普通野生稻綠色組織特異表達(dá)啟動子的克隆與鑒定
2017-08-07 23:22:05趙志強薛滿德黃珂張靜文龍艷裴新梧袁潛華
生物技術(shù)通報 2017年8期
關(guān)鍵詞:綠色
趙志強薛滿德黃珂張靜文龍艷裴新梧袁潛華
(1. 海南大學(xué)熱帶農(nóng)林學(xué)院,海口 570228;2. 中國農(nóng)業(yè)科學(xué)院生物技術(shù)研究所,北京 100081)
普通野生稻綠色組織特異表達(dá)啟動子的克隆與鑒定
趙志強1薛滿德2黃珂1張靜文2龍艷2裴新梧2袁潛華1
(1. 海南大學(xué)熱帶農(nóng)林學(xué)院,海口 570228;2. 中國農(nóng)業(yè)科學(xué)院生物技術(shù)研究所,北京 100081)
綠色組織特異表達(dá)啟動子可調(diào)控外源基因只在受體作物的綠色組織中定點、高效地表達(dá)。以普通野生稻為實驗材料,克隆了綠色組織特異表達(dá)啟動子OrGSP,構(gòu)建OrGSP和GUS基因融合的表達(dá)載體,轉(zhuǎn)入擬南芥中鑒定功能。啟動子OrGSP長度為825 bp,含有基本的轉(zhuǎn)錄起始元件TATA-box和CAAT-box,以及光響應(yīng)元件TCCC-motif、Sp1、G-box、I-box、GA-motif和as-2-box等。轉(zhuǎn)基因擬南芥GUS組織化學(xué)染色結(jié)果表明,啟動子OrGSP調(diào)控GUS基因只在綠色組織中特異表達(dá)。GUS活性測定結(jié)果顯示,葉和莖中的GUS活性比根中明顯提高。普通野生稻中克隆的啟動子OrGSP為綠色組織特異表達(dá)啟動子,可為作物分子育種提供新的調(diào)控元件。
啟動子;綠色組織特異表達(dá);普通野生稻
啟動子是基因的重要組成部分,它就像“開關(guān)”,調(diào)節(jié)著下游基因的活性。啟動子是指位于結(jié)構(gòu)基因5′端上游的一段DNA序列,它與 RNA 聚合酶作用后開始基因表達(dá)的轉(zhuǎn)錄過程。RNA聚合酶只有和啟動子結(jié)合才可以開始基因的轉(zhuǎn)錄過程,啟動子是基因轉(zhuǎn)錄調(diào)控重要的順式作用元件[1]。
綠色組織在植物的生長發(fā)育過程中起著重要的作用。光合作用是植物的重要生理過程,植物的葉片是光合作用的場所,植物利用葉片中的葉綠素等光合色素,在可見光的照射下,將二氧化碳和水轉(zhuǎn)化為有機物,并釋放出氧氣。蒸騰作用也發(fā)生在植物的葉片組織,蒸騰作用促進(jìn)植物從根系吸收水分和礦質(zhì)元素,同時降低植物葉片的葉面溫度。植物的莖是植物體內(nèi)物質(zhì)運輸?shù)闹饕ǖ溃参锶~片制造的有機物通過莖輸送到植物體的各部分,同時根部吸收的水分和無機物也通過莖運輸?shù)街参锏牟煌课弧V参锏木G色組織也是病蟲侵害的主要部位。綠色組織特異表達(dá)啟動子的克隆,能夠使外源基因在受體作物中定點、高效地表達(dá)[2]。
普通野生稻(Oryza rufipogon)是亞洲栽培稻(Oryza sativa)的近緣祖先種,具有豐富的遺傳多樣性,是栽培稻改良的重要種質(zhì)資源[3]。本研究從普通野生稻基因組中克隆了綠色組織特異表達(dá)啟動子OrGSP,并構(gòu)建表達(dá)載體轉(zhuǎn)化擬南芥驗證其功能,為作物分子育種提供了新的調(diào)控元件。
1 材料與方法
1.1 材料
