β-胡蘿卜素對脂多糖刺激巨噬細胞RAW264.7炎癥因子的影響及其機制①
2017-07-05 11:46:52張曉音張珊珊吉昱斌黃羽盛
中國免疫學雜志 2017年6期
張曉音 張珊珊 吳 旻 吉昱斌 黃羽盛 鄭 鑫
(吉林農業大學動物科學技術學院,長春130118)
β-胡蘿卜素對脂多糖刺激巨噬細胞RAW264.7炎癥因子的影響及其機制①
張曉音 張珊珊 吳 旻 吉昱斌 黃羽盛 鄭 鑫
(吉林農業大學動物科學技術學院,長春130118)
目的:探討β-胡蘿卜素對脂多糖(LPS)刺激巨噬細胞RAW264.7炎癥因子的影響及其機制。方法:采用5 μg/ml 的LPS刺激巨噬細胞24 h后用不同濃度的β-胡蘿卜素(20、40、80、160 μmol/L)處理細胞3 h,MTT法測細胞活性,熒光定量PCR測炎癥因子IL-1β、IL-6、TNF-α mRNA相對表達量,ELISA測炎癥因子IL-1β、IL-6、TNF-α的分泌量,Western blot測NF-κB p65蛋白相對表達量。5 μg/ml的LPS和不同濃度的NF-κB抑制劑PDTC(1、5、10 μg/ml)共同處理巨噬細胞24 h,Western blot測NF-κB p65蛋白相對表達量,熒光定量PCR、ELISA測炎癥因子的表達和分泌。比較LPS+PDTC組和LPS+PDTC+β-胡蘿卜素組炎癥因子和NF-κB p65蛋白相對表達的變化。結果:與LPS組相比,不同濃度的β-胡蘿卜素對LPS刺激的巨噬細胞的細胞活性有提升作用,對炎癥因子和NF-κB p65蛋白的表達有抑制作用;抑制NF-κB蛋白的表達可以抑制LPS刺激的巨噬細胞炎癥因子的分泌; LPS+PDTC+β-胡蘿卜素組對炎癥因子的抑制作用比LPS+PDTC組明顯,且差異顯著(P<0.05),但兩組對NF-κB p65蛋白的表達不明顯。結論:β-胡蘿卜素可以通過抑制NF-κB通路中NF-κB p65蛋白的表達抑制LPS刺激的巨噬細胞炎癥因子IL-1β、IL-6、TNF-α的分泌,且此通路不是唯一相關的通路。
β-胡蘿卜素;RAW264.7細胞;炎癥因子;NF-κB p65蛋白
炎癥是由激活的巨噬細胞釋放一系列炎癥因子所產生的以防御為主的一種免疫反應[1],炎癥因子的持續刺激會導致慢性炎癥的發生,與風濕性關節炎、哮喘、動脈粥樣硬化等疾病的產生密切相關[2,3]。類固醇和非類固醇藥物是傳統的抗炎藥物,被廣泛使用,但其對胃腸道、腎臟、中樞神經系統等存在極大的副作用[4],因此,尋求一種新型、高效、毒副作用小的抗炎藥物對慢性炎癥干預具有重要意義。β-胡蘿卜素是具有共軛多烯雙鍵結構的天然色素之一,具有抗氧化、抗癌、促進細胞間隙連接通訊等多種生物活性,在延緩機體衰老、預防疾病、提高機體免疫力等方面中發揮重要作用[5,6],現已被157個國家列入食品添加劑與飼料添加劑名錄。關于β-胡蘿卜素的研究大多基于其是維生素A的前體,在炎癥方面鮮有報道,本試驗擬通過細胞炎癥模型來研究β-胡蘿卜素對炎癥反應的作用。
