有降低,其他性質(zhì)基本一致[6]。通過(guò)DNAStar軟件對(duì)genbank中公布的鼠HGF mRNA序列(GI:170172521,GI:220766,GI:220437)的比較發(fā)現(xiàn),鼠HGF mRNA(GI:170172521,GI:220766)兩個(gè)序列里含有FLPSS分子,鼠HGF mRNA (GI:220437)的序列也存在了FLPSS分子的缺失。
2.2 兔HGF蛋白質(zhì)序列的同源性分析及比較
利用核酸分析軟件DNAStar,將兔HGF的測(cè)序結(jié)果與已知的人HGF(2187bp)[GI:2171032]和鼠HGF(2189bp)[GI:220437]序列進(jìn)行了同源性分析。將兔HGF-1和人HGF的核酸序列進(jìn)行比較后發(fā)現(xiàn),二者同源性為89.3%,其推導(dǎo)的氨基酸序列同源性為91.4%。此外,兔HGF-1和鼠HGF核酸序列同源性為86.8%,推導(dǎo)的氨基酸序列同源性為92.0%。將兔HGF-2和人HGF的核酸序列進(jìn)行比較后發(fā)現(xiàn),二者同源性為89.1%,其推導(dǎo)的氨基酸序列同源性為90.9%。此外,兔HGF-2和鼠HGF核酸序列同源性為86.6%,推導(dǎo)的氨基酸序列同源性為91.6%。
2.3 蛋白質(zhì)各組成及疏水性分析
兔HGF-2序列片段全長(zhǎng)2320個(gè)堿基對(duì),含有2202個(gè)堿基對(duì)組成的最大開(kāi)放閱讀框架及118bp組成的3'非翻譯區(qū)。為了更加全面地統(tǒng)計(jì)各參數(shù),用expasy中的工具ProtParam,結(jié)果如下:氨基酸數(shù)目:769個(gè); 分子量:87483.1;等電點(diǎn):8.76;pH值:2.5;負(fù)電荷殘基總數(shù)(Asp+Glu):82;正電荷殘基總數(shù)(Arg+Lys):103;分子式:C3859H5993N1091O1122S58;原子總數(shù):12123;半衰期:30h;不穩(wěn)定系數(shù):31.73,該蛋白分類為穩(wěn)定蛋白;脂肪系數(shù):67.15;Grand average of hydropathicity (GRAVY)親水性評(píng)估:-0.606,該蛋白為親水蛋白。
