人胚胎干細胞對肝癌HepG2細胞體外抑制作用的研究①
2017-07-05 11:47:07鄭良棟何雪梅霍本念劉夢楠
中國免疫學雜志 2017年6期
關鍵詞:肝癌
鄭良棟 何雪梅 張 婷 劉 潔 霍本念 劉夢楠 王 薛 馮 濤
(重慶醫科大學分子醫學與腫瘤研究中心,重慶400016)
·生物治療·
人胚胎干細胞對肝癌HepG2細胞體外抑制作用的研究①
鄭良棟②何雪梅 張 婷②劉 潔②霍本念②劉夢楠②王 薛 馮 濤②
(重慶醫科大學分子醫學與腫瘤研究中心,重慶400016)
目的:研究體外共培養情況下人胚胎干細胞H9對肝癌HepG2細胞的抑制作用。方法:建立人胚胎干細胞 (H9) 與肝癌HepG2細胞共培養體系,顯微鏡下觀察胚胎干細胞對腫瘤細胞生物學行為的影響,流式細胞儀檢測H9細胞對腫瘤細胞的凋亡與周期的影響,Transwell小室法檢測H9細胞對HepG2細胞侵襲和遷移的影響,基因芯片分析共培養后HepG2細胞全基因組的表達譜變化。結果:結果發現共培養過程中,肝癌HepG2細胞生長受到抑制,隨共培養時間延長,細胞數量逐漸減少,出現老化或凋亡跡象;流式細胞術檢測發現HepG2細胞凋亡率顯著增加,細胞周期被阻滯于G0/G1期;Transwell實驗發現HepG2細胞侵襲、遷移力均降低;基因芯片結果發現HepG2細胞全基因組表達譜發生了顯著變化,差異基因涉及多條信號通路。結論:人胚胎干細胞H9體外對人肝癌細胞HepG2有一定程度的抑制作用。
胚胎干細胞;肝癌;共培養;抑制;凋亡
人胚胎干細胞是從人早期胚胎的囊胚內細胞團中分離出來的多潛能干細胞,具有無限增殖性、自我更新性和維持未分化狀態等特性,在基礎研究和臨床應用方面具有廣闊的前景。
在腫瘤機制研究中,研究者發現腫瘤細胞尤其是腫瘤干細胞和胚胎干細胞在許多生物學行為上具有高度相似性,比如都具有無限增殖性、高表達端粒酶、侵襲、遷移性等,并且可從機體的免疫監視中逃逸,具有一些相同的信號轉導通路[1-3]。但是,二者在對腫瘤作用方面卻截然不同:腫瘤基質具有很強的致癌性,能夠體外誘導正常細胞癌變,然而胚胎干細胞微環境卻能夠使得癌細胞的惡性表型顯著改善,促進其向良性轉化[4]。
共培養是將兩種或多種細胞混合培養,使得一種細胞的形態功能能夠穩定表達并且維持相當長的時間[5]。細胞共培養技術在腫瘤的實驗性治療、干細胞分化誘導等多方面均具有廣泛應用。
為了進一步驗證人胚胎干細胞是否具有體外抗腫瘤效應。本研究選擇人胚胎干細胞系H9與肝癌HepG2細胞建立共培養模型,并觀察共培養過程中胚胎干細胞對肝癌細胞的細胞形態學、凋亡、細胞周期、侵襲、遷移等的影響,并嘗試利用基因芯片技術初步探討其作用機制。
1 材料與方法
1.1材料
1.1.1主要試劑 人胚胎干細胞系H9細胞購自北京賽貝生物技術有限公司;人肝癌HepG2細胞株由重慶醫科大學分子醫學與腫瘤研究中心保存;PSCeasy人多潛能干細胞培養基(北京賽貝生物技術有限公司);胎牛血清,DMEM培養基(HyClone,USA);hoechst33258(碧云天生物技術研究所);Transwell小室(Coster);Matrigel基質膠(Sigma公司)。
1.1.2主要儀器 CO2細胞培養箱(Thermo公司);-80℃超低溫冰箱(Thermo公司);-20℃普通冰箱(Thermo公司);倒置熒光顯微鏡(Olympus公司);YJ-1450型醫用超凈工作臺(華新器械公司);臺式離心機(湘儀離心機儀器公司);普通倒置顯微鏡(重慶光學儀器公司)。
