海島棉GbWRKY40基因的克隆及特征分析
2017-07-31 17:48:08倪志勇加得拉吐留汗邱迎風曲延英陳全家
棉花學報 2017年4期
倪志勇,加得拉·吐留汗,邱迎風,曲延英,陳全家
(新疆農業大學農學院/新疆農業大學農業生物技術重點實驗室,烏魯木齊830052)
海島棉GbWRKY40基因的克隆及特征分析
倪志勇,加得拉·吐留汗,邱迎風,曲延英,陳全家*
(新疆農業大學農學院/新疆農業大學農業生物技術重點實驗室,烏魯木齊830052)
【目的】WRKY轉錄因子調控多種生物學進程,包括植物生長發育和應答多種環境脅迫。本研究旨在分析WRKY轉錄因子在海島棉纖維發育中的功能。【方法】從海島棉中克隆了1個WRKY轉錄因子基因Gb-WRKY40,進行同源性分析、多序列比對,利用熒光定量聚合酶鏈式反應分析其表達模式,通過構建酵母表達載體并轉化酵母菌株AH109研究其轉錄激活活性。【結果】該基因cDNA全長1713 bp,5'非編碼區長261 bp,3'非編碼區長510 bp;開放閱讀框長942 bp,編碼313個氨基酸,預測相對分子質量約為34.138×103,等電點為8.46,包含5個外顯子和4個內含子;其編碼蛋白含有1個WRKY保守區 (WRKYGQK)和1個鋅指基序(C-X5-C-X23-H-X1-H),屬于WRKY家族第Ⅱ類a組,包含3個核定位信號區,與陸地棉GhWRKY40同源性最高。GbWRKY40在根和開花后25 d纖維中表達量高,而且不具有轉錄激活活性。【結論】GbWRKY40可能參與調控棉纖維次生壁發育。
海島棉;GbWRKY40;表達模式;基因克隆;轉錄激活
植物WRKY轉錄因子形成1個大的轉錄因子家族,在植物應答逆境脅迫、表皮毛發育和次生代謝中具有重要的功能[1]。WRKY蛋白包含1個或2個DNA結合結構域,每個結構域由60個氨基酸組成,其N端存在1個高度保守的WRKYGQK序列,C端具有1個鋅指類似基序。根據含有的WRKY結構域數量和鋅指類似基序的特點,WRKY轉錄因子可分為3類:多數蛋白具有1個WRKY結構域,屬于第Ⅱ類,具有2個WRKY結構域的蛋白屬于第Ⅰ類,通常這2類WRKY蛋白具有相同的鋅指類似基序C-X4-5-C-X22-23-H-X1-H;第Ⅲ類WRKY蛋白成員具有1個WRKY結構域,其鋅指類似基序為C-X7-C-X23-H-X-C[1]。
隨著不同棉種基因組測序的完成[2-7],不同棉種WRKY轉錄因子的鑒定成為研究熱點,雷蒙德氏棉 (Gossypium raimondii)、亞洲棉 (G.arboreum)和陸地棉(G.hirsutum)中分別發現116個、109個和 102個WRKY基因[8-10]。陸地棉WRKY轉錄因子的研究較多,許多陸地棉WRKY基因參與調控植物對生物和非生物脅迫的應答。一些WRKY轉錄因子是正調控子,例如在煙草中過表達GhWRKY41基因,能夠提高轉基因植株對干旱和鹽脅迫的忍耐能力[11]。相比野生型植株,過表達GhWRKY34的擬南芥耐鹽能力增強[12]。過表達GhWRKY39-1的轉基因煙草對病原菌侵染和鹽脅迫的忍耐力增強[13]。相反,一些WRKY轉錄因子起負調控子作用。