1.1.1 植物材料 普通野生稻(由本實驗室保存),擬南芥哥倫比亞生態(tài)型。
1.1.2 載體與菌株 pBinGlyRed3-35S-GUS載體(由本實驗室保存),pEASY-T1 Simple克隆載體(Transgen Biotech),農(nóng)桿菌EHA105,Trans-T1 Phage Resistan化學(xué)感受多態(tài)細(xì)胞(Trans Biotech)。
1.2 方法
1.2.1 OrGSG基因表達(dá)模式的驗證 在本實驗室前期構(gòu)建的普通野生稻cDNA文庫中篩選到1個FPKM值在苗期葉中遠(yuǎn)大于根的基因(苗期葉:3682.44033;苗期根:2.214723825),命名為OrGSG。分別提取普通野生稻苗期的根、葉和成熟期的根、莖、葉的總RNA,反轉(zhuǎn)錄成cDNA。以cDNA文庫中預(yù)測的OrGSG基因的CDS序列為模板,設(shè)計引物(FP:5′-TTCACCAGCTTCACT CGCC-3′,RP:5′-ATCGAAACCGAAGTCTCCC-3′)。通過RTPCR,驗證OrGSG基因在苗期的根、葉和成熟期的根、莖、葉中的表達(dá)模式,選用OsActin(No.AB047313)作為內(nèi)參基因(FP:5′-TTGTGTTGGACTCTGGTGATG-3′,RP:5′-AAGCTCGTAGCTCTTCTCCAC-3′),設(shè)置26、28和30循環(huán)數(shù)梯度。
1.2.2 OrGSG基因啟動子的克隆及序列分析 將 OrGSG基因的CDS序列在NCBI(https://www. ncbi. nlm.nih.gov/)數(shù)據(jù)庫進(jìn)行Blast比對,得到OrGSG基因的CDS序列比對上的粳稻染色體以及基因。OrGSG基因的啟動子命名為OrGSP。比對上的完整基因ATG起始密碼子A堿基下游部分序列和上游部分序列為克隆OrGSP啟動子的模板,OrGSP啟動子擴(kuò)增引物(FP:5′-ATCTCAT GAAGTTTAGCTTGCCG-3′,RP:5′-GAAGACGATGAAATAGATGGGTG-3′)。 使用 PlantCARE在 線 軟 件(http://bioinformatics.psb. ugent.be/we)分析啟動子的特征元件。
1.2.3 OrGSP-GUS表達(dá)載體的構(gòu)建 用加接頭的引 物(FP:5′-ACGCGTAAGGGGATCC ATCTCAT GAAGTTTAGCTTGCCG-3′,RP:5′-GATCTACCATG AATTCGAAGACGATGAAATAGATGGGTG-3′)從普通野生稻基因組DNA中克隆帶接頭序列的OrGSP啟動子,其中GGATCC為Bam HI酶切位點,GAATTC為EcoRI酶切位點。將OrGSP啟動子和pBinGly-Red3-35S-GUS表達(dá)載體骨架進(jìn)行重組,得到OrGSPGUS融合的pBinGlyRed3-OrGSP-GUS表達(dá)載體。