1.2方法
1.2.1細胞培養 HepG2細胞用含10%胎牛血清、青霉素100 U/ml、鏈霉素100 μg/ml的DMEM完全培養基在37℃、5%CO2培養箱中培養,3~4 d傳代一次,取對數期細胞進行試驗。
人胚胎干細胞系H9,培養于Basement Membrane Matrix上,培養液為胚胎干細胞完全培養基。保持每天換液1次,當細胞匯合度達到80%時,用EDTA 在37℃下溫和消化5 min,然后離心細胞棄去上清,用新鮮的PSCeasy完全培養液重懸,以1∶5~1∶10的比例傳代。
1.2.2共培養過程中HepG2細胞形態學觀察 根據上述胚胎干細胞培養詳細方法,將H9細胞接種于6孔板內,待細胞匯合度達到50%(即細胞團塊約長滿6孔板底面積的一半)時,吸棄原干細胞專用培養基,用PBS溶液清洗細胞2遍。取對數生長期的肝癌HepG2細胞,以每孔5×105個接種于上述已經鋪有50%胚胎干細胞H9的6孔板中,每孔加入DMEM完全培養基3 ml于細胞培養箱內共培養。在共培養開始時,另外于6孔板設置相同細胞量的單獨培養的胚胎干細胞和肝癌細胞作為陰性對照。每隔24 h換液一次,以保證細胞生長營養充足。共培養細胞分別于24、48、72 h后于顯微鏡下觀察各組細胞的變化并拍照。
1.2.3肝癌細胞周期、凋亡檢測 共培養:將H9細胞接種于6孔板內,待細胞匯合度達到70%時,備用。將對數期生長狀態良好的HepG2細胞5×105個接種到6孔板適用的Transwell小室(0.4 μm)底部,用DMEM培養基培養到細胞貼壁。實驗分組:肝癌HepG2與干細胞H9共培養為實驗組(H9+HepG2),單獨培養的肝癌HepG2為空白對照組(HepG2)。Transwell小室用 PBS洗2遍,然后插入預先種有H9細胞的細胞培養板里,將各組細胞同時放入孵箱中培養到48 h。
共培養結束后,收集實驗組與對照組的肝癌HepG2細胞,用PBS洗2次,然后用預冷的70%乙醇固定細胞,PI染色流式細胞儀檢測細胞周期。
共培養結束后,收集實驗組與對照組的HepG2細胞,PBS洗2遍后,PBS重懸,上流式細胞儀,用AnnexinV-PI雙染法檢測細胞凋亡。
1.2.4肝癌細胞侵襲和遷移實驗 Matrigel包被:將Matrigel 基質膠從-20℃拿出,于4℃冰箱放置過夜解凍,然后用4℃預冷的無血清1640培養基稀釋至1 mg/ml 的濃度,取稀釋過的Matrigel 膠800 μl 加入到Transwell的上室,將Transwell小室放在37℃細胞孵箱溫育4~5 h,使上室膜上形成一層凝膠。將對數期生長狀態良好的HepG2細胞5×105個接種到6孔板適用的8.0 μm孔徑Transwell小室底部(侵襲實驗使用的Transwell小室預先經Matrigel包被,遷移實驗則不需Matrigel包被),用DMEM培養基培養到細胞貼壁,血清饑餓處理12 h。實驗組的Transwell小室用 PBS洗2遍,然后插入預先種有70%匯合度H9細胞的細胞培養板孔里;對照組的Transwell小室用 PBS洗2遍后則插入未接種H9的空白培養板孔里將各組細胞同時放入孵箱中培養24 h。
