施氮肥對華北平原土壤氨氧化細菌和古菌數量及群落結構的影響
2017-07-20 13:11:03楊亞東張明才胡君蔚胡躍高曾昭海
生態學報 2017年11期
關鍵詞:研究
楊亞東,張明才,胡君蔚,張 凱,胡躍高,曾昭海
中國農業大學農學院, 北京 100193
?
施氮肥對華北平原土壤氨氧化細菌和古菌數量及群落結構的影響
楊亞東,張明才,胡君蔚,張 凱,胡躍高,曾昭海*
中國農業大學農學院, 北京 100193
利用熒光定量PCR、末端限制性片段長度多樣性(T-RFLP)和基因克隆文庫技術,比較了4種施氮水平(不施氮肥,0 kg N/hm2,CK;施低水平氮肥,75 kg N/hm2,N1;施中水平氮肥,150 kg N/hm2,N2;施高水平氮肥,225 kg N/hm2,N3)下華北平原地區小麥季表層(0—20 cm)土壤總細菌、氨氧化細菌(AOB)和氨氧化古菌(AOA)的豐度和群落結構。結果表明,土壤總細菌、AOB和AOA數量分別在每克干土5.74×109—7.50×109、8.89×106—2.66×107和3.83×108—7.78×108之間。不同施氮量土壤AOA數量均高于AOB數量,AOA/AOB值在81.72—14.38之間。增施氮肥顯著顯著提高AOB數量(P<0.05),對總細菌和AOA數量的影響不顯著(P>0.05)。與CK相比,處理N1、N2和N3中AOB數量分別提高了0.64、1.50和1.99倍。增施氮肥顯著改變了AOB和AOA的群落結構,且不同施氮量處理中AOB群落結構差異更大。系統進化分析顯示,施氮肥小麥土壤AOB主要為Nitrosospira屬類群,分布在Cluster 3的兩個分支中;AOA分布在Cluster S的4個分支中。相關性分析顯示,AOB數量與全氮和銨態氮含量呈顯著正相關關系,與土壤pH和碳氮比呈顯著負相關關系(P<0.05);AOA數量與硝態氮含量和土壤pH呈顯著正相關關系,與銨態氮含量呈顯著負相關關系(P<0.05)。研究結果表明:增施氮肥可顯著改變華北平原地區堿性土壤AOB數量與群落結構,該地區小麥土壤中AOB比AOA對氮肥響應更敏感。
氮肥;氨氧化細菌;氨氧化古菌;豐度;群落結構;末端限制性片段長度多樣性
硝化作用是農田氮素循環的重要環節,也是土壤氮素轉化的關鍵過程[1]。氨氧化作用是硝化作用的第一步反應,也是整個過程的限速步驟[2]。在很長一段時間內,來自變形菌綱的氨氧化細菌(AOB)[2- 3]被認為是其唯一承擔者,直到氨氧化古菌(AOA)[4- 5]的發現,才證實了氨氧化過程是由AOB和AOA兩類微生物共同承擔。Leininger等[6]首次報道了原始土壤和農業土壤中AOA在數量上遠高于AOB,再次證實了AOA是參與土壤環境中氨氧化過程的重要微生物。
前人的研究表明,AOB和AOA數量和群落組成受氨的有效性[3]、溫度[7]、土壤pH[8]、土壤含水量[9]和施肥措施[10]等因素影響。Zhou等[11]研究表明施用氮肥可以提高AOB數量,而施用有機肥則增加了AOA多樣性;張苗苗等[12]研究顯示長期施用氮肥降低了AOB多樣性,顯著提高了AOA數量;Di等[13]發現草地土壤中AOA數量遠高于AOB,而施用動物尿液(富含NH3)顯著促進了AOB數量的增加;Chen等[14]研究結果也證實了施肥對AOA數量影響強于對AOB的影響,且影響AOA群落組成;Long等[15]研究卻發現施氮肥對AOB和AOA數量影響不顯著;關于施氮水平對AOB和AOA的影響研究方面,Zhong等[16]研究了溫室蔬菜不同施氮量對AOB和AOA的影響,結果表明土壤施氮量增加后,AOB和AOA的數量均下降,且增施氮肥對AOB群落結構的影響強于對AOA群落結構的影響。施用氮肥對土壤AOB和AOA的影響因氮肥類型和施氮量水平不同存在顯著差異,而施氮水平對華北地區小麥生產系統土壤AOB和AOA數量和群落組成的影響仍不清楚,亟待研究。
華北平原是我國重要的糧食主產區,長期小麥-玉米集約種植模式需要大量的肥水投入,帶來溫室氣體(N2O、NO)排放和地下水污染等問題[10],給區域造成很大的生態壓力。研究不同施氮水平對土壤AOB和AOA數量和群落結構的影響可以進一步明確氮肥轉化的微生物機制。本研究以連續5a的田間定位試驗為基礎,以華北平原小麥季土壤為研究對象,采用熒光定量PCR(Real-time quantitative PCR,q-PCR)、末端限制性片段長度多樣性(T-RFLP)和基因克隆文庫技術,比較了4種施氮水平下小麥土壤AOB和AOA數量和群落組成的差異。旨在闡明華北平原小麥土壤中AOB和AOA對氮肥的響應機制,為土壤科學施肥、維護土壤微生物多樣性和農業可持續發展等提供依據。
1 材料與方法
1.1 試驗設計和土壤樣品采集
試驗地位于河北省滄州市吳橋縣中國農業大學吳橋實驗站(37°37′N,116°22′E)。該地區屬暖溫帶半濕潤大陸性季風氣候,土壤類型為中壤質潮土,成土母質為河流沖積物。年平均氣溫12.9℃,全年積溫(≥ 0℃)為4,826℃,無霜期201 d,降雨量為562 mm,日照時數2724 h。氮肥梯度試驗起始于2010年,種植模式為冬小麥-夏玉米一年兩熟,冬小麥季施氮量設置4個梯度,分別為不施氮肥(0 kg N/hm2,CK),施低水平氮肥(75 kg N/hm2,N1),施中水平氮肥(150 kg N/hm2,N2)和施高水平氮肥(225 kg N/hm2,N3),磷肥施用量90 kg P2O5/hm2,鉀肥施用量90 kg K2O/hm2。氮肥、磷肥和鉀肥分別為尿素、磷酸二氫鈣和硫酸鉀,肥料均基施。小區面積為24 m2,重復3次,完全隨機區組排列。
土壤樣品采集于2015年6月冬小麥收獲后,采用對角線五點采樣法,每個小區采集5個點0—20 cm土壤。同一處理的土壤樣品混合成一個樣品,除去碎石、根系及其他雜物,過2 mm篩,將土樣分成兩部分,一部分迅速用錫箔紙包裹后液氮冷凍,用冰盒保存帶回實驗室后-80℃保存,用于DNA提取及后續分析;另一部分裝入自封袋后帶回實驗室,用鮮土樣測定銨態氮和硝態氮含量,剩余土壤自然風干后測定土壤pH、有機質和全氮含量。