巨噬細胞是參與炎癥反應的主要細胞之一,在脂多糖的刺激下,可誘導分泌IL-1β、IL-6、TNF-α等多種炎癥因子,進而出現臨床炎癥反應[7],而分泌的這些炎癥因子是檢測炎癥嚴重程度的量化指標[8]。所以,本試驗以LPS誘導RAW264.7巨噬細胞建立炎癥模型,研究β-胡蘿卜素對其炎癥因子分泌的影響及其作用機制,為β-胡蘿卜素的使用以及炎癥的治療提供理論基礎。
1 材料與方法
1.1材料
1.1.1細胞株 巨噬細胞RAW264.7購自武漢大學中國細胞典藏中心。
1.1.2主要試劑 β-胡蘿卜素(武漢星辰生物);RPMI1640培養基、胎牛血清(美國Gibco公司);LPS、MTT(美國Sigma公司);PDTC(上海碧云天);RNA Lyzol(上海ExCell公司);cDNA逆轉錄試劑盒(美國Gene Copoeia 公司);Brilliant Ⅱ SYBR Green QPCR Master Mix(美國 Agilent Technologies);引物(上海生工生物);RIPA裂解液(北京Solarbio);ELISA試劑盒(上海朗頓);BCA法蛋白定量試劑盒(上海申能博采);NF-κB抗體、β-actin抗體(美國CST);HRP標記的羊抗鼠/兔抗體(天津三箭);ECL試劑盒(美國Millipore)。
1.1.3主要儀器 高速冷凍離心機(德國Sigma);Thermo酶標儀(上海賽默飛世爾) ;熒光定量PCR儀、凝膠成像儀(美國Bio-Rad)。
1.2方法
1.2.1RAW264.7細胞培養 用含10%胎牛血清的RPMI1640培養基(含青霉素100 U/ml,鏈霉素100 μg/ml)于5% CO2、37℃ 培養箱中培養,1~2 d換液1次。待細胞生長至80%~90%時,根據不同的試驗目的,接種于96孔細胞板或6孔細胞板中繼續培養。
1.2.2MTT法檢測β-胡蘿卜素對RAW264.7細胞活性的影響 取對數生長期的細胞以8×104個/ml的密度接種于96孔細胞板中,每孔100 μl,置于5% CO2、37℃ 培養箱中培養。待細胞貼壁后,加入終濃度為0、20、40、80、160 μmol/L的β-胡蘿卜素(DMSO的最終質量分數為0.5%),每組8個復孔,培養3、6、12 h后分別棄去培養液,然后每孔加入0.5 mg/ml的MTT溶液100 μl,培養4 h后終止培養。最后,每孔加入100 μl DMSO,置于搖床上低速震蕩10 min,在酶標儀490 nm處測量其吸光值,計算不同濃度的β-胡蘿卜素組相對于空白組的細胞活性。
1.2.3熒光定量PCR、ELISA檢測β-胡蘿卜素、PDTC對LPS刺激RAW264.7細胞炎癥因子IL-1β、IL-6、TNF-α的影響 按照1.2.2中接種96孔細胞板的方法以細胞濃度為2×106個/ml接種RAW264.7細胞于6孔細胞板中,每孔2 ml,細胞貼壁后用5 μg/ml的LPS刺激細胞24 h,棄掉培養液后加入20、40、80、160 μmol/L的β-胡蘿卜素繼續培養3 h,或者5 μg/ml的LPS和1、5、10 μg/ml NF-κB抑制劑PDTC共同處理RAW264.7細胞24 h。