培養結束后,用醫用棉簽輕柔擦去上室表面細胞,然后用甲醇固定15 min。PBS潤洗2遍后,0.1%濃度的結晶紫溶液室溫下染色30 min,然后用PBS洗凈染液,將Transwell晾干,在倒置顯微鏡下觀察穿過小室膜的細胞數目并拍照記錄。每個小室可隨機選取5個不同的視野,數出視野里的細胞數目,求出平均值。
1.2.5基因芯片檢測肝癌細胞基因變化 細胞前期共培養處理同1.2.3的“共培養”項。
共培養48 h后,胰酶消化收集細胞,磷酸鹽緩沖液(PBS)洗滌1次后收集細胞,并用PBS液重懸細胞,并送往上海伯豪生物技術有限公司[生物芯片(上海)國家工程研究中心]進行基因芯片分析,用Affymetrix表達譜芯片測定樣品的全基因組表達情況。
2 結果
2.1共培養的生物學行為觀察 人胚胎干細胞H9與肝癌HepG2細胞共培養一段時間后,肝癌細胞生長受到抑制。如圖1所示,共培養0 h時,干細胞與肝癌細胞狀態均很好,24 h后,肝癌細胞生長開始受到抑制,細胞開始變稀疏,尤其是靠近干細胞團塊的肝癌細胞受到抑制而生長稀疏。隨著共培養時間的延長,癌細胞凋亡加劇,到72 h時,已經可見細胞數量明顯減少。且在共培養過程中胚胎干細胞因為受培養基、環境等因素影響,細胞極易脫落,即原來致密的細胞團塊開始疏松并且脫落漂浮,脫落后其原來生長過的空間短時間腫瘤細胞無法生長。且發現,共培養初始時覆蓋在胚胎干細胞表面的腫瘤細胞,最終繼續貼附著在干細胞表面而漂浮死亡。而單獨培養的H9和HepG2細胞,細胞生長和增殖均正常。
2.2共培養對腫瘤細胞凋亡的影響 共培養48 h后,AnnexinV-PI雙染法檢測細胞凋亡結果見圖2。與單獨培養的HepG2細胞相比,共培養的HepG2細胞,其凋亡率有所增加,具有顯著性差異(P<0.05)。從單獨培養時的5.21%增加到共培養時的30.14%。
2.3共培養對腫瘤細胞周期的影響 與單獨培養的HepG2細胞相比,共培養組的HepG2細胞在作用48 h后,G0/G1期細胞比率增加顯著,從49.80%增加到72.91%;G2/M期有所下降,從16.21%降到7.25%;S期從33.99%降到19.85%。可見,細胞停留在G0/G1期比率增加,表現出明顯的G0/G1期阻滯,其差異具有顯著性差異(P<0.05)。見圖3。
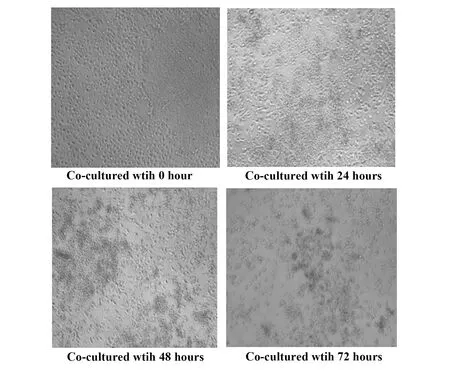
圖1 HepG2的細胞形態變化(×100)Fig.1 Morphological changes of HepG2 cells(×100)
2.4共培養對腫瘤細胞侵襲和遷移的影響 如圖4和表1所示,在侵襲和遷移實驗中,共培養組肝癌HepG2細胞穿膜細胞數均明顯少于對照組HepG2細胞(P<0.05),即共培養組肝癌HepG2細胞穿透Matrigel基底膜的侵襲力明顯低于對照組細胞,共培養組肝癌HepG2細胞穿透小室底膜的遷移能力也較對照組低,差異均有統計學意義(P<0.05)。
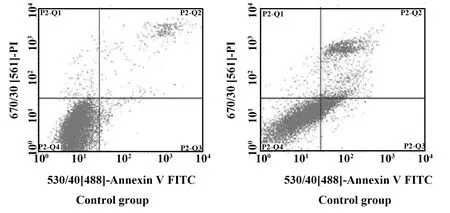
圖2 H9細胞對HepG2細胞凋亡的影響Fig.2 Effect of H9 on apoptosis of HepG2 cells

圖3 H9對HepG2細胞周期的影響Fig.3 Effect of H9 on cell cycle of HepG2 cellsNote: A.HepG2 cell line was cultured alone;B.HepG2 cell line was co-cultured with H9.