1.2.4 擬南芥遺傳轉(zhuǎn)化及分子鑒定 利用凍融法將pBinGlyRed3-35S-GUS和 pBinGlyRed3 -OrGSP-GUS表達(dá)載體轉(zhuǎn)化到根癌農(nóng)桿菌EHA105中,其中含35S啟動子的pBinGlyRed3-35S-GUS表達(dá)載體作為對照。將這兩種表達(dá)載體通過農(nóng)桿菌介導(dǎo)的蘸花法轉(zhuǎn)化擬南芥。pBinGlyRed3-35S -GUS和pBinGlyRed3-OrGSP-GUS表達(dá)載體都含有紅光標(biāo)記基因,分別轉(zhuǎn)化擬南芥后收獲成熟的T0代種子,在綠光燈下,用紅色眼鏡挑選種子,各挑選顯示紅色的轉(zhuǎn)基因T0代種子20粒,記為20個株系。當(dāng)T1代幼苗一個月后取葉片提取基因組DNA,在啟動子區(qū)域和GUS基因編碼區(qū)設(shè)計引物進(jìn)行PCR擴(kuò)增驗證。其中轉(zhuǎn)pBinGlyRed3-35S-GUS表達(dá)載體T1代幼苗PCR鑒定引 物 為(FP:5′-gtaagg gatgacgcacaatcc-3′,RP:5′-ggtcgtgtagattttcaccgg-3′), 轉(zhuǎn) pBinGlyRed3- OrGSP -GUS表達(dá)載體T1代幼苗PCR鑒定引物為(FP:5′-cagtacggatgcaatgtgtgt-3′,RP:5′-cag cttgctttcgtaccactt-3′)。
1.2.5 轉(zhuǎn)基因擬南芥的GUS組織化學(xué)分析 轉(zhuǎn)基因陽性擬南芥T1代種子取一部分在1/2MS培養(yǎng)基上培養(yǎng)。分別取4 d、7 d、14 d、21 d的擬南芥進(jìn)行GUS染色分析,GUS染色方法見Zhou等[4];轉(zhuǎn)基因陽性擬南芥T1代種子再取一部分在培養(yǎng)基質(zhì)(珍珠巖:蛭石:擬南芥種植專用土體積比1∶1∶2)中培養(yǎng),待其開花后,取其根、莖、葉、花、莢果進(jìn)行GUS染色分析。
1.2.6 轉(zhuǎn)基因擬南芥的GUS活性分析 選5個株系的T1代轉(zhuǎn)基因陽性擬南芥種子,在培養(yǎng)基質(zhì)中培養(yǎng)。一個月后分別取苗期的根和葉,待擬南芥開花后取成熟期的根、莖和葉,進(jìn)行GUS活性檢測,GUS活性檢測方法見文獻(xiàn)[5]。
2 結(jié)果
2.1 OrGSG基因表達(dá)模式驗證與序列分析
通過RT-PCR驗證OrGSG基因在不同發(fā)育時期的根、莖、葉的表達(dá)模式,結(jié)果(圖1)表明該基因在幼苗的葉中表達(dá),在根中不表達(dá),在成熟期的莖和葉中表達(dá),在根中不表達(dá)。
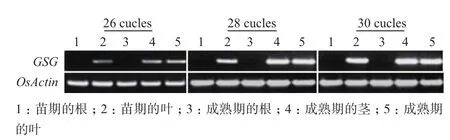
圖1 OrGSG基因表達(dá)模式的循環(huán)數(shù)梯度RT-PCR驗證
OrGSG基因的CDS序列99%比對到粳稻9號染色體26810號基因上的16288855到16289372位置。26810號基因長度為2 947 bp,可以編碼265個氨基酸,預(yù)測功能為可以合成捕光葉綠素a/b結(jié)合蛋白。
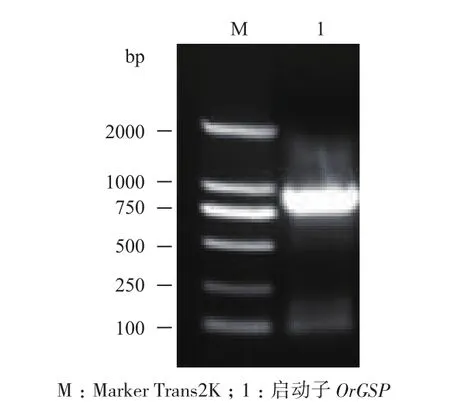
圖2 啟動子OrGSP的PCR產(chǎn)物電泳結(jié)果
2.2 啟動子OrGSP的克隆與序列分析
以普通野生稻基因組 DNA 為模板,擴(kuò)增OrGSG基因的啟動子序列,擴(kuò)增出 1 條約 825 bp的條帶(圖2),與預(yù)期大小一致。