①根據RNA Lyzol說明書提取細胞RNA,樣品在-80℃保存,并取部分樣品測RNA的質量,OD260/OD280在1.8~2.0之間,可用于下一步試驗。然后按照cDNA逆轉錄試劑盒的步驟將RNA轉錄為cDNA。最后熒光定量PCR,采用20 μl的反應體系,Master Mix 10 μl、上下游引物各0.5 μl、cDNA模板2 μl、染料1 μl 、ddH2O 6 μl,按照95℃ 預變性 5 min,95℃ 變性10 s,60℃ 退火75 s,40個循環進行PCR反應,每次試驗均設置18s作為內參。用2-ΔΔCt法計算炎性因子的相對表達量,引物序列表如表1所示[9]。②根據ELISA說明書,3 000 r/min、4℃離心20 min,用無菌管收集細胞上清,嚴格按照試劑盒步驟操作,繪制標準曲線,分別計算各組IL-1β、IL-6、TNF-α的含量(ng/L)。

表1 熒光定量PCR引物序列表
1.2.4Western blot法檢測PDTC、β-胡蘿卜素對LPS刺激RAW264.7細胞NF-κB p65蛋白表達的影響 取六孔細胞板中處理后的RAW264.7,預冷的PBS洗滌2次,加入含蛋白酶抑制劑的RIPA裂解液,在冰上裂解后用細胞刮輕輕刮下,12 000 r/min、4℃ 離心5 min,取上清液,BCA法測蛋白濃度。取30 μg總蛋白進行SDS-PAGE電泳,電轉印至PVDF 膜上,用3% 的BSA封閉3 h,一抗為鼠源的NF-κB p65抗體(1∶1 000),兔源的β-actin抗體(1∶5 000),4℃孵育過夜。1×TBST洗膜后分別加入相應的二抗:HRP標記的山羊抗鼠IgG(1∶2 000)和HRP標記的山羊抗兔IgG(1∶3 000),37℃ 孵育1 h。1×TBST洗膜后,感光,ECL顯色,用密度比值表示目的蛋白量。
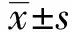
2 結果
2.1β-胡蘿卜素對RAW264.7細胞活性的影響 如圖1所示,β-胡蘿卜素作用RAW264.7細胞3 h,可以提升細胞活性,且在濃度為20、40、80 μmol/L時,差異極顯著(P<0.01);β-胡蘿卜素作用RAW264.7細胞6 h,濃度為160 μmol/L時,極顯著降低細胞活性(P<0.01);β-胡蘿卜素作用RAW264.7細胞12 h,濃度為80、160 μmol/L時,極顯著降低細胞活性(P<0.01),所以,選取不同濃度的β-胡蘿卜素作用細胞3 h作為本試驗的作用時間,在此濃度和時間范圍內,β-胡蘿卜素對細胞無明顯毒性,可作為試驗用安全濃度范圍。
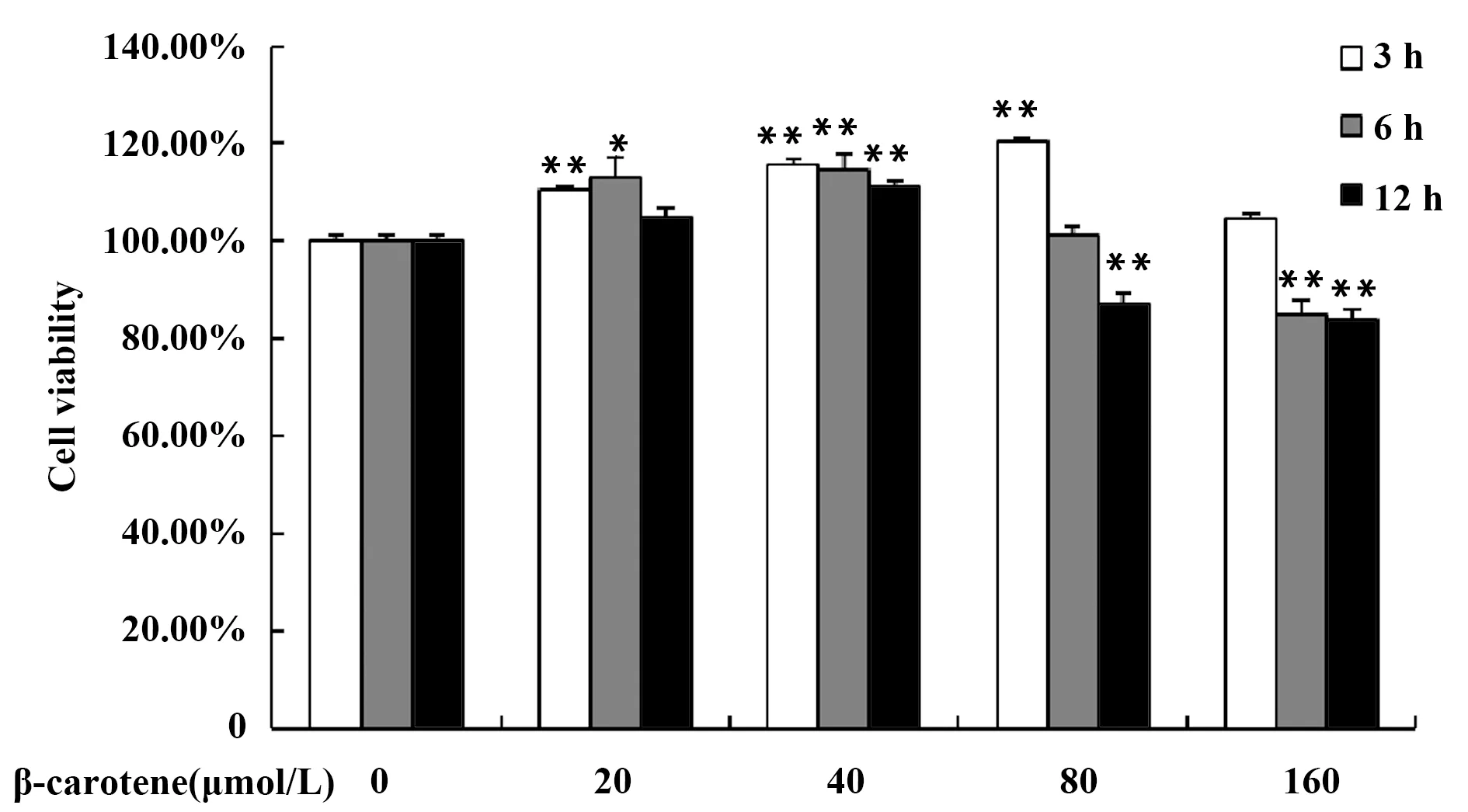
圖1 不同濃度的β-胡蘿卜素作用不同的時間對RAW 264.7細胞活性的影響Fig.1 Different concentration and different time of β-carotene on cell viability of RAW264.7 cellsNote:Compared with the control group(0 μmol/L),*.P<0.05,**.P<0.01.