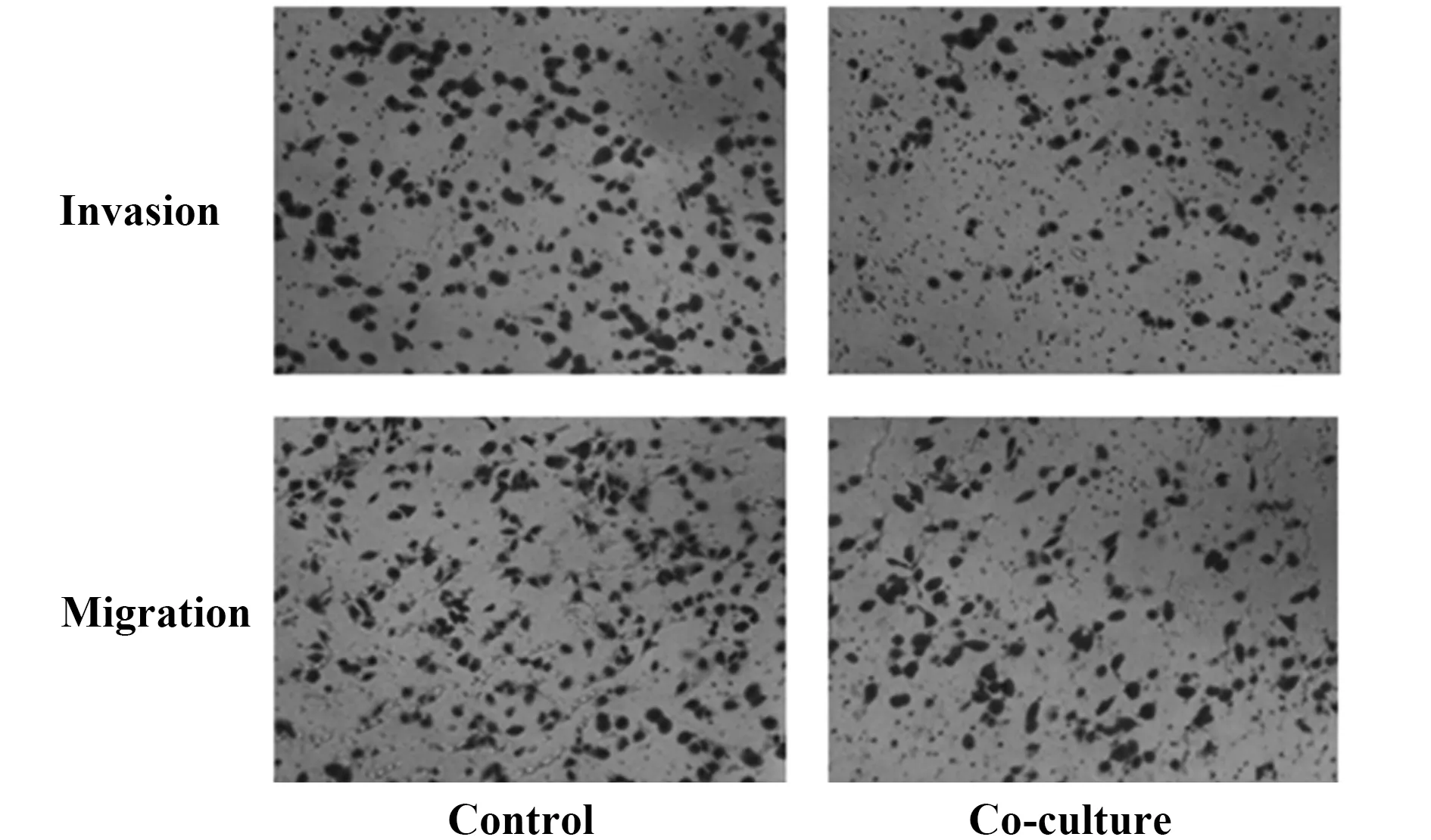
圖4 H9細胞對HepG2細胞侵襲和遷移的影響(×200)Fig.4 Effect of H9 on invasion and migration of HepG2 cells(×200)
2.5共培養對腫瘤細胞全基因組表達譜的影響 采用信號比值方法對芯片結果進行分析。針對掃描到的探針信號值實驗組/對照組。篩選2倍以上變化基因為差異基因。結果發現,差異的基因共15 588個。其中上調的基因6 640個,下調的基因8 948個。差異基因散點圖見圖5。
這些差異基因涉及多個方面,包括細胞周期調控、細胞信號轉導、細胞凋亡、細胞增殖、遷移、免疫應答等。其中凋亡相關基因有BCL-2、caspase3、caspase7、caspase8、caspase9、caspase10、AKT1、TRAF1等數百條,涉及多條凋亡調控通路。免疫相關基因也有IL-1、IL-2、IL-4、IL-8、CCL1、CCL16、CCL20、HLA-A、HLA-B、HLA-C等數百條基因,涉及B細胞、T細胞和NK細胞、免疫因子如干擾素、腫瘤壞死因子等參與的信號通路。由于涉及的差異基因數目和通路較多,只列舉出差異基因GO富集分析中部分我們感興趣的結果,見表2。
表1 HepG2細胞侵襲和遷移的數量

圖5 散點圖Fig.5 Plot scatter plotNote:s3.Co-culture group;s5.Control group.
3 討論
細胞共培養技術在腫瘤的實驗性治療、腫瘤的侵襲性[6-10]、體內基因表達的模擬等方面均具有重要意義[9-11]。本研究中我們采用共培養體系,研究體外環境中胚胎干細胞及其微環境對肝癌HepG2細胞的抑制作用,并探索其機制。首先,我們發現經與胚胎干細胞共培養的肝癌HepG2細胞發生了形態學變化,具體表現為癌細胞狀態不佳,細胞生長緩慢,部分細胞出現老化或凋亡跡象。為了進一步驗證胚胎干細胞的這種體外抑癌效應,我們使用了流式細胞儀檢測共培養后肝癌HepG2細胞的凋亡率和細胞周期變化,流式檢測結果與觀察到的現象一致。與單獨培養的HepG2細胞相比,共培養組的HepG2細胞處于G0/G1期細胞數目顯著增多,而G2/M、S期細胞數目減少,表現出明顯的G0/G1期阻滯,并且細胞凋亡率也有明顯的升高。可能的原因是胚胎干細胞生長過程中或其微環境中,存在一些細胞因子,可調控細胞周期和誘導肝癌細胞進入凋亡程序,并在某種程度上對腫瘤細胞基因重編程,使一些惡性表型轉化為良性[12,13]。
