高寒干旱草原披針葉黃華植物群落土壤種子庫與地上植被的關系
2017-08-28 15:14:37張起鵬趙成章張志剛馮婉婉
生態學報 2017年14期
關鍵詞:物種
張起鵬,王 建,趙成章, 張志剛,馮婉婉, 王 珂
1 南京師范大學地理科學學院, 南京 210023 2 西北師范大學地理與環境科學學院, 蘭州 730070 3 甘肅民族師范學院, 合作 747000 4 虎臺中學, 西寧 810001
高寒干旱草原披針葉黃華植物群落土壤種子庫與地上植被的關系
張起鵬1,3,王 建1,趙成章2,*, 張志剛1,馮婉婉3, 王 珂4
1 南京師范大學地理科學學院, 南京 210023 2 西北師范大學地理與環境科學學院, 蘭州 730070 3 甘肅民族師范學院, 合作 747000 4 虎臺中學, 西寧 810001
以祁連山高寒干旱退化草地披針葉黃華(Thermopsislanceolate)群落為研究對象,采取野外調查取樣和室內分析相結合的方法,對群落土壤種子庫與地上植被的關系進行了對比研究。結果表明:地上植被物種共調查出15種,土壤種子庫檢測出植物18種,地上植被阿爾泰針茅(S.krylovii)、賴草(A.dasystachys)和扁穗冰草等可食草以及不食草披針葉黃華和阿爾泰狗哇花(H.altaicus)優勢度明顯,其它物種不明顯;土壤種子庫與地上植被物種組成的相似性程度較低;土壤種子庫與地上植被中可食草的物種數多于不食草,土壤種子庫的物種數均不小于地上植被中的物種數,地上植被中存在而未在土壤種子庫中檢測到的可食草物種有阿爾泰針茅和賴草,土壤種子庫中存在而地上植被中未被發現的物種有早熟禾(P.annua)、灰綠藜(C.glaucum)、堿韭(A.polyrhizum)、酸模(R.patientia)、獨行菜(L.apetalum)和秦艽(G.macrophylla);原生禾草物種在可食草功能群中生物量所占比例較低,毒雜草物種生物量所占比例增加顯著,而土壤種子庫的數量百分含量較低;可食草與不食草的生物量在植被梯度下的變化呈顯著負相關(r=-0.940,P=0.018),而土壤種子庫密度變化無相關性;可食草生物量變化與其土壤種子庫密度變化不相關,不食草生物量變化與其土壤種子庫密度的增加呈顯著正相關(r=0.902,P=0.036)。
干旱草原;披針葉黃華;斑塊分布;地上植被;群落相似性
草原退化是在不合理的人類活動及利用下草原植被的產量和質量下降,土壤環境惡化,使草原生態系統的生產與生態功能衰退的現象[1- 2]。近年來,祁連山北坡高寒干旱草原由于自然和人為的影響出現了嚴重的退化現象:適口性好的草本植物,特別是優良禾本科植物被牲畜大量啃食,生產力下降,許多地帶性植被建群種分布規模退縮,同時,毒雜草植物生長茂盛,形成大量以毒雜草為優勢種的斑塊狀植物群落。
披針葉黃華是我國北方草地廣泛分布的一種豆科野決明屬有毒多年生草本植物,植株新鮮狀態牲畜一般不采食,誤食后發生慢性中毒,干枯后采食不中毒[3]。目前,祁連山北坡干旱草原出現以披針葉黃華(Thermopsislanceolate)為代表的有毒植物繁茂生長的景象,草場土壤種子庫后備資源亦劣質化,牧草種子存量不容樂觀[4],草原植被生態系統出現了嚴重問題。披針葉黃華大面積生長已嚴重影響當地牧民的生產與生活,而其種群的存在不同于其它草原原生植物的環境反饋過程,尚不清楚其群落適應干擾的生存繁衍機制,無法深入理解其繁衍擴散機理。