LPS可以激活巨噬細胞,如圖2所示,5 μg/ml的LPS刺激RAW264.7細胞24 h,細胞活性略有上升,繼續用不同濃度的β-胡蘿卜素處理細胞3 h,細胞活性隨著β-胡蘿卜素濃度的升高而升高,有劑量依賴效應。由圖2可知,β-胡蘿卜素對LPS刺激后的巨噬細胞有增殖作用,基于細胞活性方面,此細胞模型建模成功,對細胞沒有毒性。
2.2β-胡蘿卜素對LPS刺激RAW264.7細胞炎癥因子IL-1β、IL-6、TNF-α的影響 LPS可激活巨噬細胞,誘導其分泌一系列的炎癥因子,炎癥因子的分泌是評價炎癥嚴重程度的一個量化指標,如圖3A所示,LPS誘導巨噬細胞炎癥因子IL-1β、IL-6、TNF-α mRNA的表達,用不同濃度的β-胡蘿卜素處理細胞后,炎癥因子mRNA相對表達量降低,且差異極顯著(P<0.01);如圖3B所示,LPS刺激巨噬細胞炎癥因子的分泌,不同濃度的β-胡蘿卜素可降低其分泌量,提示β-胡蘿卜素對炎癥因子的表達和分泌有一定的抑制作用。
2.3β-胡蘿卜素對LPS刺激RAW264.7細胞NF-κB p65蛋白表達的影響 NF-κB通路是炎癥通路中重要的通路之一,如圖4所示,用不同濃度的β-胡蘿卜素處理LPS刺激后的巨噬細胞,NF-κB p65蛋白表達量隨β-胡蘿卜素濃度的升高而降低,說明,β-胡蘿卜素在一定程度上可以抑制NF-κB p65蛋白的表達。但LPS組與對照組比,NF-κB p65蛋白表達基本沒變化,可能是細胞處理24 h,在長期饑餓狀態下,對照組中NF-κB通路已被激活。激活的NF-κB可結合到編碼炎癥因子基因啟動子區域的特定序列,進而產生對應的炎癥因子[1],所以,根據圖3、4推測,β-胡蘿卜素對炎癥因子的抑制作用可能是通過抑制NF-κB p65蛋白的表達來實現的。
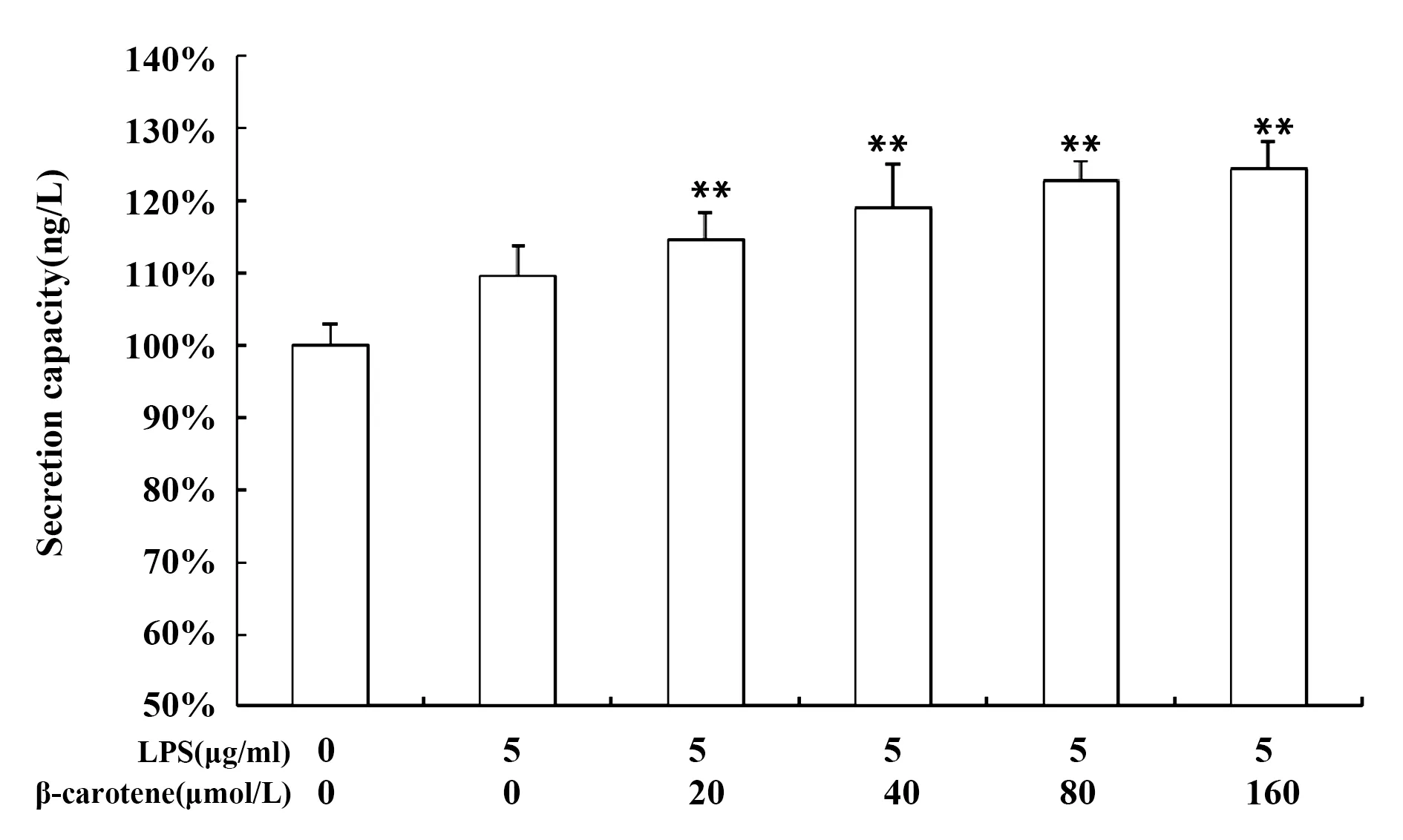
圖2 不同濃度的β-胡蘿卜素對LPS刺激的RAW264.7細胞活性的影響Fig.2 Different concentration of β-carotene on cell viability of LPS-induced RAW264.7 cellsNote:Compared with the LPS group,**.P<0.01.