1.2 土壤理化性質測定
土壤pH采用電位法(水∶土=2.5∶1)測定,土壤全氮采用濃硫酸-蒸餾滴定法測定,土壤銨態氮和硝態氮用2 mol/L KCl溶液浸提新鮮土樣后,采用流動分析儀測定(SAN++, Skalar, Holland),土壤有機質采用重鉻酸鉀容量法測定[17]。
1.3 土壤總DNA提取
取0.2—0.5 g冷凍土壤樣品,用E.Z.N.A.? Soil DNA Kit(Omega,USA)試劑盒提取土壤總DNA。用1%瓊脂糖凝膠電泳檢測總DNA完整性,用NANO Quant(Tecan,Switzerland)測定提取DNA濃度和純度。
1.4 總細菌16S rDNA、AOB和AOAamoA基因的擴增與熒光定量PCR分析
分別用引物BACT1369F/PROK1541R、amoA- 1F/amoA- 2R和Arch-amoAF/Arch-amoAR擴增細菌16S rDNA、AOB和AOAamoA基因(表1)。PCR體系為50 μL,分別含10× Buffer 5.0 μL,dNTP(各2.5 μmol/L)4.0 μL,上下游引物(10 μM)各1.0 μL,DNA 模板(1—10 ng)1 μL,rTaq(5 U/μL)1.0 μL,最后用ddH2O補至50.0 μL。PCR程序:95℃預變性3 min;95℃變性30 s,55℃退火30 s,72℃延伸45 s,35個循環;最后72℃延伸5 min。回收PCR產物連接至pMD18-T載體,轉化至大腸桿菌DH5α感受態中,經Amp+、IPTG和X-gal的LB平板篩選陽性克隆,測序分析。
標準曲線的制作:提取測序正確的細菌16S rDNA、AOB和AOAamoA基因的陽性克隆質粒,用NANO Quant(Tecan,Switzerland)測定質粒濃度,按10倍梯度稀釋質粒至103—108拷貝數/微升,-80℃保存,用于制備標準曲線。
熒光定量PCR在ABI 7300(ABI,USA)上進行,反應體系為20 μL,分別含SYBR?PremixExTaqⅡ(Thi RNaseH Plus)10.0 μL,上下游引物各0.8 μL(10 μM),ROX Reference Dye(50x)0.4 μL,DNA 模板2.0 μL(1—10 ng),用ddH2O補至20.0 μL。PCR引物及程序如表1所示。

表1 熒光定量PCR引物及反應條件
Y表示C或T;R表示A或G;K表示G或T;S表示C或G;AOB:氨氧化細菌Ammonia oxidizing bacteria;AOA: 氨氧化古菌Ammonia oxidizing archaea
1.5 末端限制性片段長度多樣性分析
對AOB和AOA進行T-RFLP分析,選用amoA- 1F/amoA- 2R和Arch-amoAF/Arch-amoAR擴增AOB和AOAamoA基因,其中引物amoA- 1F和Arch-amoAF的5′端用6-FAM進行熒光標記,PCR體系和程序同1.4所示。回收PCR產物后,用限制性內切酶RsaⅠ(Taraka,大連)分別酶切AOB和AOAamoA基因。反應體系為20 μL,分別含10× T Buffer 2 μL,0.1% BSA 2 μL,RsaⅠ(10 U/μL)1 μL,PCR產物DNA 10 μL,最后用ddH2O補至20.0 μL。37℃水浴酶切3 h,重復3次,用乙醇沉淀法純化酶切產物,最后將3個重復混勻成一個樣品。取回收的酶切產物0.3 μL、分子量內標(GS500LIZ)0.5 μL和甲酰胺9.5 μL混合均勻后加入96孔板中,95℃變性5 min,4℃冷卻后離心,1× Buffer緩沖液上機檢測,采用ABI 3730(ABI,USA)測序儀測定分析。用GeneMarker V 2.2(ABI,USA)分析電泳圖中各峰的片段長度和峰面積,忽略長度小于50 bp和峰面積小于1%的峰。將峰的信號強度標準化,每種末端限制性片段(T-RF)類型的相對豐度用百分比形式表示。
1.6 克隆和測序
將4個土壤樣品的DNA混合后做模板,采用amoA- 1F/amoA- 2R和Arch-amoAF/Arch-amoAR 擴增AOB和AOAamoA基因,體系和擴增條件同1.4所示,引物不添加熒光標記。擴增產物回收后,分別連接至pMD18-T載體(Taraka,大連)上,轉化至大腸桿菌DH5α感受態中,經篩選后分別選取了53和56個陽性克隆進行測序分析。測序結果經分析比對后,將同源性≥97%的序列歸為一類,選一條用于構建系統發育樹。將具有代表性的AOB和AOAamoA基因序列提交至NCBI數據庫中,獲AOB和AOA登錄號分別為KU365731—KU365744和KU365745—KU365753。
1.7 數據分析
數據的方差分析和相關性分析用SPSS 20.0軟件(SPSS 20.0 for Windows,SPSS Inc,USA)完成;土壤總細菌、AOB和AOA數量的數據取對數后,用SigmaPlot 12.5軟件制圖;測序核苷酸序列用DNAMAN 6.0軟件分析,并用MEGA 5.0軟件構建系統進化樹,選用鄰-接法(Neighbour-Joining);用末端限制性片段(T-RF)的長度和相對含量數據進行主成分分析,由Canoco 5.0軟件完成。
2 結果
2.1 土壤基礎理化性質
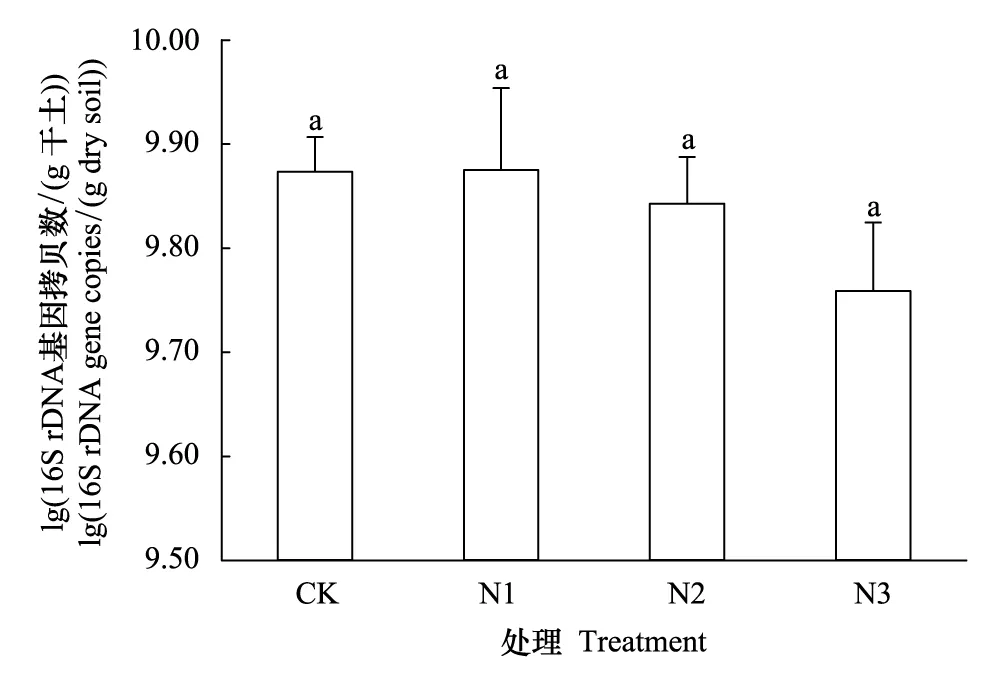
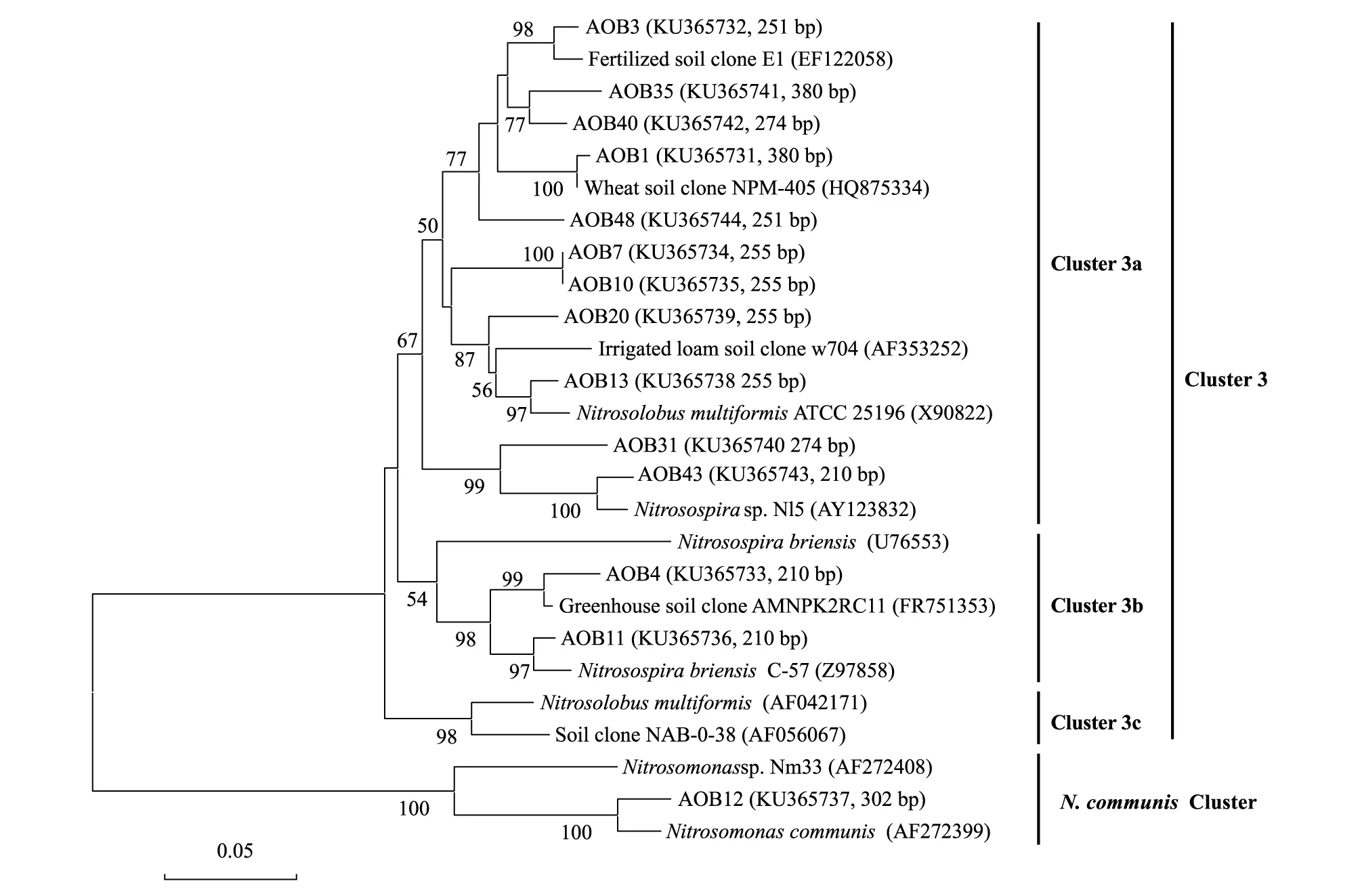