例如,相比野生型煙草,過表達GhWRKY27a的煙草對干旱和立枯絲核菌的抵抗能力降低[14]。GhWRKY68[15]和GhWRKY17[16]降低轉基因煙草對鹽和干旱的忍耐能力。相比野生型煙草,過表達GhWRKY15的轉基因煙草對病毒和真菌侵染敏感[17]。此外,有些WRKY轉錄因子在不同的脅迫應答中功能不同:例如在煙草中過表達GhWRKY25會降低植物對干旱脅迫的忍耐能力,但增強了其耐鹽能力,轉基因植株對葡萄孢菌敏感[18];過表達Gh-WRKY40增強轉基因煙草的傷害耐受力和對青枯病的敏感性[19]。陸地棉WRKY轉錄因子也參與調控植物衰老過程,例如GhWRKY11延緩轉基因擬南芥衰老[20]。相比陸地棉,其他棉種WRKY基因報道較少。最近,Fan等利用轉錄組分析方法從耐鹽的棉花野生種旱地棉(G.aridum)中鑒定了109個WRKY基因,過表達GarWRKY17和GarWRKY104基因的轉基因擬南芥耐鹽性增強[21]。亞洲棉GaWRKY1參與調控倍半萜烯代謝[22]。海島棉(G.barbadenceL.)GbWRKY1是磷缺乏應答的正調控子[23]。
關于海島棉WRKY轉錄因子研究的報道不多,WRKY轉錄因子在海島棉纖維發育中的作用仍不清楚。本研究從海島棉中克隆到1個棉花WRKY類轉錄因子基因GbWRKY40,分析了該基因的序列、轉錄激活活性及表達模式。
1 材料與方法
1.1 植物材料
以海島棉新海21(Xinhai 21)為供試品種,取盛花期的苞葉、花、根、莖、葉及不同發育階段的棉纖維,液氮速凍,-80℃保存備用。
1.2 RNA的提取和cDNA第1鏈的合成
按RNA plant plus reagent試劑盒(Tiangen)說明書操作步驟,提取苞葉、花、根、莖、葉組織總RNA。按多糖多酚植物總RNA提取試劑盒(Tiangen)說明書操作步驟,提取不同發育階段的棉纖維總RNA。使用DNaseⅠ(Thermo)除去總RNA中的基因組DNA。利用First Strand cDNA Synthesis試劑盒(Thermo)將各組織總RNA反轉錄合成cDNA第1鏈。
1.3GbWRKY40基因的克隆及序列分析
根據本課題組前期海島棉胚珠轉錄組和纖維不同發育表達譜數據,共獲得131個WRKY unigene序列,選取1個unigene序列CL2113.Contig3_All設計 1對引物,GbW40-F:5'-ATGGAATCGACTTGGGTGGATACGA-3'和GbW40-R: 5'-TTACCACTTGTGATCTAGAACTTTTCCAGAA-3'。以合成的棉花葉片cDNA第1鏈為模板,用TransStart Taq DNA酶擴增 (TransGen)獲得GbWRKY40的cDNA序列。回收純化PCR(Polymerase chain reaction) 產 物 , 連 接 到pMD19-T載體(TaKaRa),采用菌液PCR方法鑒定獲得陽性克隆,送上海美吉公司測序。
利用DNAMAN7進行同源性分析,利用Clustalx1.83軟件進行多序列比對,利用MEGA4.1軟件構建系統發生樹。