2.4β-胡蘿卜素對LPS刺激RAW264.7細胞炎癥因子影響的分子機制 PDTC是常用的一種NF-κB抑制劑,能特異性的抑制NF-κB的活化,如圖5所示,不同濃度的PDTC與LPS共同作用細胞24 h,與LPS組相比,可顯著降低NF-κB p65蛋白的表達,驗證了其抑制NF-κB活化的作用,且在濃度為10 μg/ml時,抑制效果最好。
PDTC可以抑制NF-κB p65的表達,如圖6A、B所示,與LPS共同作用后,也可以抑制炎癥因子IL-1β、IL-6、TNF-α mRNA相對表達和分泌,說明,通過抑制NF-κB p65的表達可以抑制炎癥因子IL-1β、IL-6、TNF-α mRNA相對表達和分泌,因此,結合圖3、4的結果,β-胡蘿卜素可以通過抑制NF-κB p65蛋白的表達來抑制炎癥因子IL-1β、IL-6、TNF-α的分泌,β-胡蘿卜素對炎癥因子的抑制作用與NF-κB通路有關。
圖3 β-胡蘿卜素對LPS刺激RAW264.7炎癥因子的影響Fig.3 Different concentration of β-carotene on inflamma-tory factors of LPS-induced RAW264.7 cellsNote:A.The effects of β-carotene on the mRNA relative expression of inflammatory factors in LPS-induced RAW264.7 cells;B.The effects of β-carotene on the secretion capacity of inflammatory factors in LPS-induced RAW264.7 cells.Compared with the LPS group,*.P<0.05,**.P<0.01.
如圖7A、B所示,80 μmol/L的β-胡蘿卜素、10 μg/ml的PDTC以及兩者共同作用,均可以抑制LPS引起的炎癥因子IL-1β、IL-6、TNF-α mRNA的表達和分泌。對比LPS+PDTC組和LPS+PDTC+β-胡蘿卜素組發現,后者對炎癥因子IL-1β、IL-6、TNF-α的抑制作用更明顯,而且,如圖8所示,這兩組的NF-κB p65蛋白表達差異不顯著,這可能是β-胡蘿卜素的抑制作用還與其他通路有關,LPS+PDTC組和LPS+β-胡蘿卜素組的差異趨勢也驗證了這一點。
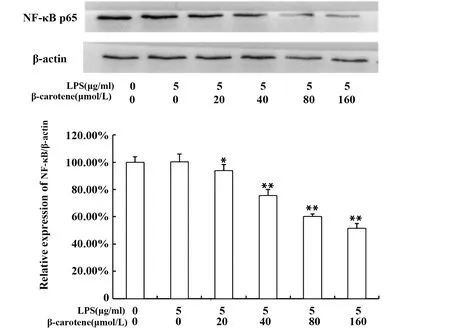
圖4 β-胡蘿卜素對LPS刺激RAW264.7細胞NF-κB p65蛋白表達的影響Fig.4 Different concentration of β-carotene on protein NF-κB p65 of LPS-induced RAW264.7 cellsNote: Compared with the LPS group,*.P<0.05,**.P<0.01.

圖5 PDTC對LPS刺激RAW264.7細胞NF-κB p65蛋白表達的影響Fig.5 Effect of different concentration of PDTC on prot-ein NF-κB p65 of LPS-induced RAW264.7 cellsNote: Compared with the LPS group,*.P<0.05,**.P<0.01.

圖6 PDTC對LPS刺激RAW264.7炎癥因子的影響Fig.6 Different concentration of PDTC on inflammatory factors of LPS-induced RAW264.7 cellsNote: A.The effects of PDTC on the mRNA relative expression of inflammatory factors in LPS-induced RAW264.7 cells;B.The effects of PDTC on the secretion capacity of inflammatory factors in LPS-induced RAW264.7 cells.Compared with the LPS group,*.P<0.05,**.P<0.01.

圖8 PDTC、β-胡蘿卜素對LPS刺激RAW264.7NF-κB p65蛋白比較圖Fig.8 Different concentration of PDTC and β-carotene on protein NF-κB p65 of LPS-induced RAW264.7 cellsNote: Compared with LPS group,*.P<0.05,**.P<0.01.