土壤種子庫(Thesoilseedbank)是指某一特定時間,存在于土壤表面及其下的土層中的具有生活力的種子總數[5],它與地表植被有著密切的關系。作為潛在植物群落[6],它所含有的物種信息在植被的更新和恢復、群落的演替和擴散、生物多樣性的保護及生態系統演替和發展趨勢等方面都具有重要作用。許多學者從土壤種子庫的特征[4,7-10]、空間格局與異質性[11-13]、多樣性[14-15]和干擾[16-20]等方面對毒雜草植物群落進行了大量研究,而結合土壤種子庫與地上植被探究草原退化機制的研究較為薄弱[4,12]。為更全面和系統地理解土壤種子庫與地上植被的受損過程及變化模式,本文以祁連山高寒干旱草原披針葉黃華退化草地為研究對象,通過野外植被調查和室內試驗,分析了披針葉黃華植物群落土壤種子庫和地上植被功能群的關系以及和地上生物量的變化特征,揭示其群落在草原利用過程中土壤種子庫與地上植被的變化規律,以期為有效抑制披針葉黃華的生長和擴散,恢復和持續利用草原植被提供基礎。
1 研究區概況與研究方法
1.1 研究區概況
祁連山高寒干旱草原位于我國西北內陸地區的生態環境脆弱區,河西走廊與青藏高原的過渡帶,具有大陸性氣候和山地垂直氣候特征。研究區位于祁連山北坡肅南縣皇城草原,地理坐標為(37°28′—38°02′N、101°06′—102°13′E),海拔2610—2750 m,年平均氣溫1—2℃,最熱和最冷月的氣溫分別為12—15℃和-13— -11℃,≥0℃的年積溫為2450— 2600℃。年均降水量為350 mm左右,且主要集中在6—9月,年均蒸發量為1500—1800 mm,相對濕度65%。年平均日照時數達到2200—2800 h,相對無霜期80 d左右。土壤以山地栗鈣土為主,原生地帶性植被為山地草甸草原類、坡地苔草禾草組,阿爾泰針茅(S.krylovii) +扁穗冰草(A.cristatum) -雜類草型。天然牧草一般4月下旬萌發,7月上旬進入生長旺盛期,10月中旬開始枯黃,枯草期近7個月。受各種環境因子干擾,草原形成以披針葉黃華聚集分布的斑塊狀景觀,禾草種群分布規模萎縮,草原生態系統功能減弱。
1.2 研究方法
1.2.1 野外調查與種子鑒定
Thompson和Grime[5]將種子庫歸為短暫土壤種子庫和持久土壤種子庫,而持久土壤種子庫具有在承受了空間或時間上不可預測的干擾的植被中發揮繁殖能力的潛勢[21]。7、8月份采集的土樣,一般用于持久種子庫的研究,此時短暫土壤種子庫的種子基本于5、6月份萌發,而新的種子雨未降落[22]。草原植被調查與土壤種子庫的取樣在2013年8月上旬進行,根據植被特征及披針葉黃華物種生長狀況,在研究區沿南北方向選取地勢平坦,生境相對無差別的典型樣帶作為調查對象,設置50 m×50 m的調查樣地,樣地設置10個重復,重復之間間隔5 m。每個樣地沿雙對角線均勻布置9個0.5 m×0.5 m樣方進行調查,測定并分種記錄植株的蓋度、密度以及高度,然后齊地面不擾動凋落物情況下分種刈割并收集植物生物量;再在刈割后的樣方內用內徑35 mm土鉆隨機鉆取土壤種子庫樣品,深度15 cm,重復10次,將所鉆取土樣混合裝袋作為一個土壤種子庫樣品,共取土樣90份,帶回實驗室分析。