1.4 實時定量PCR
實時定量PCR(Real-time quantitative PCR,RT-PCR)使用Maxima SYBR Green/ROX qPCR Master Mix(2×)試劑盒(Fermentas),GbWRKY40引物序列為,GbW40-qF:5'-AGCCTGATTGTGAGAGATGGAT-3'和GbW40-qR:5'-TTTCTTGACGGGACAACTTGG-3'。GbUBQ7為內參基因,引物序列為,UBQ7-F:5'-GACCTACACCAAGCCCAAGAAG-3'和UBQ7-R:5'-TGAGCCCACACTTACC ACAATAGT-3'。按照試劑盒說明書推薦體系和反應程序操作。使用ABI 7500 Fast實時熒光定量PCR系統進行實時定量PCR。基因的相對表達量采用方法分析。試驗進行3次生物學重復,每組生物學重復做3次技術重復。數據采用SPSS 17.0(SPSS Inc.,USA)最小顯著差數法(Least significant difference,LSD)進行差異顯著性分析。
1.5 酵母轉化
使用帶有EcoRⅠ和BamHⅠ酶切位點的引物擴增GbWRKY40的開放閱讀框序列,引物序列為:w40-F:5'-TTTGAATTCATGGAATCGACTTGGGTGGATACGA-3',w40-R:5'-TTTGGATCCTTACCACTTGTGATCTAGAACTTTTCCAGA-3',下劃線表示酶切位點。凝膠回收帶有酶切位點GbWRKY40的開放閱讀框序列,雙酶切后亞克隆至pGBKT7載體中,通過測序驗證載體構建的正確性。按照Frozen-EZ Yeast TransformationⅡ試劑盒(ZYMO RESEARCH,Orange,CA)說明書操作步驟,將GbWRKY40-pGBKT7、pGBKT7、陽性對照GmNAC115-pGBKT7質粒[24]轉化酵母菌株AH109,涂布于單缺SD/-Trp營養缺陷型培養基(SD/-Trp)和三缺SD/-Trp/-His/-Ade營養缺陷型培養基 (SD/-Trp/-His/-Ade),30℃培養2~3 d后,觀察菌落的生長情況。
2 結果與分析
2.1GbWRKY40基因全長cDNA克隆及序列分析
根據本課題組海島棉胚珠轉錄組和纖維不同發育表達譜數據得到的棉花WRKY相關unigene序列CL2113.Contig3_All,用RT-PCR方法,獲得全長cDNA序列,將其命名為GbWRKY40。GbWRKY40全長cDNA為1 713 bp,5'端非編碼區261 bp,3'非編碼區為510 bp;編碼區開放閱讀框長942 bp,編碼313個氨基酸,預測相對分子質量約為34.138×103,等電點為8.46。氨基酸序列分析表明該蛋白含有1個WRKY保守區,位于第 156~212個 氨基酸 ,1個鋅 指 基 序C-X5-C-X23-H-X1-H,3個核定位信號區,暗示該轉錄因子可能定位于細胞核(圖1)。系統發生樹分析表明GbWRKY40屬于WRKY家族第Ⅱ類a組(圖 2)。