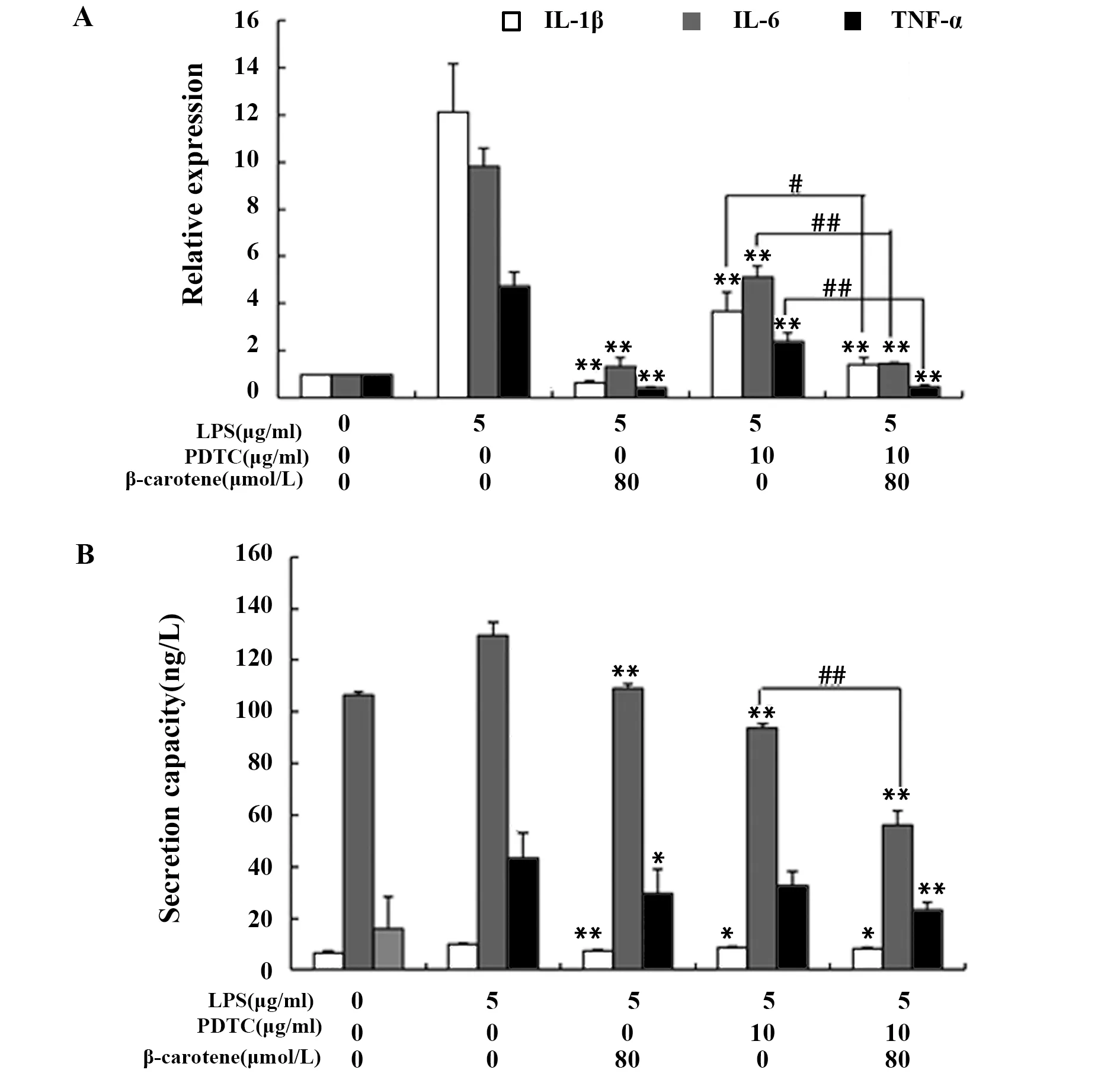
圖7 PDTC、β-胡蘿卜素對LPS刺激RAW264.7炎癥因子的影響Fig.7 Different concentration of PDTC and β-carotene on inflammatory factors of LPS-induced RAW2 64.7 cellsNote:A.The effects of PDTC and β-carotene on the mRNA relative expression of inflammatory factors in LPS-induced RAW264.7 cells;B.The effects of PDTC and β-carotene on the secretion capacity of inflammatory factors in LPS-induced RAW264.7 cells.Compared with the LPS group,*.P<0.05,**.P<0.01,compared with the LPS+PDTC group,#.P<0.05,##.P<0.01.