2.4 蛋白質(zhì)功能基序分析
應(yīng)用Gene Runner對(duì)兔HGF-2蛋白進(jìn)行分析,分析提示:其分子內(nèi)部含有一特殊結(jié)構(gòu)RGD。RGD是促進(jìn)細(xì)胞粘附的蛋白質(zhì)中的特有結(jié)構(gòu),圍繞RGD的序列都是可變的,但RGD序列本身卻是高度保守的。識(shí)別RGD序列的整合素有α3β1、α5β1、IIb/β3等。1個(gè)cAMP-/cGMP依賴蛋白激酶磷酸化位點(diǎn);2個(gè)酪氨酸激酶(Tyr)磷酸化位點(diǎn);3個(gè)N-糖基化位點(diǎn);4個(gè)kringle環(huán);9個(gè)酪蛋白激酶II(CK2)磷酸化位點(diǎn);10個(gè)N-豆蔻酰化位點(diǎn);含12個(gè)蛋白質(zhì)激酶C(PKC)磷酸化位點(diǎn)。
2.5 蛋白質(zhì)亞細(xì)胞定位預(yù)測(cè)
通過(guò)工具PSORT進(jìn)行預(yù)測(cè),結(jié)果如下:細(xì)胞外質(zhì),包括細(xì)胞壁:13.0%;線粒體:21.7%;液泡結(jié)構(gòu):4.3%;胞核:47.8%。細(xì)胞質(zhì):8.7%;細(xì)胞骨架:4.3%。
2.6 蛋白質(zhì)信號(hào)肽及酶切位點(diǎn)預(yù)測(cè)
通過(guò)采用基于神經(jīng)網(wǎng)絡(luò)(NN)的算法和隱藏的馬爾可夫模型(HMM)預(yù)測(cè)兔HGF-2蛋白質(zhì)的信號(hào)肽及其切割位點(diǎn),結(jié)果表明,兔HGF-2蛋白具有一段由32個(gè)氨基酸組成的信號(hào)肽。信號(hào)肽概率:0.990;信號(hào)錨定概率:0.008。最大可能切割位點(diǎn):0.763 其切割位點(diǎn)可能位于32-33AA(AEG-QK)之間。見(jiàn)圖1。
2.7 蛋白酶切位點(diǎn)預(yù)測(cè)