啟動子OrGSP長度為825 bp,與比對上的栽培稻序列相似性為100%。對啟動子OrGSP序列的順式作用元件分析,結(jié)果(表1)顯示,該啟動子的正鏈以及互補鏈上存在著多種順式作用元件,包括決定著轉(zhuǎn)錄的起始和方向的核心啟動子元件TATA-box和在調(diào)控基因轉(zhuǎn)錄效率中發(fā)揮著重要作用的基本順式作用元件CAAT-box;6種光響應(yīng)元件,如TCCC-motif、Sp1、G-box、I-box、GA-motif和 as-2-box,其中as-2-box還是莖特異表達(dá)元件;2種激素誘導(dǎo)元件,如水楊酸誘導(dǎo)元件TCA-element和赤霉素誘導(dǎo)元件GARE-motif;1種脅迫防御元件TC-rich repeats。
2.3 pBinGlyRed3-OrGSP-GUS表達(dá)載體的構(gòu)建
將OrGSP啟動子替換pBinGlyRed3-35S-GUS表達(dá)載體上的35S啟動子,構(gòu)建綠色組織特異表達(dá)的表達(dá)載體pBinGlyRed3-OrGSP-GUS(圖3,圖4)。
2.4 轉(zhuǎn)基因擬南芥T1代幼苗PCR檢測
轉(zhuǎn) pBinGlyRed3-35S-GUS和 pBinGlyRed3-OrGSP-GUS表達(dá)載體的擬南芥T0代紅色種子各取20粒,在培養(yǎng)基質(zhì)中培養(yǎng)一個月后,取T1代幼苗取葉片進(jìn)行PCR鑒定。從圖5和圖6可知,轉(zhuǎn)pBinGlyRed3-35S-GUS表達(dá)載體的擬南芥T1代有13個株系(泳道1、2、3、4、5、7、8、9、11、16、18、19和20)擴(kuò)增出大小約為850 bp的片段,為陽性株系;轉(zhuǎn)pBinGlyRed3-OrGSP-GUS表達(dá)載體的擬南芥T1代有15個株系(泳道1、2、3、4、5、6、7、8、11、12、13、14、15、17、19)擴(kuò)增出大小約為850 bp的片段,為陽性株系。
2.5 轉(zhuǎn)基因擬南芥的GUS染色分析
分別對轉(zhuǎn)pBinGlyRed3-35S-GUS和pBinGlyRed3-OrGSP-GUS表達(dá)載體的陽性擬南芥T2代株系進(jìn)行GUS染色,染色材料為:4 d、7 d、14 d和21 d時的幼苗整株和成熟期的擬南芥根、莖、葉、花和果實。結(jié)果(圖7,圖8)顯示:幼苗期時,轉(zhuǎn)pBinGlyRed3-35S-GUS表達(dá)載體的擬南芥植株在4 d、
7 d、14 d和21 d時所有組織中均可以染出顏色,轉(zhuǎn)pBinGlyRed3-OrGSP-GUS表達(dá)載體的擬南芥植株在4 d、7 d、14 d和21 d時的根部染不出顏色,其余組織中可以染出顏色;成熟期時,轉(zhuǎn)pBinGlyRed3-35S-GUS表達(dá)載體的擬南芥的根、莖、葉、花和角果中都能染出顏色,轉(zhuǎn)pBinGlyRed3-OrGSP-GUS表達(dá)載體的擬南芥的根、花和角果染不出顏色,莖和葉中可以染出顏色。這就表明OrGSP啟動子在幼苗期和成熟期時都可以調(diào)控GUS基因在綠色組織中表達(dá),OrGSP啟動子為綠色組織特異表達(dá)啟動子,OrGSP調(diào)控的基因OrGSG為綠色組織特異表達(dá)基因。2.6 轉(zhuǎn)基因擬南芥GUS活性檢測

表1 啟動子OrGSP順式作用元件預(yù)測

圖3 pBinGlyRed3-35S-GUS表達(dá)載體圖譜
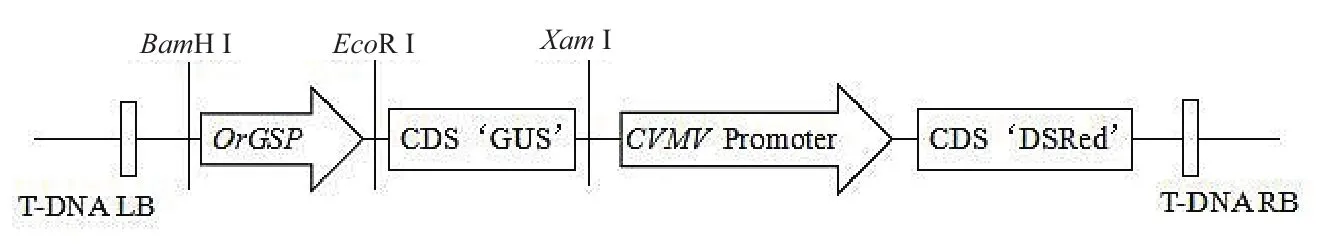
圖4 pBinGlyRed3-OrGSP-GUS表達(dá)載體圖譜
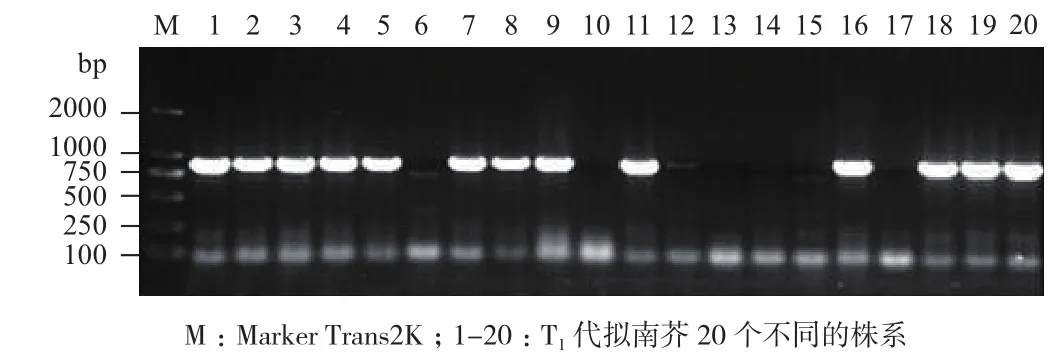