同源性分析表明,GbWRKY40與TcWRKY40、CcWRKY40、PtWRKY40和PeWRKY的氨基酸序列一致性分別為72.87%,55.86%,56.48%和55.69%。在棉屬中,GbWRKY40與同為四倍體的陸地棉GhWRKY40的氨基酸序列一致性最高(99.04%),與亞洲棉GaWRKY40和草棉 Gr-WRKY40的氨基酸序列一致性分別為81.96%和95.21%。
海島棉基因組數據庫比對發現,GbWRKY40基因定位于 Gbscaffold17271.6.0 scaffold17271: 72937~74804(+strand),GbWRKY40基因全長1868 bp,包含5個外顯子和4個內含子。
2.2 GbWRKY40的表達模式分析
利用熒光定量PCR方法分析GbWRKY40在海島棉不同組織中的表達情況,結果表明在所檢測的組織中GbWRKY40在根中表達量最高,在莖中最低,其他組織中的表達量依次為葉、花和苞葉(圖3),暗示該基因在這些組織中的表達水平可能與這些組織行使的生物學功能有關。GbWRKY40在開花后5 d的纖維中表達量最低,在開花后25 d的纖維中表達量最高(圖4),推測該基因可能在棉纖維次生壁合成過程中發揮作用。

圖1 GbWRKY40與其他已知WRKY蛋白的氨基酸序列比對Fig.1 Alignment of the amino acid sequences of GbWRKY40 and other known WRKY homologs proteins
2.3 GbWRKY40轉錄激活活性分析
用GbWRKY40-pGBKT7、pGBKT7和Gm-NAC115-pGBKT7轉化酵母菌株AH109,并將其涂布于單缺SD/-Trp營養型缺陷培養基后觀察發現,轉化GbWRKY40-pGBKT7、pGBKT7和Gm-NAC115-pGBKT7的酵母菌落都能正常生長 (圖5),說明上述3個質粒均已轉化到酵母中。而在三缺SD/-Trp/-His/-Ade營養型缺陷培養基上,轉化GbWRKY40-pGBKT7的酵母菌落和陰性對照pGBKT7的酵母菌落均不能正常生長,而陽性對照GmNAC115-pGBKT7的酵母菌落能正常生長(圖5),說明GbWRKY40不具有轉錄激活活性,推測GbWRKY40可能是1個轉錄抑制子。
3 討論
本研究從海島棉中克隆了1個WRKY類轉錄因子基因GbWRKY40,氨基酸序列和進化樹分析發現GbWRKY40含有1個WRKY保守區和 1個鋅指基序 C-X5-C-X23-H-X1-H,屬于WRKY轉錄因子家族第Ⅱ類a組。不同物種的同一亞族成員間可能具有類似的功能,GbWRKY40與擬南芥AtWRKY18/40/60同屬WRKY家族第Ⅱ類a組,之前研究發現AtWRKY18/40/60形成1個高度互作的調控網絡,通過轉錄激活或抑制調控植物防御和非生物脅迫應答基因的表達,在植物逆境脅迫應答中具有重要的功能[25-26]。Gb-WRKY40轉錄因子是否參與調控植物逆境脅迫響應需要進一步研究。
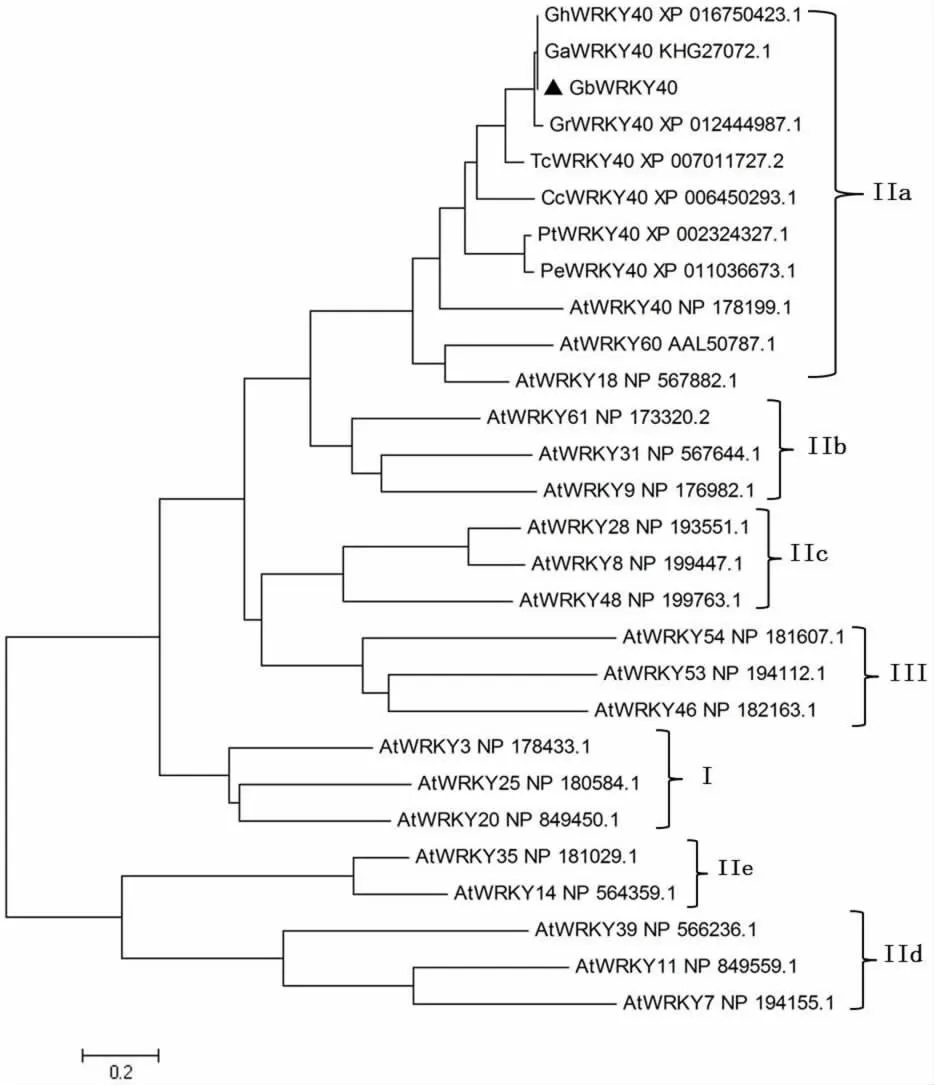
圖2 GbWRKY40蛋白系統發生樹分析Fig.2 Phylogenetic analysis of the GbWRKY40 protein