通過(guò)工具Peptide Cutter進(jìn)行預(yù)測(cè),結(jié)果發(fā)現(xiàn),共有如下蛋白酶對(duì)兔HGF 2蛋白有酶切作用:精氨酸-C蛋白水解酶、Asp-N內(nèi)切酶、Asp-N內(nèi)切酶+N端亮氨酸、BNPS-Skatole、細(xì)胞凋亡蛋白酶1、2、3、4、5、6、7、8、9、10、高特異性糜蛋白酶、低特異性糜蛋白酶、梭菌蛋白酶、CNBr、腸激酶、顆粒酶B、凝血因子X(jué)a、甲酸、谷氨酰基肽鏈內(nèi)切酶、鹽酸羥胺、氧代苯甲酸、LysC、LysN、NTCB、胃蛋白酶(pH1.3)、胃蛋白酶(pH>2)、脯氨酸肽鏈內(nèi)切酶、蛋白酶K、葡萄球菌肽酶I、煙草蝕紋病毒酶、嗜熱菌蛋白酶和凝血酶,及胰蛋白酶等。
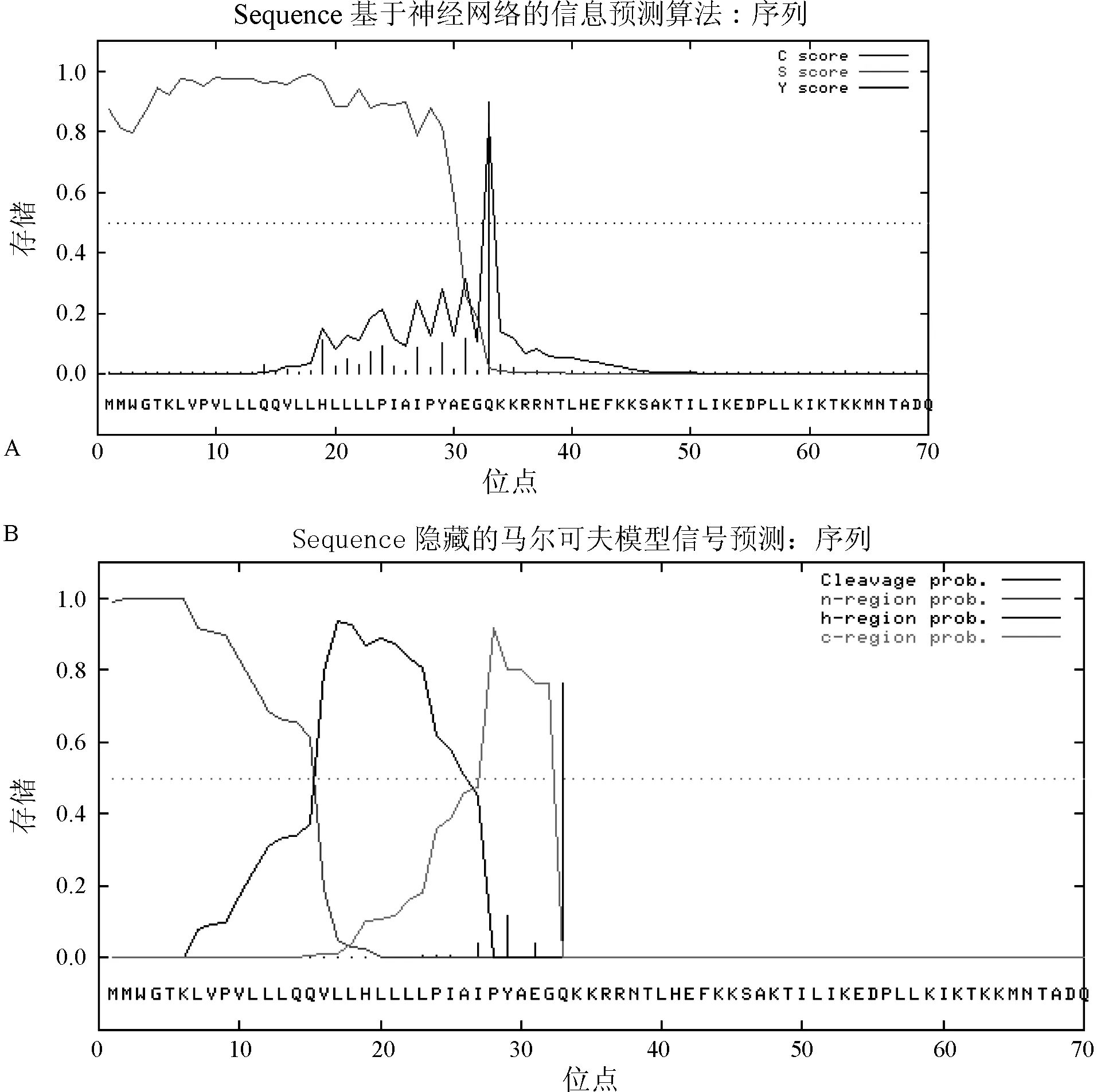
圖1 兔HGF1蛋白質(zhì)信號(hào)肽預(yù)測(cè) 注:A:NN(神經(jīng)網(wǎng)絡(luò))算法;B:HMM(馬爾可夫模型)算法
2.8 蛋白質(zhì)跨膜區(qū)預(yù)測(cè)
為了更直觀的顯示兔HGF-2蛋白是否具有跨膜區(qū),借助網(wǎng)站預(yù)測(cè)結(jié)果:ExpAA=19.92,該數(shù)值大于18;First 60 AAs=19.88,該數(shù)值大于10;PredHel=1;Topology=i7-29o,拓?fù)浣Y(jié)構(gòu)(跨膜螺旋)位于膜內(nèi)側(cè),氨基酸位置是7-29。見(jiàn)圖2。
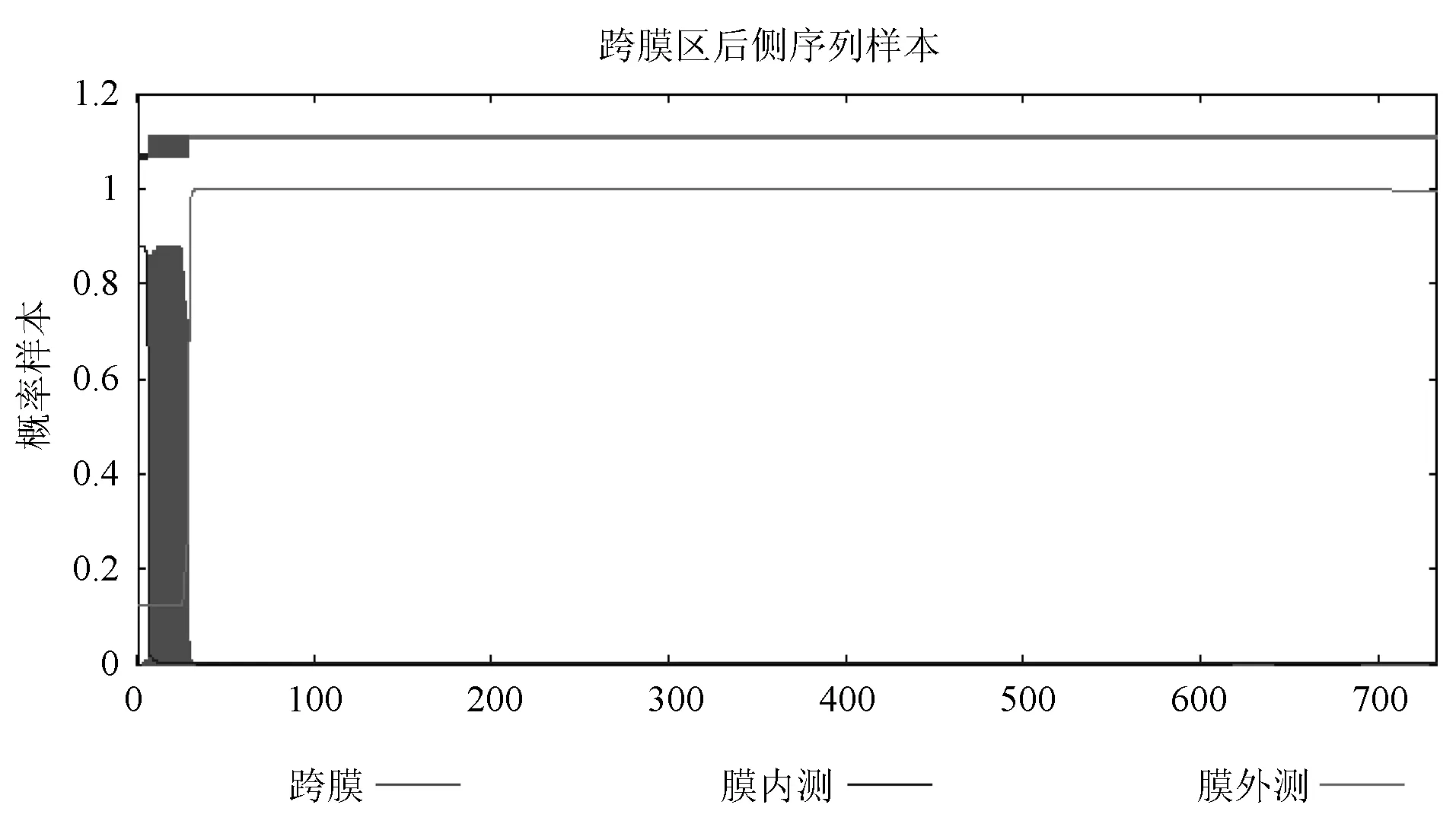
圖2 兔HGF-2蛋白質(zhì)跨膜區(qū)預(yù)測(cè)
3 討論
通過(guò)Gene Runner分析和DNAStar的Clustal V方法推斷兔HGF-2和兔HGF-1二者編碼蛋白質(zhì)的活性是相似或者相同的。因此,以兔HGF-2蛋白為例進(jìn)行比較全面的蛋白序列分析。兔HGF-2通過(guò)generuner分析表明,其具有3個(gè)糖基化位點(diǎn)、4個(gè)kringle環(huán),分子量為87KD的多肽鏈,兔HGF蛋白半衰期為30h,屬于穩(wěn)定的親水蛋白。預(yù)測(cè)兔HGF-2蛋白是否具有跨膜區(qū),ExpAA=19.92,該數(shù)值大于18,說(shuō)明這個(gè)蛋白結(jié)構(gòu)中更可能存在跨膜蛋白,或者說(shuō)有一個(gè)信號(hào)肽;First 60 AAs=19.88,該數(shù)值大于10,說(shuō)明這個(gè)蛋白結(jié)構(gòu)中在N-端可能存在信號(hào)肽;PredHel=1預(yù)測(cè)這個(gè)蛋白結(jié)構(gòu)中可能存在1個(gè)跨膜蛋白。