圖5 轉(zhuǎn)pBinGlyRed3-35S-GUS擬南芥T1代幼苗PCR檢測
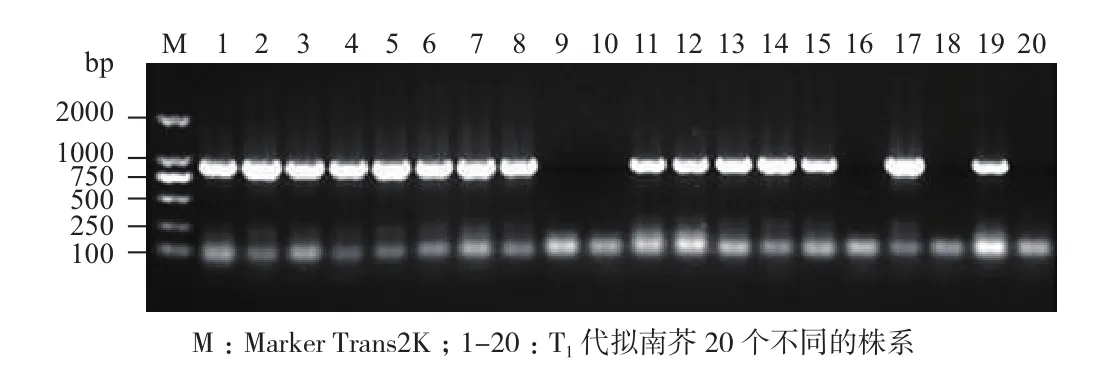
圖6 轉(zhuǎn)pBinGlyRed3-OrGSP-GUS擬南芥T1代幼苗PCR檢測
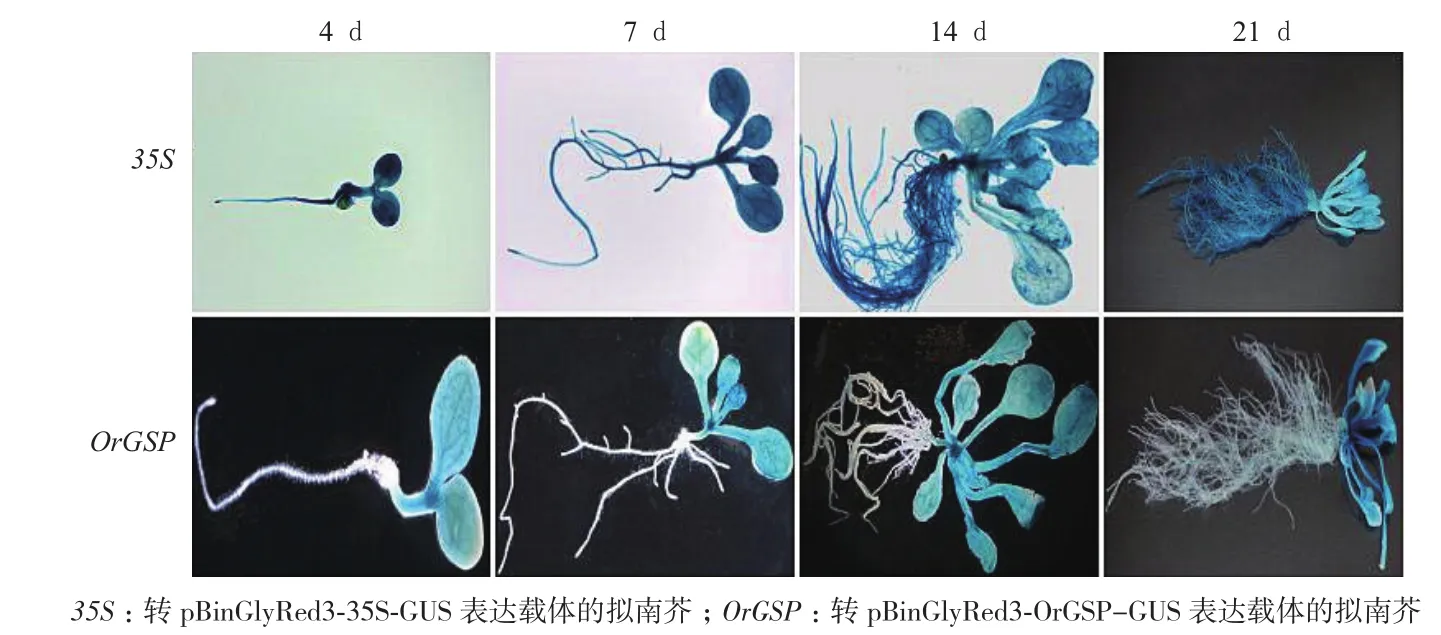
圖7 幼苗期轉(zhuǎn)基因陽性擬南芥GUS染色
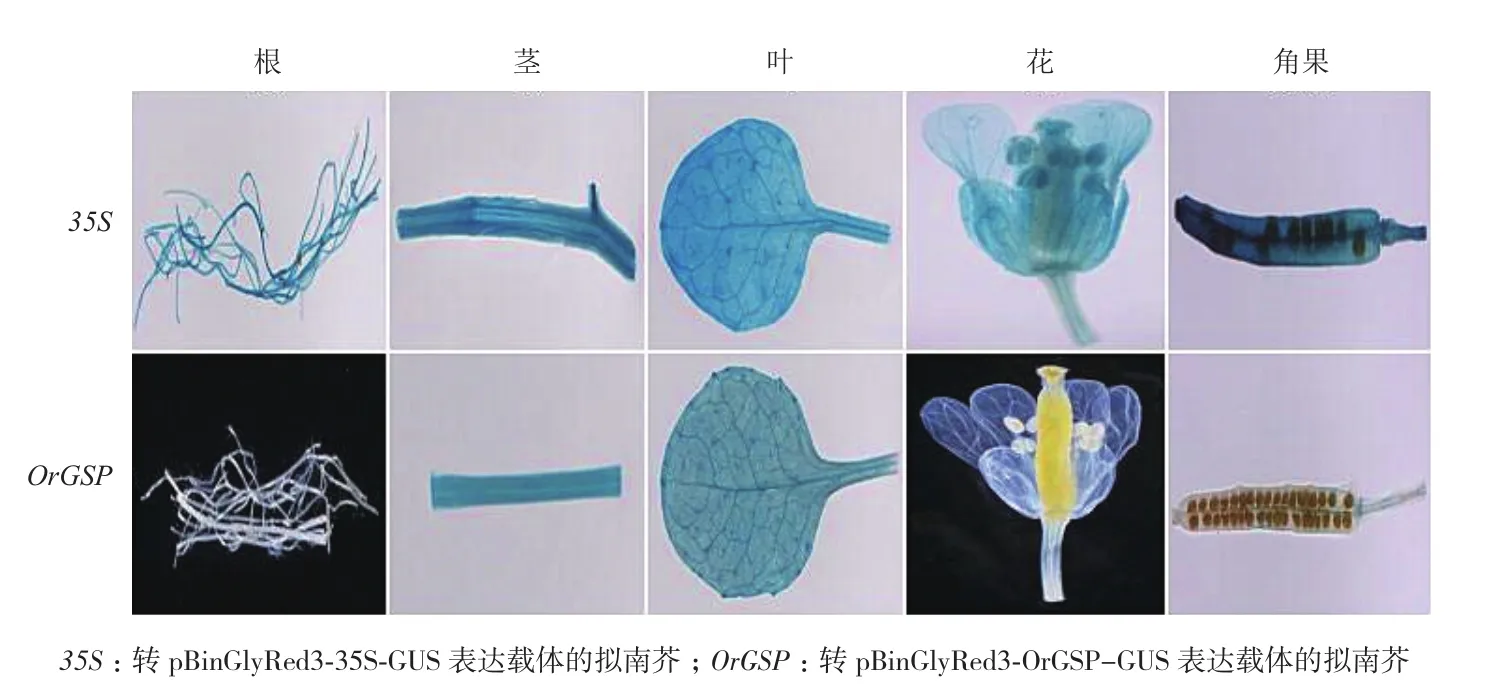
圖8 成熟期轉(zhuǎn)基因陽性擬南芥各組織GUS染色
轉(zhuǎn)pBinGlyRed3-OrGSP-GUS表達(dá)載體的T2代陽性擬南芥苗期葉中的GUS活性遠(yuǎn)比根中高,經(jīng)計算,葉中活性約為根中活性的256倍;在成熟期時,轉(zhuǎn)pBinGlyRed3-OrGSP-GUS表達(dá)載體的T2代陽性擬南芥莖和葉中的GUS活性遠(yuǎn)遠(yuǎn)比根中高,經(jīng)計算,莖中活性約為根中的310倍,葉中活性約為根中的325倍(圖9)。
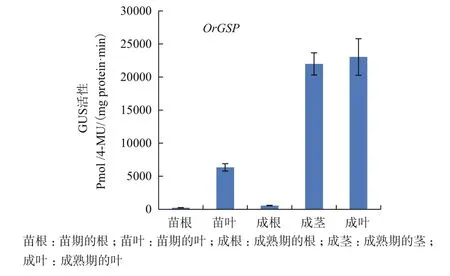
圖9 啟動子OrGSP在轉(zhuǎn)基因陽性擬南芥不同組織中的GUS活性檢測
3 討論
植物綠色組織特異表達(dá)啟動子的克隆,可以為植物基因工程提供調(diào)控元件,使外源基因能在受體作物中定點、高效表達(dá)。近年來,在水稻[6-9]、棉花[10]、馬鈴薯[11]、裂葉牽牛[12]、擬南芥[13]、玉米[14]、番茄[15]等多種植物中克隆出綠色組織特異表達(dá)啟動子。普通野生稻是亞洲栽培稻的祖先種,廣泛分布于東南亞、南亞以及我國的南方地區(qū),具有豐富的遺傳多樣性[16],但對普通野生稻綠色組織特異表達(dá)啟動子的研究還未見報道。
本研究從本實驗室前期構(gòu)建的普通野生稻cDNA文庫中篩選到一個綠色組織特異表達(dá)基因OrGSG。該基因的CDS序列99%比對到粳稻9號染色體26810號基因上的16288855到16289372位置。26810號基因長度為2 947 bp,功能預(yù)測為可以合成捕光葉綠素a/b蛋白。捕光葉綠素a/b結(jié)合蛋白與光合色素所形成的色素蛋白復(fù)合體(LHC)是一類吸收、傳遞光能并引起光合作用的光反應(yīng)的色素蛋白復(fù)合體。色素蛋白復(fù)合體(LHC)在維持葉綠體囊狀結(jié)構(gòu)薄膜結(jié)構(gòu),調(diào)節(jié)光系統(tǒng)I和光系統(tǒng)II之間激發(fā)能的分配以及適應(yīng)不同環(huán)境等方面都起著重要的作用[17]。捕光葉綠素a/b結(jié)合蛋白基因的啟動子之前報道為綠色組織特異表達(dá)啟動子[18],李為民等[19]從金華中棉中克隆得到捕光葉綠素a/b結(jié)合蛋白基因Gacab轉(zhuǎn)錄起始位點上游1 009 bp的啟動子GacabP,構(gòu)建GacabP和GUS基因融合的表達(dá)載體轉(zhuǎn)化煙草,GUS組織化學(xué)分析發(fā)現(xiàn),GacabP調(diào)控GUS基因在轉(zhuǎn)基因陽性煙草的葉片表達(dá),表明GacabP是一個葉特異表達(dá)啟動子。