惡性腫瘤細胞的侵襲和遷移現象是腫瘤細胞有別于正常細胞的重要特性,也是臨床治療過程中腫瘤復發和轉移的主要原因之一[14]。因此,我們檢測了共培養條件下HepG2細胞的侵襲力和遷移力的變化。結果顯示,共培養組HepG2細胞的侵襲力和遷移力均明顯降低,表明胚胎干細胞及其微環境對肝癌HepG2細胞的侵襲和遷移能力也有抑制作用。
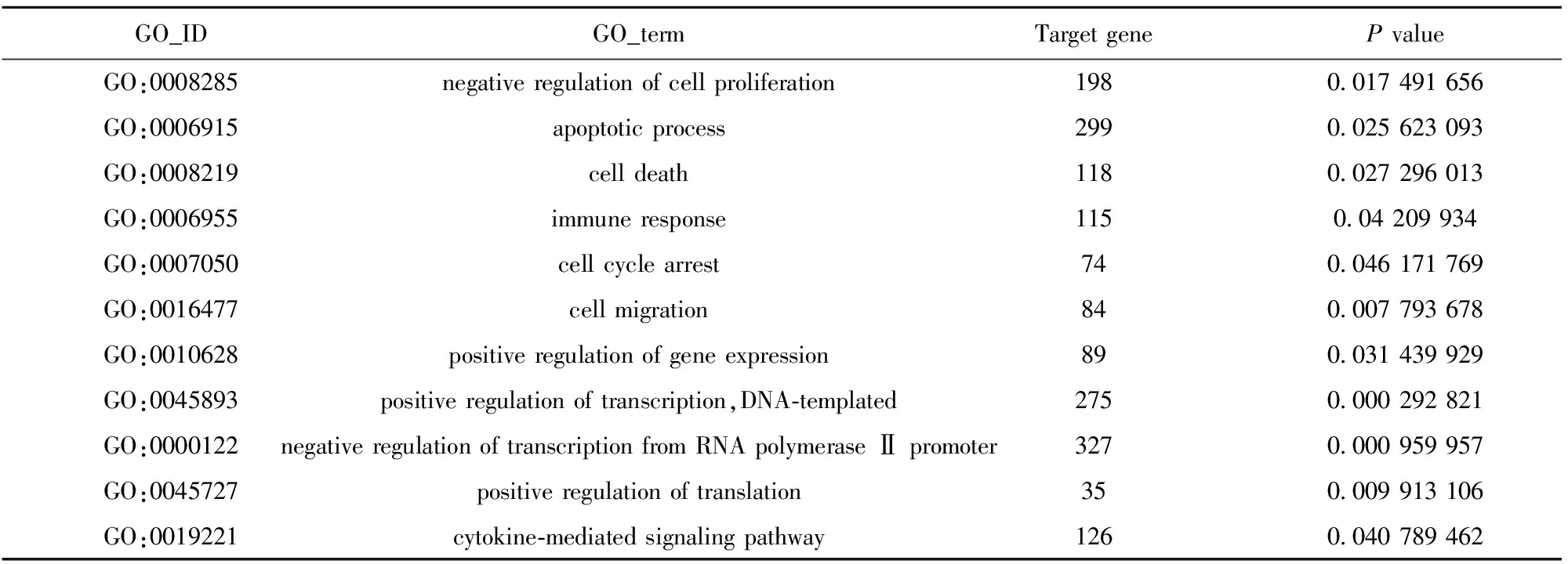
表2 GO分析結果
基因芯片技術的發展為大規模分析基因表達提供了工具[15,16]。我們通過基因芯片技術分析共培養后肝癌肝癌細胞HepG2全基因組的表達譜變化,共檢測出數萬個差異基因。進一步的差異基因結果富集分析也篩選出數百條GO-terms,其中涉及細胞周期調控、細胞信號轉導、細胞凋亡、細胞增殖、遷移、免疫應答等,從基因層面佐證了前述實驗中胚胎干細胞對肝癌HepG2細胞細胞生長、凋亡、周期、侵襲、遷移等的影響。Yaddanapudi等[17]研究證實,由胚胎干細胞誘導的抗腫瘤免疫并不是一種機體的“異物反應”,而有可能是由多種效應細胞和細胞因子參與構成的針對胚胎干細胞表面抗原的特異性免疫應答。而我們GO分析富集得出的GO 條目immune response下的115個免疫應答相關基因發生了顯著變化,涉及B細胞、T細胞和NK細胞、免疫因子如干擾素、腫瘤壞死因子等參與的信號通路。從基因層面證實了胚胎干細胞可能通過影響肝癌HepG2細胞的免疫應答等產生對HepG2細胞的抑制作用。我們猜測可能是共培養過程中,由肝癌細胞作為抗原刺激胚胎干細胞分化產生一些免疫細胞或免疫因子等,對肝癌細胞產生了免疫重編程作用。基因芯片雖然有大規模、高通量、可快速篩選基因等優點,但仍然有誤差存在。對篩選出的差異基因及富集分析得到的GO term仍需PCR和Western等進一步篩選驗證。
綜上所述,本研究初步證實了人胚胎干細胞H9體外對人肝癌細胞HepG2有一定程度的抑制作用,具體表現為對肝癌細胞的細胞生長、凋亡、侵襲、遷移等的抑制和細胞周期的影響。然而,其具體的機制仍有待進一步研究。