總結(jié)比較已報(bào)道的不同的哺乳動(dòng)物的HGF,雖然它們均有促進(jìn)肝細(xì)胞生長(zhǎng)和增殖的功能,但在來(lái)源、分子量和性質(zhì)等方面有較大差異[7]。
HGF 是一種具有多種生物學(xué)功能的細(xì)胞因子,如促細(xì)胞分裂劑、促細(xì)胞運(yùn)動(dòng)劑和促形態(tài)形成劑等作用[8]。HGF通過(guò)結(jié)合到c-Met受體而作用于多種類型靶細(xì)胞,包括內(nèi)皮細(xì)胞,并能誘導(dǎo)非內(nèi)皮細(xì)胞釋放其他的內(nèi)皮細(xì)胞分裂原,是強(qiáng)有力的血管新生的刺激因子[9]。蔣逸風(fēng)等[10]實(shí)驗(yàn)研究提示,HGF可能主要通過(guò)促進(jìn)血管內(nèi)皮細(xì)胞增殖,加速內(nèi)皮系愈合而間接抑制血管平滑肌細(xì)胞的增殖,從而起到預(yù)防再狹窄的作用。
本研究表明,兔HGF基因可轉(zhuǎn)染至COS-7細(xì)胞并分泌表達(dá)兔HGF蛋白,對(duì)細(xì)胞具有促進(jìn)細(xì)胞生長(zhǎng)的生物學(xué)活性,從而利用兔HGF基因和已建立的兔心肌缺血模型進(jìn)行下一步缺血性心臟病的基因治療實(shí)驗(yàn)研究。
[1] NAKAMURA T,MIZUNO S,MATSUMOTO K,etal.Myocardial protection from ischemia/ reperfusion injury by endogenous and exogenous HGF [J].Clin Invest,2000,106 (12):1511-1519.
[2] MORISHITA R,SA KAKIM,YAMAMOTO K,etal.Impairment of collateral formation in Lp transgenic mice:therapeutic angiogenesis induced by human hepatocyte growth fator gene[J].Circulation,2001,105(10):1491-l496.