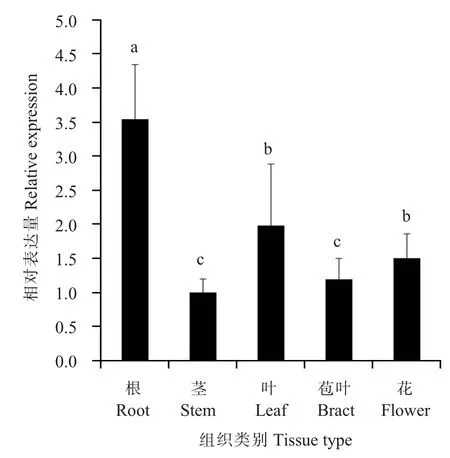
圖3GbWRKY40基因在不同組織中的表達模式Fig.3 Expression profile ofGbWRKY40 gene in different tissues
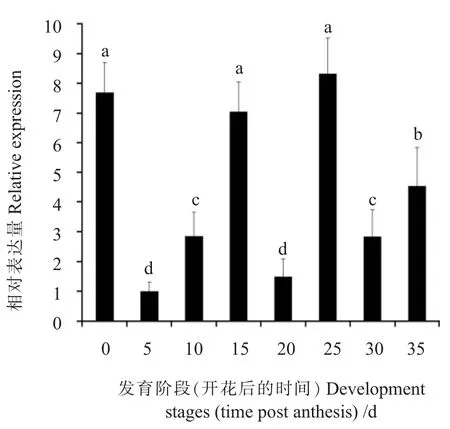
圖4GbWRKY40在不同纖維發育階段的表達模式Fig.4 Expression profile ofGbWRKY40 gene at different fiber developmental stages

圖5 GbWRKY40轉錄因子的轉錄激活試驗Fig.5 Transcriptional activation ability assay of the GbWRKY40 transcription factor
前人研究表明棉花不同的WRKY轉錄因子可以作為轉錄激活子或抑制子調控下游基因的表達,對新克隆的WRKY轉錄因子進行轉錄激活活性分析有助于了解WRKY轉錄因子的調控功能。酵母系統是常用的研究轉錄因子轉錄激活活性的體內研究方法。例如,通過轉化酵母細胞發現GhWRKY31/33[27]、GhWRKY40[19]、GhWRKY41[11]具有轉錄激活活性,而GhWRKY12/16/32/34[27]和海島棉GbWRKY32[28]不具有轉錄激活效應。本研究通過酵母系統發現GbWRKY40不具有轉錄激活活性,與GbWRKY40屬于同一亞組的擬南芥AtWRKY40是轉錄抑制子,而AtWRKY18/60是弱轉錄激活子[26];因此,推測GbWRKY40可能是1個轉錄抑制子。下一步將通過擬南芥原生質體系統驗證GbWRKY40對下游基因的抑制作用。
目前,已報道的棉花WRKY轉錄因子主要參與逆境脅迫應答和衰老進程[14-20,29],參與棉纖維發育的WRKY轉錄因子報道不多。Wan等研究表明棉纖維的發育可能與擬南芥的表皮毛發育具有類似的分子機制[30],擬南芥TTG2編碼1個WRKY轉錄因子,激活下游表皮毛起始基因TTG1和GL1[31],這些研究表明WRKY轉錄因子可能在棉纖維發育過程中具有一定的功能。周立研究發現GhWRKY12和GhWRKY16在開花后3~10 d的棉纖維中優勢表達,推測這些基因可能在棉纖維發育的起始和伸長期發揮一定作用[27]。本研究發現GbWRKY40在根和開花后25 d的纖維中優勢表達,推測該基因可能在棉纖維次生壁發育中具有一定的功能,下一步將通過轉化海島棉來研究基因的生物學功能。
4 結論
從海島棉中克隆了1個WRKY轉錄因子基因GbWRKY40,其不具有轉錄激活活性,在根和開花后25 d的纖維中表達量高。推測該基因可能參與調控棉纖維次生壁發育。
[1]Eulgem T,Rushton P J,Robatzek S,et al.The WRKY superfamily of plant transcription factors[J].Trends in Plant Science,2000, 5(5):199-206.