不同施氮量0—20 cm土層土壤中除有機質含量差異不顯著(14.96—15.49 g/kg,P>0.05)外,其他理化性質存在顯著差異(表2)。隨施氮量增加,土壤pH呈顯著下降趨勢(8.46—8.29),依次為N3 表2 不同施氮處理土壤基礎理化性質 不同字母表示處理間差異顯著(P<0.05),數據為平均值±標準差,n=3;CK 0 kg N/hm2,N1 75 kg N/hm2,N2 150 kg N/hm2,N3 225 kg N/hm2 2.2 土壤總細菌、AOB和AOA的數量 以16S rDNA和amoA基因為靶標,用熒光定量PCR測定土壤總細菌、AOB和AOA數量。結果顯示,不同施氮處理土壤總細菌數量在5.74×109—7.50×109拷貝數/g干土,處理間差異不顯著(P>0.05,圖1)。不同施氮處理土壤AOB數量在8.89×106—2.66×107拷貝數/g干土,不同施氮處理AOB數量均差異顯著,隨施氮量增加,AOB數量顯著增加(P<0.05)。與CK相比,N1、N2、N3中AOB數量分別提高了64%、150%和199%。AOA數量在3.83×108—7.78×108拷貝數/g干土,CK、N1和N2中AOA數量顯著高于N3(P<0.05,圖2)。相關性分析結果顯示,總細菌數量僅與硝態氮(r=-0.582,n=12,P<0.05)呈顯著負相關關系。AOB數量與全氮(r=0.838,n=12,P<0.01)和銨態氮(r=0.597,n=12,P<0.05)呈顯著正相關關系,與土壤pH(r=-0.859,n=12,P<0.01)和碳氮比(r=-0.784,n=12,P<0.01)呈顯著負相關關系。AOA數量與硝態氮(r=0.926,n=12,P<0.01)和土壤pH(r=0.594,n=12,P<0.05)呈顯著正相關關系,與銨態氮(r=-0.717,n=12,P<0.05)呈顯著負相關關系。 不同施氮處理土壤AOA數量均顯著高于AOB數量,AOA與AOB數量比值在81.72—14.38之間,隨施氮量增加,AOA與AOB數量比值呈下降趨勢。熒光定量結果表明,增施氮肥能顯著提高小麥土壤AOB數量,對總細菌和AOA數量影響不顯著。此外,總細菌、AOB和AOA數量與土壤各環境因子的相關性存在明顯差異。 圖1 不同施氮處理細菌16S rDNA基因拷貝數 Fig.1 Abundance of bacteria 16S rDNA gene under different N fertilizer application rate treatments 圖2 不同施氮處理氨氧化細菌和氨氧化古菌amoA基因拷貝數 Fig.2 Abundance of ammonia oxidizing bacteria and archeae amoA genes under different N fertilizer application rate treatments 2.3 AOB和AOA的群落結構 選用限制性內切酶RsaⅠ分別對AOB和AOA進行T-RFLP分析。4個處理共得到10個AOB的T-RF片段長度(圖3a),其中96、210、251、244、255、270、274和380 bp在4個處理中均存在,63 bp只存在于CK、N1和N2中,248 bp只存在于N2和N3中。長度為210、251、255、274和380 bp的T-RFs為主要片段,分別占總片段的3.91%—11.38%、26.60%—29.71%、26.97%—39.49%、6.50%—11.20% 和11.87%—17.76%,合計占總片段的92.74%—93.34%。這些片段所代表的AOB在土壤中占優勢,其中251、255和380 bp長度所占比例均超過10%,為占絕對優勢的3個AOB類群。 4個處理共得到11個AOA的T-RF片段長度(圖3b),其中60、98、196、259、261、263和299 bp在4個處理中均存在,63和65 bp只存在于CK、N1和N2中,256 bp只存在于N1、N2和N3中,261 bp只存在于CK中,265 bp只存在于N1和N2中。長度為60、196、256、259和299 bp的T-RFs為主要片段,分別占總片段的53.26%—64.76%、7.79%—12.88%、12.26%—16.17%、2.94%—10.45% 和2.46%—5.34%,合計占總片段的85.13%—94.91%。這些片段所代表的AOA在土壤中占優勢,其中60和256 bp長度所占比例均超過10%,為占絕對優勢的2個AOA類群。 2.4 AOB和AOAamoA基因序列的系統發育樹構建 經測序比對分析后,分別得到14和9個AOB和AOAamoA基因操作分類單元(operational taxonomic unit,OTU)。AOBamoA基因序列兩兩同源性在69.92%—96.54%之間,14條序列的同源性為91.77%;AOAamoA基因序列兩兩同源性在77.48%—96.22%之間,9條序列的同源性為87.44%。系統進化分析結果顯示:AOBamoA基因序列聚類在兩個Cluster中,其中,AOB 12聚類在N.communisCluster中,其余AOB都聚類在Cluster 3中。聚類在Cluster 3中的AOB分布在Cluster 3a和Cluster 3b兩個分支中,除片段長度為210 bp的T-RF聚類在Cluster 3b外,其他片段長度T-FRs均聚類于Cluster 3a(圖4);所有AOAamoA基因序列都聚類在Cluster S中,分布在4個分支中。其中,196 bp T-RF聚類在分支Ⅰ和Ⅲ中,60 bp T-RF聚類在分支Ⅱ中,63 bp T-RF聚類在分支Ⅱ和Ⅲ中,299 bp T-RF聚類在分支Ⅰ和Ⅳ中(圖5)。 圖4 氨氧化細菌amoA基因系統發育樹Fig.4 Phylogenetic tree of ammonia oxidizing bacteria amoA gene sequences 圖5 氨氧化古菌amoA基因系統發育樹分析Fig.5 Phylogenetic tree of ammonia oxidizing archaea amoA gene sequences 2.5 主成分分析 基于T-RFs的主成分分析(PCA)結果顯示,第一、二排序軸是主成分軸。