3 討論
炎癥是機體保護自身并消除有害刺激的過程,一般是有利的,分為急性炎癥和慢性炎癥,炎癥因子的持續刺激是導致慢性炎癥的主要原因,如高濃度的炎癥因子會誘導免疫細胞攻擊自身組織,嚴重的與糖尿病、風濕性關節炎、哮喘等疾病的產生密切相關,所以炎癥也是抗損傷和損傷的統一過程[10]。巨噬細胞在病原微生物引起的炎癥中發揮重要作用,LPS即可活化巨噬細胞并誘導其分泌多種促炎因子[11],最終會導致嚴重的組織損傷以及系統并發癥[12]。
炎癥因子的分泌是檢測炎癥嚴重程度的量化指標,本試驗以IL-1β、IL-6、TNF-α這三種炎癥因子作為檢測指標。TNF-α、IL-1β是炎癥介質級聯反應的始發因子[13],其中TNF-α是調節免疫細胞中重要的促炎細胞因子,通過產生IL-1β、IL-6誘導機體發熱、細胞凋亡等,進而誘導炎癥的發生,TNF-α含量的異常與多種疾病的發生密切相關。IL-6是調節免疫、炎癥反應等生理過程的多功能細胞因子,其表達受LPS和IL-1β的正向調節[14]。本試驗中,5 μg/ml的LPS刺激巨噬細胞后,IL-1β、IL-6、TNF-α的mRNA相對表達量明顯升高,即細胞已經處于炎癥的一個嚴重程度,當用不同濃度的β-胡蘿卜素處理后,這些炎癥因子開始降低,說明β-胡蘿卜素可以減輕LPS刺激的巨噬細胞引起的炎癥反應。
NF-κB通路在炎癥反應中發揮重要作用,正常狀態下,NF-κB以同二聚體或異二聚體存在胞質中,其p65亞基和抑制蛋白IκB結合,在受到外界刺激如LPS時,其抑制蛋白IκB發生磷酸化并降解,誘導p65亞基轉位至細胞核,與靶基因啟動子上的結合原件結合,誘導炎癥相關基因的轉錄[15,16]。本試驗中,LPS刺激的巨噬細胞用不同濃度的β-胡蘿卜素處理后,NF-κB p65蛋白的表達劑量依賴性降低,這可能是β-胡蘿卜素抑制炎癥因子的原因。
NF-κB抑制劑PDTC主要抑制NF-κB的活化,在本試驗中,不同濃度的PDTC可以抑制炎癥因子IL-1β、IL-6、TNF-α的mRNA相對表達和分泌,已有研究證實,IL-1β、IL-6、TNF-α基因的啟動子均存在NF-κB的結合位點,其表達受NF-κB活性的調控[17,18]。說明通過抑制NF-κB p65的表達的確可以抑制炎癥因子的分泌,間接說明,β-胡蘿卜素可以通過抑制NF-κB p65的表達從而抑制LPS刺激的巨噬細胞炎癥因子的分泌。
機體中炎癥信號通路受到多種復雜機制的調節,NF-κB通路只是重要的經典通路之一,大量的研究發現,LPS刺激的巨噬細胞可以激活MAPKs家族,巨噬細胞通過不同的MAPKs途徑調節炎癥反應[19]。本試驗中,LPS刺激的巨噬細胞,在PDTC、β-胡蘿卜素共同作用組和PDTC組,這兩組的NF-κB p65蛋白的表達基本無差異的情況下,共同作用組炎癥因子的表達和分泌較PDTC組顯著降低,說明,β-胡蘿卜素可能也通過其他的通路來抑制炎癥因子的分泌。
綜上所述,β-胡蘿卜素可以抑制LPS刺激的巨噬細胞RAW264.7炎癥因子的分泌,此抑制作用是通過抑制NF-κB p65蛋白的表達來實現的,與NF-κB通路有關,且此通路不是β-胡蘿卜素抑制炎癥因子的唯一通路。
[1] Ferrero-Miliani L,Nielsen OH,Andersen PS,etal.Chronic inflammation:importance of NOD2 and NALP3 in interleukin-1beta generation[J].Clin Exp Immunol,2007,147(2):227-235.
[2] Hwang SJ,Kim YW,Park Y,etal.Anti-inflammatory effects of chlorogenic acid in lipopolysaccharide-stimulated RAW 264.7 cells[J].Inflamm Res,2014,63(1):81-90.
[3] Ahn CB,Cho YS,Je JY.Purification and anti-inflammatory action of tripeptide from salmon pectoral fin by product protein hydrolysate[J].Food Chem,2015,168(1):151-156.
[4] Islam MN,Ishita IJ,Jin SE,etal.Anti-inflammatory activity of edible brown alga Saccharina japonica and its constituents pheophorbide a and pheophytin a in LPS-stimulated RAW 264.7 macrophage cells[J].Food Chem Toxicol,2013,55:541-548.
[5] 劉冬松,邵劍鋼,韓培濤,等.β-胡蘿卜素在高原功能食品中的應用[J].食品研究與開發,2015,36(13):147-149.
[6] Lin FM,Chen LR,Lin EH,etal.Compounds from wedelia chinensis synergisti-cally suppress androgen activity and growth in prostate cancer cells [J].Carcinogenesis,2007,28(12):2521-2529.
[7] Cotran RS,Kumar V,Collins T,etal.Robbins pathologic basis of disease[M].Philadelphia:WB Saunders Company,1998:1583-1584.
[8] Qin X,Qiu C,Zhao L.Lysophosphatidylcholine perpetuates macrophage polarization toward classically activated phenotype in inflammation[J].Cell Immunol,2014,289(1-2):185-190.
[9] Liu H,Beier JI,Arteel GE,etal.Transient receptor potential vanilloid 1 gene deficiency meliorates hepatic injury in a mouse model of chronic binge alcohol-induced alcoholic liver disease[J].Am J Pathol,2015,185(1):43-54.