[2]Wang Kunbo,Wang Zhiwen,Li Fuguang,et al.The draft genome of a diploid cottonGossypium raimondii[J].Nature Genetics, 2012,44(10):1098-1103.
[3]Li Fuguang,Fan Guangyi,Wang Kunbo,et al.Genome sequence of the cultivated cottonGossypium arboreum[J].Nature Genetics,2014,46(6):567-572.
[4]Li Fuguang,Fan Guangyi,Lu Cairui,et al.Genome sequence of cultivated upland cotton (Gossypium hirsutumTM-1)provides insights into genome evolution[J].Nature Biotechnology,2015, 33(5):524-530.
[5]Zhang Tianzhen,Hu Yan,Jiang Wenkai,et al.Sequencing of allotetraploid cotton(Gossypium hirsutumL.acc.TM-1)provides a resource for fiber improvement[J].Nature Biotechnology,2015, 33(5):531-537.
[6]Yuan Daojun,Tang Zhonghui,Wang Maojun,et al.The genome sequence of sea-island cotton(Gossypium barbadense)provides insights into the allopolyploidization and development of superior spinnable fibres[J/OL].Scientific Reports,2015,5:17662 [2016-10-12].https://www.scienceopen.com/document_file/ab63 1975-4060-4707-85cf-cf20fd7f6b1a/PubMedCentral/ab631975-4060-4707-85cf-cf20fd7f6b1a.pdf.DOI:10.1038/srep17662.
[7]Liu Xia,Zhao Bo,Zheng Huajun,et al.Gossypium barbadensegenome sequence provides insight into the evolution of extra-long staple fiber and specialized metabolites[J/OL].Scientific Reports,2015,5:14139[2016-10-12].https://www.scienceopen.com/document_file/81afb1e3-a417-4978-9837-e810d6c78d89/PubMedCentral/81afb1e3-a417-4978-9837-e810d6c78d89.pdf.DOI:10.1038/srep14139.
[8]Ding Mingquan,Chen Jiadong,Jiang Yurong,et al.Genomewide investigation and transcriptome analysis of theWRKYgene family inGossypium[J].Molecular Genetics Genomics,2015, 290(1):151-171.
[9]Dou Lingling,Zhang Xiaohong,Pang Chaoyou,et al.Genomewide analysis of theWRKYgene family in cotton[J].Molecular Genetics and Genomics,2014,289(6):1103-1121.
[10]Cai Caiping,Niu Erli,Du Hao,et al.Genome-wide analysis of the WRKY transcription factor gene family inGossypium raimondiiand the expression of orthologs in cultivated tetraploid cotton[J].The Crop Journal,2014,2(2/3):87-101.
[11]Chu Xiaoqian,Wang Chen,Chen Xiaobo,et al.The cottonWRKYgeneGhWRKY41 positively regulates salt and drought stress tolerance in transgenicNicotiana benthamiana[J/OL].PLoS ONE,2015,10(11):e0143022[2016-10-12].http://dx.doi.org/10.1371/journal.pone.0143022.
[12]Zhou Li,Wang Nana,Gong Siying,et al.Overexpression of a cotton(Gossypium hirsutum)WRKYgene,GhWRKY34,inArabidopsisenhances salt-tolerance of the transgenic plants[J].Plant Physiology and Biochemistry,2015,96:311-320.
[13]Shi Weina,Hao Lili,Li Jing,et al.TheGossypium hirsutum WRKYgeneGhWRKY39-1 promotes pathogen infection defense responses and mediates salt stress tolerance in transgenicNicotiana benthamiana[J].Plant Cell Reports,2014,33(3):483-498.
[14]Yan Yan,Jia Haihong,Wang Fang,et al.Overexpression ofGh-WRKY27areduces tolerance to drought stress and resistance toRhizoctonia solaniinfection in transgenicNicotiana benthamiana[J/OL].Frontiers in Physiology,2015,6:265[2016-10-12].http://dx.doi.org/10.3389/fphys.2015.00265.
[15]Jia Haihong,Wang Chen,Wang Fang,et al.GhWRKY68 reduces resistance to salt and drought in transgenicNicotiana benthamiana[J/OL].PLoS ONE,2015,10(3):e0120646[2016-10-12].http://journals.plos.org/plosone/article/asset?id=10.1371/journal.pone.0120646.PDF.