本研究克隆了普通野生稻綠色組織特異表達(dá)啟動子OrGSP,長度為825 bp,其正鏈及互補鏈上有多種順式作用元件,除含有決定轉(zhuǎn)錄的起始和方向的核心啟動子元件TATA-box和在調(diào)控基因轉(zhuǎn)錄效率中發(fā)揮著重要作用的基本順式作用元件CAAT-box外,還存在一些與光反應(yīng)、激素誘導(dǎo)和脅迫防御相關(guān)的順式作用元件。轉(zhuǎn)OrGSP和GUS基因融合的表達(dá)載體的T2代陽性擬南芥GUS組織化學(xué)分析表明OrGSP為綠色組織特異表達(dá)啟動子。轉(zhuǎn)基因擬南芥植株的GUS活性檢測結(jié)果顯示,在苗期和成熟期時,綠色組織中的GUS活性都比根中明顯提高。
植物的綠色組織是病蟲侵害的主要部位,稻瘟病、紋枯病和白葉枯病主要發(fā)生在水稻的綠色組織中[20-22]。Cai等[8]將水稻綠色組織特異表達(dá)啟動子PD540與cry1AC基因融合的表達(dá)載體轉(zhuǎn)化水稻,結(jié)果顯示,轉(zhuǎn)基因陽性水稻不僅在抗稻縱卷葉螟上有著明顯的優(yōu)勢,而且水稻胚乳中檢測不到cry1AC的表達(dá),說明該啟動子在轉(zhuǎn)基因抗蟲水稻的培育和和轉(zhuǎn)基因生物安全方面有著重要的應(yīng)用價值。Yang等[23]利用合成型綠色組織特異表達(dá)啟動子pGreen和合成型抗蟲基因融合表達(dá)載體轉(zhuǎn)化水稻,結(jié)果表明,轉(zhuǎn)基因陽性水稻對水稻二化螟和棉鈴蟲有著良好的抗性。
4 結(jié)論
本研究克隆了普通野生稻的啟動子OrGSP,在擬南芥中驗證了功能,該啟動子為綠色組織特異表達(dá)啟動子。
[1] Porto M, Pinheiro M, Batista V, et al. Plant promoters:An approach of structure and function[J]. Molecular Biotechnology, 2014, 56(1):38-49.
[2] Chen L, Jiang B, Wu C, et al. The characterization of GmTIP, a root-specific gene from soybean, and the expression analysis of its promoter[J]. Plant Cell, Tissue and Organ Culture, 2015, 121(2):259-274.
[3] Tian X, Long Y, Wang J, et al. De novo transcriptome assembly of common wild rice(Oryza rufipogon Griff. )and discovery of drought-response genes in root tissue based on transcriptomic data[J]. PLoS One, 2015, 10(7):e0131455.
[4] Zhou Y, Huang W, Liu L, et al. Identification and functional characterization of a rice NAC gene involved in the regulation of leaf senescence[J]. BMC Plant Biology, 2013, 13:132.