[1] Reya T,Clevers H.Wnt signalling in stem cells and cancer[J].Nature,2005,434(7035):843-850.
[2] Li L,Neaves WB.Normal stem cells and cancer stem cells:the niche matters[J].Cancer Res,2006,66(9):4553-4557.
[3] Pardal R,Clarke MF,Morrison SJ.Applying the principles of stem-cell biology to cancer[J].Nat Rev Cancer,2003,3(12):895-902.
[4] Miller SJ,Lavker RM,Sun TT.Interpreting epithelial cancer biology in the context of stem cells:tumor properties and therapeutic implications[J].Biochim Biophys Acta,2005,1756(1):25-52.
[5] Nakazawa K,Kalassy M,Sahuc F,etal.Pigmented human skin equivalent-as a model of the mechanisms of controI of cell-cell and cell-matrix interactions[J].Med Bio I Eng Comput,1998,36(6):813-820.
[6] Xie J,Haslam SZ.Extracellular matrix regulates ovarian hormone dependent proliferation of mouse mammary epithelial cells[J].Endocrinology,1997,138(6):2466-2473.
[7] Woodward TL,Xie J,Fendrick JL,etal.Proliferation of mouse mammary epithelial cells in vitro:Interactions among epidermal growth factor,insulin-like growth factorⅠ,ovarian hormones,and extracellular matrix proteins[J].Endocrinology,2000,141(10):3578-3586.
[8] Thiery JP,Chopin D.Epithelial cell plasticity in development and tumor progression[J].Cancer Metastasis Rev,1999,18(1):31-42.
[9] Kamolz LP,Kolbus A,Wick N,etal.Cultured human epithelium:human umbilical cord blood stem cells differentiate into keratinocytes under in vitro conditions[J].Burns,2006,32(1):16-19.
[10] Ubels JL,Hoffman HM,Srikanth S,etal.Gene expression in rat lacrimal gland duct cells collected using laser capture microdissection:evidence for K+secretion by duct cells[J].Invest Ophthalmol Vis Sci,2006,47(5):1876-1885.
[11] Seam N,Gonzales DA,Kern SJ,etal.Quality control of serum albumin depletion for proteomic analysis[J].Clin Chem,2007,53(11):1915-1920.
[12] Lee LM,Seftor EA,Bonde G,etal.The fate of human malignant melanoma cells transplanted into zebrafish embryos:assessment of migration and cell division in the absence of tumor formation[J].Dev Dyn,2005,233(4):1560-1570.