[16]Yan Huiru,Jia Haihong,Chen Xiaobo,et al.The cotton WRKY transcription factor GhWRKY17 functions in drought and saltstress in transgenicNicotiana benthamianathrough ABA signaling and the modulation of reactive oxygen species production[J].Plant and Cell Physiology,2014,55(12):2060-2076.
[17]Yu Feifei,Huaxia Yifeng,Lu Wenjing,et al.GhWRKY15,a member of the WRKY transcription factor family identified from cotton(Gossypium hirsutumL.),is involved in disease resistance and plant development[J/OL].BMC Plant Biology, 2012,12:144[2016-10-12].http://www.biomedcentral.com/1471-2229/12/144.DOI:10.1186/1471-2229-12-144.
[18]Liu Xiufang,Song Yunzhi,Xing Fangyu,et al.GhWRKY25,a groupⅠ WRKY gene from cotton,confers differential tolerance to abiotic and biotic stresses in transgenicNicotiana benthamiana[J].Protoplasma,2016,253(5):1265-1281.
[19]Wang Xiuling,Yan Yan,Li Yuzhen,et al.GhWRKY40,a multiple stress-responsive cottonWRKYgene,plays an important role in the wounding response and enhances susceptibility toRalstonia solanacearuminfection in transgenicNicotiana benthamiana[J/OL].PLoS ONE,2014,9(4):e93577[2016-10-12].http://journals.plos.org/plosone/article/asset?id=10.1371/journal.pone.0093577.PDF.
[20]竇玲玲,李光雷,龐朝友,等.棉花轉錄因子GhWRKY11的克隆及功能分析[J].農業生物技術學報,2016,24(5):625-636.Dou Lingling,Li Guanglei,Pang Chaoyou,et al.Cloning and function analysis ofGhWRKY11 in cotton (Gossypium hirsutum)[J].Journal of Agricultural Biotechnology,2016,24(5): 625-636.
[21]Fan Xinqi,Guo Qi,Xu Peng,et al.Transcriptome-wide identification of salt-responsive members of theWRKYgene family inGossypium aridum[J/OL].PLoS ONE,2015,10(5):e0126148 [2016-10-12].http://journals.plos.org/plosone/article/asset?id= 10.1371/journal.pone.0126148.PDF.
[22]Xu Yanhua,Wang Jiawei,Wang Shui,et al.Characterization of GaWRKY1,a cotton transcription factor that regulates the sesquiterpene synthase gene(+)-delta-cadinene synthase-A[J].Plant Physiology,2004,135(1):507-515.
[23]Xu Li,Jin Li,Long Lu,et al.Overexpression ofGbWRKY1 positively regulates the Pi starvation response by alteration of auxin sensitivity inArabidopsis[J].Plant Cell Reports,2012,31 (12):2177-2188.
[24]倪志勇,于月華,陳全家,等.大豆GmNAC115基因克隆及特征分析[J].大豆科學,2016,35(5):16-20.Ni Zhiyong,Yu Yuehua,Chen Quanjia,et al.Cloning and characterization ofGmNAC115 gene in soybean[J].Soybean Science,2016,35(5):16-20.
[25]Xu Xinping,Chen Chunhong,Fan Baofang,et al.Physical and functional interactions between pathogen-induced Arabidopsis WRKY18,WRKY40,and WRKY60 transcription factors[J].Plant Cell,2006,18(5):1310-1326.
[26]Chen Han,Lai Zhibing,Shi Junwei,et al.Roles ofArabidopsisWRKY18,WRKY40 and WRKY60 transcription factors in plant responses to abscisic acid and abiotic stress[J/OL].BMC Plant Biology,2010,10:281[2016-10-12].http://www.biomedcentral.com/1471-2229/10/281.DOI:10.1186/1471-2229-10-281.
[27]周立.棉花(Gossypium hirsutum)WRKY基因分離與鑒定[D].武漢:華中師范大學,2014.Zhou Li.Isolation and characterization ofWRKYgenes in cotton(Gossypium hirsutum)[D].Wuhan:Central China Normal University,2014.