對AOB群落變異的解釋量達到95.16%,其中軸1和軸2分別為76.81%和18.35%(圖6a);對AOA群落變異的解釋量達到94.44%,其中軸1和軸2分別為77.22%和17.22%(圖6b)。處理CK、N1、N2和N3的AOB都明顯分開,分布在4個象限中;而4個處理的AOA群落結構差異弱于AOB,其中,N1和N2聚在一起(第三象限),與CK和N3明顯分開。PCA結果表明:施氮量顯著影響小麥土壤AOB和AOA的群落結構,且不同施氮處理的AOB群落結構差異強于AOA。 圖6 不同施氮處理氨氧化細菌(a)和古菌(b)amoA基因T-RFs主成分分析Fig.6 Principal component analysis (PCA) of T-RFs of ammonia oxidizing bacteria (a) and archaea (b) amoA genes under different N fertilizer application rate treatments 3.1 施氮肥對小麥土壤總細菌、AOB和AOA數量的影響 土壤總細菌數量是反映土壤總微生物活性的一個重要參數。本研究中土壤總細菌16S rDNA基因在5.74×109—7.50×109拷貝數/g干土之間,與Shen等[17]研究發現細菌數量在3.39×109—1.23×1010拷貝數/g干土之間的結果相似,相較于He等[8]在水稻土壤中細菌數量1010拷貝數/g干土數量級低,且總細菌數量僅與硝態氮呈顯著負相關關系。小麥與水稻土壤類型和理化性質差異可能是引起總細菌數量差異的主要原因。此外,本研究不同施氮水平土壤總細菌數量差異不顯著,與He等[8]和Shen等[17]研究結果一致,表明土壤總細菌數量對施氮肥響應不明顯。 AOB和AOA是土壤氮循環的關鍵微生物,直接影響著土壤健康、肥力狀況和生產力等。本研究不同施氮量處理AOA數量均高于AOB,AOA與AOB數量比值在81.72—14.38之間,與Leininger等[6]、He等[8]、Shen等[17]和Jia等[18]研究結果相符,進一步證實了堿性土壤更適合AOB。AOB數量在8.89×106—2.66×107拷貝數/g干土,與He等[8]、Chen等[14]和Wessén等[19]研究中AOB數量級一致,比Shen等[16]、Wang等[20]和Yao等[21]研究中AOB高出2—3個數量級。AOA數量在3.83×108—7.78×108拷貝數/g干土,與Chen等[14]研究中水稻土壤AOA數量級一致,比He等[8]、Shen等[17]和Wang等[20]研究中AOA數量高出1—2個數量級。此外,本研究中AOB數量隨施氮量增加而顯著增加,而AOA數量差異不顯著,與Di等[22]研究發現高氮草地土壤中AOB數量隨施氮量增加而增加一致,而與Zhong 等[16]研究表明隨著施氮量增加,AOB和AOA數量均下降不同。 He等[10]研究表明土壤pH是造成AOB和AOA數量差異的主要原因,而Chen等[14]認為肥料是引起AOA數量變化的主要原因,土壤pH和肥料均對AOB數量產生影響;Di等[22]認為硝態氮含量是引起AOB數量增加的原因;但Zhong等[22]在溫室條件下卻得到相反的結果,認為硝態氮含量是導致AOB數量下降的原因,而AOA數量下降則可能是由于土壤pH降低造成。本研究中AOB數量與土壤pH呈極顯著負相關關系,與全氮含量呈極顯著正相關關系,AOB數量上升可能是土壤pH下降和肥料作為底物導致,與Di等[22]結果一致。AOA數量與土壤pH呈顯著正相關關系,而與全氮含量相關性不顯著,AOA數量的降低,可能是由施氮量增加引起的土壤pH下降造成,而非氮肥作為底物造成,與He等[8]和Chen等[14]研究結果相符。不同施氮處理土壤pH均呈堿性,且長期增施氮肥引起土壤pH降低導致了AOA數量下降和AOB數量上升,與Shen等[23]報道的AOB適合在中性至堿性、氮素含量豐富的土壤中生存,而AOA適合在酸性土壤中生存的結果一致。本研究中不同施氮處理AOA在數量上占優勢,隨著施氮量增加,AOB數量顯著上升,AOA數量上的優勢減弱,表明AOB對氮肥更敏感,高氮條件下土壤中氨氧化過程的主要承擔者可能是AOB而非AOA,與Jia等[18]和Shen等[23]結論一致。 3.2 施氮肥對小麥土壤AOB和AOA群落組成的影響 本研究用限制性內切酶RsaⅠ酶切AOB和AOAamoA基因經后測序分別得到了10和11個T-RFs,與Zhou等[11]研究紫土中AOB用AccⅡ酶切得到11個T-RFs,莫旭華等[24]研究小麥土壤中AOA用AfaⅠ酶切得到10個T-RFs的結果一致;但Chen等[25]研究發現草原土壤中AOB用MobⅠ酶切后僅得到4個T-RFs,Chen等[14]研究發現水稻土壤中AOA用MobⅠ酶切后僅得到4個T-RFs。T-RFs數量不同,可能是選用內切酶不同導致,也可能是由于土壤類型差異引起。 不同施氮處理中,T-RF長度為63、96、274、380、248和255 bp的AOB類群所占比例差異較大,且隨施氮量的增加表現出不同的變化趨勢;而T-RF長度為256和265 bp的AOA類群差異較大,且4個處理中AOA均含有2個絕對優勢的類群(>10%),表明小麥土壤中AOB的群落組成受施氮量的影響較大,而AOA具有較穩定性的群落結構,與Shen等[17]堿性土壤條件下得到的結果一致,而與He等[8]、Chen等[14]、Ying等[26]酸性土壤條件下得到的結果相反。另外,PCA分析也顯示4個處理AOB比AOA群落結構差異性更大,AOB群落結構更易受氮肥的影響。