[5] Banerjee J, Sahoo D, Raha S, et al. A region containing an as-1 element of Dahlia mosaic virus(DaMV)subgenomic transcript promoter plays a key role in green tissue-and root-specific expression in plants[J]. Plant Mol Biol Reporter, 2015, 33(3):532-556.
[6] Ye R, Zhou F, Lin Y. Two novel positive cis-regulatory elements involved in green tissue-specific specific promoter activity in rice(Oryza sativa L ssp. )[J]. Plant Cell Report, 2012, 31(7):1159-1172.
[7] Yang Z, Lu Q, Wen X, et al. Functional analysis of the rice rubisco activase promoter in transgenic Arabidopsis[J]. Biochemical and Biophysical Research Communications, 2012, 418(3):565-570.
[8] Cai M, Wei J, Li X, et al. A rice promoter containing both novel positive and negative cis-elements for regulation of green tissue- specific gene expression in transgenic plants[J]. Plant Biotechnology Journal, 2007, 5(5):664-674.
[9] Thilmony R, Guttman M, James G, et al. The LP2 leucine-rich repeat receptor kinase gene promoter directs organ-specific, lightresponsive expression in transgenic rice[J]. Plant Biotechnology Journal, 2009, 7(9):867-882.
[10] 劉聚波, 張玉娟, 陳潔, 等. 棉花GhRCA1基因啟動子的克隆及序列分析[J]. 分子植物育種, 2014, 12(4):701-711.
[11] Qu D, Song Y, Li W, et al. Isolation and characterization of the organ-specific and light-inducible promoter of the gene encoding rubisco activase in potato(Solanum tuberosum)[J]. Genetics and Molecular Research, 2011, 10(2):621-631.
[12] 楊予濤, 楊國棟, 劉石娟, 等. 一個光合組織特異表達(dá)強啟動子的分離及功能分析[J]. 中國科學(xué)(C輯生命科學(xué)), 2003, 33(4):298-306.
[13] 王利軍, 范三紅, 郭藹光. 擬南芥ats1A基因啟動子的克隆和功能分析[J]. 西北植物學(xué)報, 2004, 24(10):1856-1860.