[3] TANIYAMA Y,MORISHITA R,AOKI M,etal.Therapeutic angiogenesis induced by human hepatocyte growth factor gene in rat and rabbit hindlimb ischemia models:preclinical study for treatment of peripheral arterial disease[J].Gene Ther,2001,8(1):181-189.
[4] TANIYAMA Y,MOFISHITA R,AOKI M,etal.Angiogensis and antifibrotic action by hepatocyte growth factor in cardiomyopathic hamsters[J].Circulation,2000,2(2):246-252.
[5] 俞水亮,楊復(fù)華.肝細(xì)胞生長(zhǎng)因子的分子生物學(xué)研究[J].生物工程學(xué)報(bào),2002,18(1):1-4.
[6] 張武.肝細(xì)胞生長(zhǎng)因子研究進(jìn)展[J].免疫學(xué)雜志,2001,17(3)125-128.
[7] 羅運(yùn)權(quán),吳孟超.肝細(xì)胞生長(zhǎng)因子[J].新消化病學(xué)雜志,1997,5(3)198-199.
[8] SEIDEL C,BORSET M.Role of hepatocyte growth factor and its recptor C-Met in multiple myeloma[J].Med Oncol,1998,15(3):145-153.
[9] MORISHITA R,AOKI M,HASHIYA N,etal.Therapeutic angiogenesis using hepatocyte growth factor[J].Curr Gene Ther,2004,4:199-206.
[10] 蔣逸風(fēng),林曉耘,陳雙紅,等.肝細(xì)胞生長(zhǎng)因子和血管內(nèi)皮細(xì)胞生長(zhǎng)因子對(duì)血管壁細(xì)胞增殖的影響[J].解放軍醫(yī)學(xué)雜志,2003,28(9):814-815.