這些結果都證實了施肥引起AOB和AOA群落結構的變化,且堿性條件下AOB比AOA對氮肥施用量更敏感。 3.3 施氮肥小麥土壤中AOB和AOA的系統發育 本研究中絕大多數AOB都聚類在Cluster 3的兩個分支中,除AOB 12屬于Nitrosomonas屬外,其他AOB都聚類在Nitrosospira屬中,與莫旭華等[24]和Chu等[27]在小麥土壤中得到的AOB都屬于Nitrosospira屬的結果相符;也與水稻土壤[14]、黃土旱塬黑壚土壤[28]和草原[25]等土壤中AOB都主要聚類在Nitrosospira屬中的結果一致;符合Kowalchuk等[29]報道的土壤環境中氨氧化細菌以Nitrosospira屬為主,而非Nitrosomonas屬的結論。He等[8]、Chen等[14]和Shen等[16]在水稻土壤中均發現了聚類于Cluster S和Cluster M的AOA類群,pH越低,聚類于Cluster M的AOA類群越多,并且Cluster S在農田和草地生態系統中占絕對優勢。本研究中所有AOA均聚類于Cluster S中,未發現聚類在Cluster M的AOA類群,與Chen等[25]和Shen等[30]的結果一致,與農田生態系統中聚類于Cluster S的AOA占絕對優勢的結果相符,本研究中土壤pH呈堿性是導致所有AOA都分布在Cluster S中的主要原因。 本研究對長期施用不同氮肥水平下小麥季土壤氨氧化微生物的群落結構和豐度特征進行了研究,結果發現,隨施氮量增加引起土壤pH下降及其他理化性質的改變,施氮量增加顯著提高AOB的數量,對AOB群落結構的影響強于對AOA群落結構的影響,表明該地區小麥土壤中AOB比AOA對施用氮肥的響應更敏感。 致謝:本研究得到農業部都市農業(北方)重點實驗室馬蘭青教授、楊明峰副教授的幫助,特此致謝。 [1] 賀紀正, 張麗梅. 氨氧化微生物生態學與氮循環研究進展. 生態學報, 2009, 29(1): 406- 415. [2] Prosser J I. Autotrophic nitrification in bacteria. Advances in Microbial Physiology, 1989, 30: 125- 181. [3] Kowalchuk G A, Stephen J R. Ammonia-oxidizing bacteria: a model for molecular microbial ecology. Annual Review of Microbiology, 2001, 55(1): 485- 529. [4] Venter J C, Remington K, Heidelberg J F, Halpern A L, Rusch D, Eisen J A, Wu D Y, Paulsen I, Nelson K E, Nelson W, Fouts D E, Levy S, Knap A H, Lomas M W, Nealson K, White O, Peterson J, Hoffman J, Parsons R, Baden-Tillson H, Pfannkoch C, Rogers Y H, Smith H O. Environmental genome shotgun sequencing of the Sargasso Sea. Science, 2004, 304(5667): 66- 74. [5] K?nneke M, Bernhard A E, De La Torre J R, Walker C B, Waterbury J B, Stahl D A. Isolation of an autotrophic ammonia-oxidizing marine archaeon. Nature, 2005, 437(7058): 543- 546. [6] Leininger S, Urich T, Schloter M, Schwark L, Qi J, Nicol G W, Prosser J I, Schuster S C, Schleper C. Archaea predominate among ammonia-oxidizing prokaryotes in soils. Nature, 2006, 442(7104): 806- 809. [7] Tourna M, Freitag T E, Nicol G W, Prosser J I. Growth, activity and temperature responses of ammonia-oxidizing archaea and bacteria in soil microcosms. Environmental Microbiology, 2008, 10(5): 1357- 1364. [8] He J Z, Shen J P, Zhang L M, Zhu Y G, Zheng Y M, Xu M G, Di H J. Quantitative analyses of the abundance and composition of ammonia-oxidizing bacteria and ammonia-oxidizing archaea of a Chinese upland red soil under long-term fertilization practices. Environmental Microbiology, 2007, 9(9): 2364- 2374. [9] Hastings R C, Butler C, Singleton I, Saunders J R, McCarthy A J. Analysis of ammonia-oxidizing bacteria populations in acid forest