[14] Taniguchi M, Izawa K, Ku M, et al. The promoter for the maize C4 pyruvate, orthophosphate dikinase gene directs cell-and tissuespecific transcription in transgenic maize plants[J]. Plant Cell Physiology, 2000, 41(1):42-48.
[15] Kyozuka J, McElroy D, Hayakawa T, et al. Light-regulated and cellspecific expression of tomato rbcS-gusA and rice rbcS-gusA fusion genes in transgenic rice[J]. Plant Physiology, 1993, 102(3):991-1000.
[16] Kovach M, Sweeney M, McCouch S. New insights into the history of rice demestication[J]. Trends In Genetics, 2007, 23(11):578-587.
[17] Kong F, Zhou Y, Sun P, et al. Identification of light-harvesting chlorophyll a/b-binding protein genes of zostera marina L. and their expression under different environmental conditions[J]. Oceanic and Coastal Sea Research, 2016, 15(1):152-162.
[18] 王淼, 王旭靜, 唐巧玲, 等. 高等植物綠色組織特異表達(dá)啟動子研究進(jìn)展[J]. 中國農(nóng)業(yè)科技導(dǎo)報, 2010, 12(2):33-37.
[19] 李為民, 王志興, 裴新梧, 等. 中棉光誘導(dǎo)基因Gacab啟動子的克隆及其功能分析[J]. 農(nóng)業(yè)生物技術(shù)學(xué)報, 2004, 12(3):253-257.
[20] 章琦. 中國雜交水稻白葉枯病抗性的遺傳改良[J]. 中國水稻科學(xué), 2009, 23(2):111-119.
[21] 夏更壽. 水稻抗紋枯病基因OsSeh1的克隆及功能鑒定[J].核農(nóng)學(xué)報, 2016, 30(2):231-239.
[22] 何秀英, 王玲, 吳偉懷, 等. 水稻稻瘟病抗性基因的定位、克隆及育種應(yīng)用研究進(jìn)展[J]. 中國農(nóng)學(xué)通報, 2014, 3(6):1-12.
[23] Yang Y, Mei F, Zhang W, et al. Creation of Bt rice expressing a fusion protein of Cry1Ac and Cry1I-Like using a green tissuespecific promoter[J]. Journalof Entomology, 2014, 107(4):1674 -1679.
(責(zé)任編輯 李楠)
Cloning and Functional Analysis of Green Tissue-specific Expression Promoter of Common Wild Rice(Oryza rufipogon Griff.)
ZHAO Zhi-qiang1XUE Man-de2HUANG Ke1ZHANG Jing-wen2LONG Yan2PEI Xin-wu2YUAN Qian-hua1
(1. College of Tropical Agriculture and Forestry,Hainan University,Haikou 570228;2. Institute of Biotechnology,Chinese Academy of Agricultural Sciences,Beijing 100081)
Green tissue-specific expression promoter,which makes the exogenous gene express efficiently in green tissues of receptor crop. The green tissue-specific promoter OrGSP was cloned from Oryza rufipogon,and the fusion vector of OrGSP and GUS gene was constructed and transferred into Arabidopsis thaliana in order to identify the function of the promoter. Bioinformatics analysis shows that the length of 825 bp of OrGSP,contains the basic transcription initiation elements TATA-box and CAAT-box,and light responsive elements TCCC-motif,Sp1,G-box,I-box,GA-motif and as-2-box,etc. The GUS histochemical staining of transgenic Arabidopsis thaliana shows that OrGSP regulates the expression of GUS gene only in green tissue,and the GUS activity in leaf and stem is significantly higher than in roots. The promoter of Oryza rufipogon,OrGSP,is a green tissue-specific promoter and the results can provide new regulatory elements for crop molecular breeding.
promoter;green tissue-specific expression;common wild rice
10.13560/j.cnki.biotech.bull.1985.2017-0443
2017-01-16
國家轉(zhuǎn)基因重大專項(2016ZX08011-001),國家自然科學(xué)基金項目(31560188)
趙志強,男,碩士研究生,研究方向:植物生物技術(shù)與種質(zhì)創(chuàng)新;E-mail:zzq19900317@126.com
裴新梧,研究員,研究方向:植物基因工程;E-mail:peixw@mail.caas.net.cn
袁潛華,研究員,研究方向:作物栽培、農(nóng)業(yè)生物技術(shù);E-mail:qhyuan@163.com
猜你喜歡
品牌研究(2022年29期)2022-10-15 08:01:00
品牌研究(2022年26期)2022-09-19 05:54:46
品牌研究(2021年31期)2022-01-25 06:41:46
品牌研究(2021年36期)2022-01-21 09:29:28
品牌研究(2021年34期)2022-01-18 10:54:04
品牌研究(2021年35期)2022-01-18 08:45:18
品牌研究(2021年20期)2022-01-11 04:07:56
品牌研究(2021年27期)2022-01-04 06:41:14
品牌研究(2021年17期)2021-12-09 05:06:54
海峽姐妹(2019年3期)2019-06-18 10:37:10