[10] Martin P,Leibovich SJ.Inflammatory cells during wound repair:the good,the bad and the ugly[J].Trends Cell Biol,2005,15(11):599-607.
[11] Roger T,Delaloye J,Chanson AL,etal.Macrophage migration inhibitory factor deficiency is associated with impaired killing of gram-negative bacteria by macrophages and increased susceptibility to Klebsiella pneumonias sepsis[J].J Infect Dis,2013,207(2):331-339.
[12] Rosenblat JD,Cha DS,Mansur RB,etal.Inflamed moods:a review of the interactions between inflammation and disorders[J].Prog Neuropsychopharmacol Biol Psychiatry,2014,53:23-24.
[13] Malinin NL,Boldin MP,Kovalenko AV,etal.MAP3K-related kinase involved in NF-kappaB induction by TNF,CD95 and IL-1[J].Nature,1997,385(6616):540-544.
[14] 宋佳佳,白涵瑜,成 雪,等.血紅素氧合酶-1介導海參蛋白肽對脂多糖誘導RAW264.7巨噬細胞炎癥反應的抑制作用[J].現代食品科技,2016,32(4):1-7.
[15] Wan F,Lenardo MJ.The nuclear signaling of NF-kappaB:current knowledge, new insights,and future perspectives[J].Cell Res,2010,20(1):24-33.
[16] 吳 廣,符 平,周玉生,等.芹菜素抑制脂多糖誘導小鼠巨噬細胞分泌炎癥介質的分子機制[J].中國免疫學雜志,2015,31(6):753-757.
[17] 劉遠錦,田媛媛,劉 博,等.γ-谷維素對脂多糖誘導巨噬細胞RAW264.7炎癥因子表達的影響[J].食品科學,2015,36(19):238-243.
[18] Baud V,Derudder E.Control of NF-κB activity by proteolysis[J].Curr Top Microbiol Immunol,2011,349:97-114.
[19] Keshet Y,Seger R.The MAP kinase signaling cascades:a system of hundreds of components regulates a diverse array of physiological functions[J].Methods Mol Biol,2010,661(9):33-38.
[收稿2016-05-27 修回2016-07-20]
(編輯 許四平)
Effectsandmechanismofβ-caroteneoninflammatoryfactorsinLPS-inducedRAW264.7cells
ZHANGXiao-Yin,ZHANGShan-Shan,WUMin,JIYu-Bin,HUANGYu-Sheng,ZHENGXin.CollegeofAnimalScienceandTechnology,JilinAgriculturalUniversity,Changchun130118,China
Objective:To investigate the effects and mechanism of β-carotene on inflammatory factors(IL-1β,IL-6,TNF-α) in LPS-induced RAW264.7 cells.Methods:Firstly,RAW264.7 cells of being induced by 4(5 μg/ml)for 24 h were treated with different concentration of β-carotene(20,40,80,160 μmol/L)for 3 h.The cells viability was measured by MTT,the mRNA relative expression of IL-1β,IL-6,TNF-α was detected by fluorescence quantitative PCR,the secretion capacity of IL-1β,IL-6,TNF-α was detected by ELISA and the protein relative expression of NF-κB p65 protein was measured by Western blot.Secondly,RAW264.7 cells were induced by LPS(5 μg/ml) and different concentration of PDTC(1,5,10 μg/ml)for 24 h,NF-κB p65 protein was measured by Western blot and inflammatory factors were detected by fluorescence quantitative PCR and ELISA.Finally,compared the changes in the relative expression of inflammatory factors and NF-κB p65 protein between LPS+PDTC group and LPS+PDTC+β-carotene group.Results:Compared with the LPS-induced group,β-carotene could increase the cell viability of LPS-induced RAW264.7 cells and inhibied the relative expression of inflammatory factors and NF-κB p65 protein.Inhibited the relative expression of NF-κB p65 protein could reduce the relative expression of inflammatory factors.Compared with the LPS+PDTC group,LPS+PDTC+β-carotene group could inhibit the relative expression of inflammatory factors significantly(P<0.05).But,there was little difference about the relative expression of NF-κB p65 protein between this two groups.Conclusion:β-carotene inhibits the relative expression of inflammatory factors(IL-1β,IL-6,TNF-α) in LPS-induced RAW264.7 cells through inhibition of NF-κB p65 protein in NF-κB pathway,this pathway isn′t unique.
β-carotene;RAW264.7 cell;Inflammatory factor;NF-κB p65 protein
10.3969/j.issn.1000-484X.2017.06.007
①本文為吉林省科技廳科技成果轉化促進計劃(20150307021NY)。
張曉音(1991年-),女,在讀碩士,主要從事細胞免疫與動物育種方面的研究,E-mail:951770634@qq.com。
及指導教師:鄭 鑫(1965年-),女,博士,教授,主要從事細胞免疫與動物營養方面的研究,E-mail: zhengxinjilin@126.com。
S828
A
1000-484X(2017)06-0838-06