soil during conditions of moisture limitation. Letters in Applied Microbiology, 2000, 30(1): 14- 18. [10] Strauss S L, Reardon C L, Mazzola M. The response of ammonia-oxidizer activity and community structure to fertilizer amendment of orchard soils. Soil Biology and Biochemistry, 2014, 68: 410- 418. [11] Zhou Z F, Shi X J, Zheng Y, Qin Z X, Xie D T, Li Z L, Guo T. Abundance and community structure of ammonia-oxidizing bacteria and archaea in purple soil under long-term fertilization. European Journal of Soil Biology, 2014, 60: 24- 33. [12] 張苗苗, 王伯仁, 李冬初, 賀紀正, 張麗梅. 長期施加氮肥及氧化鈣調節對酸性土壤硝化作用及氨氧化微生物的影響. 生態學報, 2015, 35(19): 6362- 6370. [13] Di H J, Cameron K C, Shen J P, Winefield C S, O′Callaghan M, Bowatte S, He J Z. Ammonia-oxidizing bacteria and archaea grow under contrasting soil nitrogen conditions. FEMS Microbiology Ecology, 2010, 72(3): 386- 394. [14] Chen X, Zhang L M, Shen J P, Wei W X, He J Z. Abundance and community structure of ammonia-oxidizing archaea and bacteria in an acid paddy soil. Biology and Fertility of Soils, 2011, 47(3): 323- 331. [15] Long X E, Chen C R, Xu Z H, Linder S, He J Z. Abundance and community structure of ammonia oxidizing bacteria and archaea in a Sweden boreal forest soil under 19-year fertilization and 12-year warming. Journal of Soils and Sediments, 2012, 12(7): 1124- 1133. [16] Zhong W H, Bian B Y, Gao N, Min J, Shi W M, Lin X G, Shen W S. Nitrogen fertilization induced changes in ammonia oxidation are attributable mostly to bacteria rather than archaea in greenhouse-based high N input vegetable soil. Soil Biology and Biochemistry, 2016, 93: 150- 159. [17] Shen J P, Zhang L M, Zhu Y G, Zhang J B, He J Z. Abundance and composition of ammonia-oxidizing bacteria and ammonia-oxidizing archaea communities of an alkaline sandy loam. Environmental Microbiology, 2008, 10(6): 1601- 1611. [18] Jia Z J, Conrad R. Bacteria rather than archaea dominate microbial ammonia oxidation in an agricultural soil. Environmental Microbiology, 2009, 11(7): 1658- 1671. [19] Wessén E, Nyberg K, Jansson J K, Hallin S. Responses of bacterial and archaeal ammonia oxidizers to soil organic and fertilizer amendments under long-term management. Applied Soil Ecology, 2010, 45(3): 193- 200. [20] Wang Y, Zhu G B, Song L Y, Wang S Y, Yin C Q. Manure fertilization alters the population of ammonia-oxidizing bacteria rather than ammonia-oxidizing archaea in a paddy soil. Journal of Basic Microbiology, 2014, 54(3): 190- 197. [21] Yao H Y, Gao Y M, Nicol G W, Campbell