[28]加得拉·吐留汗,倪志勇,邱迎風,等.海島棉GbWRKY32基因的克隆及特性分析[J].分子植物育種,2016,14(6):1361-1368.Jadera Toluhan,Ni Zhiyong,Qiu Yingfeng,et al.Cloning and characterization ofGbWRKY32 in sea island cotton(Gossypium barbadenseL.)[J].Molecular Plant Breeding,2016,14(6): 1361-1368.
[29]李光雷,范術麗,宋美珍,等.棉花轉錄因子GhWRKY4基因的克隆及特征分析[J].棉花學報,2013,25(3):205-210.Li Guanglei,Fan Shuli,Song Meizhen,et al.Molecular cloning and characterization analysis ofGhWRKY4,a transcription factor gene in cotton(Gossypium hirsutumL.)[J].Cotton Science, 2013,25(3):205-210.
[30]Wan Qun,Zhang Hua,Ye Wenxue,et al.Genome-wide transcriptome profiling revealed cotton fuzz fiber development having a similar molecular model asArabidopsistrichome[J/OL].PLoS ONE,2014,9(5):e97313 (2014-05-13)[2016-10-12].http://journals.plos.org/plosone/article/file?id=10.1371/journal.pone.0097313.PDF.
[31]Johnson C S,Kolevski B,Smyth D R.TRANSPARENT TESTA GLABRA2,a trichome and seed coat development gene ofArabidopsis,encodesa WRKYtranscription factor[J].Plant Cell, 2002,14(6):1359-1375.
Cloning and Characterization of theGbWRKY40 Transcription Factor Gene fromGossypium barbadenseL.
Ni Zhiyong,Jiadela Tuliuhan,Qiu Yingfeng,Qu Yanying,Chen Quanjia*
(College of Agronomy,Xinjiang Agricultural University/Key Laboratory of Agricultural Biological Technology,Xinjiang Agricultural University,Urumqi830052,China)
[Objective]The WRKY transcription factors modulate various biological processes,including plant growth,development,and responses to various environmental stresses.This study aimed to clarify the role of WRKY transcription factors related to cotton fiber development.[Method]TheGbWRKY40 gene,which encodes a WRKY transcription factor,was isolated from cotton(Gossypium barbadenseL.).The obtained sequence was analyzed for homology with genes from other cotton species using the DNAMAN7 program.The sequences of the homologous genes were aligned with Clustalx1.83.Additionally,theGb-WRKY40 expression pattern was analyzed by a quantitative real-time polymerase chain reaction.Furthermore,a yeast expression vector was constructed and inserted into yeast strain AH109 cells to investigate the transcriptional activity ofGbWRKY40.[Result]Full-lengthGbWRKY40 cDNA contains 1713 nucleotides,including a 942 bp open reading frame,a 261 bp 5'-untranslated region,and a 510 bp 3'-untranslated region.The open reading frame was predicted to encode a 313 amino acid protein with a relative molecular mass of 34.138×103and an isoelectric point of 8.46.The genomicGbWRKY40 sequence was observed to comprise five exons and four introns.The predicted GbWRKY40 protein includes a WRKY domain consisting of approximately 60 amino acids,including the conserved WRKYGQK sequence and a zinc-finger motif(C-X5-C-X23-H-X1-H).These characteristics indicated that GbWRKY40 belongs to group IIa of the WRKY family.Moreover,GbWRKY40 contains three putative nuclear localization signals.GbWRKY40 is highly homologous toGhWRKY40.Expression analyses involving a quantitative real-time polymerase chain reaction revealed thatGbWRKY40 is highly expressed in cotton roots and fibers at 25 days post anthesis.Transcriptional activation assay results suggested that the function of the GbWRKY40 transcription factor is unrelated to the activation of transcription.[Conclusion]These results imply thatGbWRKY40 may be involved in regulating secondary cell walldevelopment in cotton fibers.
cotton;GbWRKY40;expression profile;gene clone;transcriptional activation
S562.035
A
1002-7807(2017)04-0393-08
10.11963/1002-7807.nzycqj.20170601
2016-10-13
倪志勇(1981―),男,博士,nizhiyong@126.com。*通信作者:chqjia@126.com
新疆維吾爾自治區青年科技創新人才培養工程優秀青年科技人才(qn2015yx012)