[13] Giuffrida D,Rogers IM,Nagy A,etal.Human embryonic stem cells secrete soluble factors that inhibit cancer cell growth[J].Cell Prolif,2009,42(6):788-798.
[14] Liotta LA.Cancer cell invasion and metastasis[J].Sci Am,1992,266(2):54-59,62-53.
[15] Chang BD,Swift ME,Shen M,etal.Molecular determinants of terminal growth arrest induced in tumor cells by a chemoth era-peutic agent[J].Proc Natl Acad Sci U S A,2002,99(1):389-394.
[16] Zhou Y,Gwadry FG,Reinhold WC,etal.Transcriptional regulation of mitotic genes by camptothecin-induced DNA damage:Microarray analysis of dose-and time-dependent effects[J].Cancer Res,2002,62(6):1688-1695.
[17] Yaddanapudi K,Mitchell RA,Putty K,etal. Vaccination with embryonic stem cells protects against lung cancer:is a broad-spectrum prophylactic vaccine against cancer possible? [J]. PLoS One,2012,7:e42289.
[收稿2016-09-23 修回2016-10-19]
(編輯 張曉舟)
InhibitoryeffectofhumanembryonicstemcellsonHepG2cellsinvitro
ZHENGLiang-Dong,HEXue-Mei,ZHANGTing,LIUJie,HUOBen-Nian,LIUMeng-Nan,WANGXue,FENGTao.ResearchCenterofMolecularMedicineandCancer,ChongqingMedicalUniversity,Chongqing400016,China
Objective:To study the inhibitory effect of human embryonic stem cells on the HepG2 cells in vitro.Methods:The co-culture system of Human embryonic stem cells (H9) and liver cancer HepG2 cells was established.The effect of H9 on the biological behavior of HepG2 cells was observed by microscope,the flow cytometry was used to detect the apoptosis of tumor cells and the cell cycle alteration.Transwell assay was used to detect the migration and invasion of tumor cells.Gene microarray technique was used to examine the change of gene expression profile of HepG2 cells.Results:In the process of co-culture,the growth of hepatoma cells was inhibited.With the extension of the culture time,cells decreased gradually,and occurred signs of aging or apoptosis.Flow cytometry test results showed that the apoptosis rate of hepatoma cells was significantly increased,and the cell cycle was blocked in the G0/G1 phase.Transwell test results showed that the invasion and migration of HepG2 cells were decreased.The gene chip results showed that the whole genome expression profile of HepG2 cells had a significant change.Conclusion:The human embryonic stem cells had an inhibitory effect on HepG2 cells in vitro.
Embryonic stem cells;Hepatoma;Co-culture;Inhibition;Apoptosis
10.3969/j.issn.1000-484X.2017.06.012
①本文為國家自然科學基金資助項目(81071770,81201679)。
②同時供職于重慶市生物化學與分子藥理學重點實驗室,重慶400016。
鄭良棟(1989年-),男,碩士,主要從事分子藥理學與腫瘤藥理學研究,E-mail:zishangjun@126.com。
及指導教師:馮 濤(1963年-),男,教授,博士生導師,主要從事腫瘤機理和治療方面的研究,E-mail:478240069@qq.com。
R392
A
1000-484X(2017)06-0864-05
猜你喜歡
天津醫科大學學報(2019年3期)2019-08-13 06:53:08
中成藥(2016年8期)2016-05-17 06:08:14
癌癥進展(2016年12期)2016-03-20 13:16:17
罕少疾病雜志(2016年5期)2016-03-11 16:34:44
吉林大學學報(醫學版)(2015年1期)2015-12-17 07:47:28
腫瘤預防與治療(2015年1期)2015-09-26 07:26:20
中國當代醫藥(2015年16期)2015-03-01 02:03:11
中國醫藥導報(2015年26期)2015-02-28 22:07:59
肝膽胰外科雜志(2015年4期)2015-02-27 11:12:34
肝膽胰外科雜志(2015年4期)2015-02-27 11:12:24