C D, Prosser J I, Zhang L M, Han W Y, Singh B K. Links between ammonia oxidizer community structure, abundance, and nitrification potential in acidic soils. Applied and Environmental Microbiology, 2011, 77(13): 4618- 4625. [22] Di H J, Cameron K C, Shen J P, Winefield C S, O′Callaghan M, Bowatte S, He J Z. Nitrification driven by bacteria and not archaea in nitrogen-rich grassland soils. Nature Geoscience, 2009, 2(9): 621- 624. [23] Shen J P, Zhang L M, Di H J, He J Z. A review of ammonia-oxidizing bacteria and archaea in Chinese soils. Frontiers in Microbiology, 2012, 3: 296- 296, dio: 10.3389/fmicb.2012.00296. [24] 莫旭華, 史榮久, 李慧, 鄭佳, 王元芬, 徐慧. 華北典型旱地小麥土壤amoA基因的PCR-RFLP分析. 生態科學, 2009, 28(1): 49- 55. [25] Chen Y L, Xu Z W, Hu H W, Hu Y J, Hao Z P, Jiang Y, Chen B D. Responses of ammonia-oxidizing bacteria and archaea to nitrogen fertilization and precipitation increment in a typical temperate steppe in Inner Mongolia. Applied Soil Ecology, 2013, 68: 36- 45. [26] Ying J Y, Zhang L M, He J Z. Putative ammonia-oxidizing bacteria and archaea in an acidic red soil with different land utilization patterns. Environmental Microbiology Reports, 2010, 2(2): 304- 312. [27] Chu H Y, Fujii T, Morimoto S, Lin X G, Yagi K, Hu J L, Zhang J B. Community structure of ammonia-oxidizing bacteria under long-term application of mineral fertilizer and organic manure in a sandy loam soil. Applied and Environmental Microbiology, 2007, 73(2): 485- 491. [28] 劉桂婷, 程林, 王保莉, 趙其國, 曲東. 長期不同施肥對黃土旱塬黑壚土氨氧化細菌多樣性的影響. 中國農業科學, 2010, 43(13): 2706- 2714. [29] Kowalchuk G A, Stephen J R, De Boer W, Prosser J I, Embley T M, Woldendorp J W. Analysis of ammonia-oxidizing bacteria of the beta subdivision of the classProteobacteriain coastal sand dunes by denaturing gradient gel electrophoresis and sequencing of PCR-amplified 16S ribosomal DNA fragments. Applied and Environmental Microbiology, 1997, 63(4): 1489 - 1497. [30] Shen X Y, Zhang L M, Shen J P, Li L H, Yuan C L, He J Z. Nitrogen loading levels affect abundance and composition of soil ammonia oxidizing prokaryotes in semiarid temperate grassland. Journal of Soils and Sediments, 2011, 11(7): 1243- 1252. Effects of nitrogen fertilizer application on abundance and community structure of ammonia oxidizing bacteria and archaea in a North China agricultural soil YANG Yadong, ZHANG Mingcai, HU Junwei, ZHANG Kai, HU Yuegao, ZENG Zhaohai* CollegeofAgronomyandBiotechnology,ChinaAgriculturalUniversity,Beijing100193,China The abundance and community structure of total bacteria ammonia oxidizing bacteria, and archaea were investigated by real-time PCR, T-RFLP, and cloning library approaches in a wheat field under different N fertilization rates (no N fertilization, 0 kg N/hm2, CK; low N fertilization rate, 75 kg N/hm2, N1; moderate N fertilization rate, 150 kg N/hm2, N2; high N fertilization rate, 225 kg N/hm2, N3) in North China. The population sizes of total bacteria, AOB and AOA were 5.74×109—7.50×109, 8.89×106—2.66×107and 3.83×108—7.78×108copies/g dry soil, respectively. The population numbers of AOA were higher than that of AOB in all treatments, with AOA to AOB ratios ranged from 81.72 to 14.38. Significant higher numbers for AOB were detected for the high level of N fertilization soil (P<0.05), but not in the population of total bacteria and AOA. The population numbers of AOB observed in N1, N2 and N3 were 1.64, 2.50 and 2.99 times greater than that in the CK treatment, respectively. The N fertilization rates significantly changed the communities of both AOB and AOA, and more variation was observed in the community of AOB. Phylogenetic results showed that the majority AOB sequences fell into two branches of Cluster 3, affiliated withNitrosospiraspecies, and all AOA sequences fell within four branches in Cluster S. Significant positive correlations were observed among the population sizes of AOB to total nitrogen and ammonium, and the population sizes of AOA to soil pH and nitrate (P<0.05). Significant negative correlations were observed among the population sizes of AOB to soil pH and C/N ratio, and the population sizes of AOA to ammonium (P<0.05). These results demonstrated that AOB were more sensitive than AOA to N fertilization in alkaline wheat fields in North China. nitrogen fertilizer; ammonia oxidizing bacteria (AOB); ammonia oxidizing archaea (AOA); abundance; community structure; terminal restriction fragment length polymorphism (T-RFLP) 公益性行業(農業)科研專項(201503121- 11); 河北省科技攻關項目(14227008D) 2016- 03- 22; 網絡出版日期:2017- 02- 22 10.5846/stxb201603220512 *通訊作者Corresponding author.E-mail: zengzhaohai@cau.edu.cn 楊亞東,張明才,胡君蔚,張凱,胡躍高,曾昭海.施氮肥對華北平原土壤氨氧化細菌和古菌數量及群落結構的影響.生態學報,2017,37(11):3636- 3646. Yang Y D, Zhang M C, Hu J W, Zhang K, Hu Y G, Zeng Z H.Effects of nitrogen fertilizer application on abundance and community structure of ammonia oxidizing bacteria and archaea in a North China agricultural soil.Acta Ecologica Sinica,2017,37(11):3636- 3646.



3 討論
4 結論
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