在室內,首先,將每份土樣依次過孔徑為3 mm和0.25 mm[23]的土壤篩去除枯落物、草根及雜物后進行濃縮,對留在0.25 mm篩中的土樣反復沖洗去除土壤后風干,將風干后的土樣放在雙筒顯微鏡下(4×10倍)進行種子辨別與挑選,分類并統計。對挑選出的種子用TTC[24]法進行種子生活力測定。其次,陸續將挑揀剩余物對應篩選過的土樣(90份)分別鋪在90個發芽盤內,約2—3 cm厚,在密閉玻璃溫室內接受陽光進行發芽實驗,每天定時噴灑適量水分,使土壤保持濕潤。種子萌發出苗并生長一段時間后,將其進行鑒定,統計種類與數量并將其去除。發芽盤內連續兩周無種子萌發即視為試驗結束,整個萌發試驗持續60 d。最后將物理篩選并具有生活力的植物種子與幼苗萌發所得的同種類種子數量累加。生物量是在65℃條件下烘干24 h,稱重后統計每種植物的生物量。
1.2.2 數據處理與分析
功能群是指在群落中功能相似的所有物種的集合,植物功能群往往作為一個相對統一的整體對生態因子的波動或外界干擾做出反映[25]。結合研究區干旱草原植物的利用屬性,通過訪問及直接觀察法[26],將物種劃分為牲畜可食的草類和不食草類2個功能群類型。按披針葉黃華的分蓋度將所有樣方依次歸類于5個植被退化梯度:Ⅰ(1%—10%)、Ⅱ(11%—20%)、Ⅲ(21%—30%)、Ⅳ(31%—40%)、Ⅴ(>40%)。
根據實測數據,計算群落內種群的優勢度。計算公式[27]:
優勢度=(相對密度+相對蓋度+相對高度)/3
地上植被功能群的優勢度值為功能群中不同物種優勢度之和,土壤種子庫功能群的優勢度值為功能群中不同物種的密度之和,地上生物量為物種齊地面刈割植物體烘干后每平方米的質量。
土壤種子庫與地表植被的相似性采用Sorensen(similarity coefficient)的相似性系數[28]計算。
SC=2X/(a+b)
式中,a為土壤種子庫中的物種數;b為地表植被的物種數;X為種子庫和地表植被共有的物種數。
應用Excel對實驗數據進行統計、整理,采用SPSS軟件相關分析和單因素方差分析(one-wayANOVA)對土壤種子庫及地上植被的特征與差異進行分析。
2 調查結果分析
2.1 樣方概況
本研究共涉及植被調查樣方72個以及相對應的72個種子庫樣品(表1)。
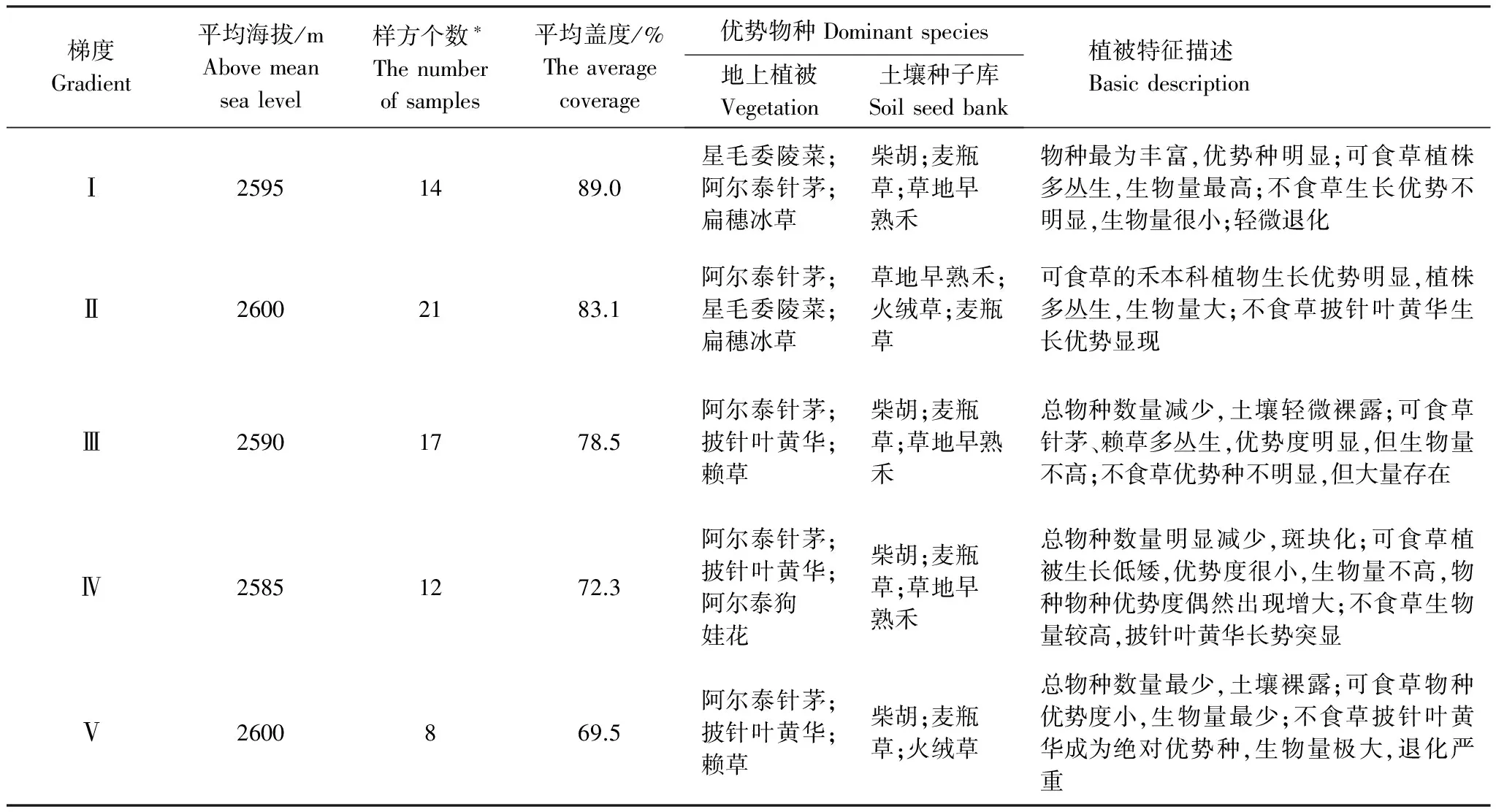
表1 研究區地上植被和土壤種子庫特征描述
*根據披針葉黃華物種存現及樣方生境確定的樣方數
總體看來,在植被梯度下,地上植被阿爾泰針茅、賴草(A.dasystachys)和扁穗冰草等禾本科可食草以及不食草的披針葉黃華和阿爾泰狗哇花(H.altaicus)優勢度明顯,可食草中的禾本科優勢度的變化不大而不食草的優勢度呈現明顯的加強;土壤種子庫中可食草早熟禾(P.annua)、扁穗冰草、小蘭花棘豆(O.glabra)和不食草柴胡(B.falcatum)、麥瓶草(S.conoidea)、火絨草(L.leontopodioides)等種子數量較多并穩定存在。不食草披針葉黃華、火絨草和狼毒(S.chamaejasme)種子出現頻次增加,其他物種的變化規律不明顯。
2.2 群落功能群的關系
2.2.1 土壤種子庫與植被功能群的相似性
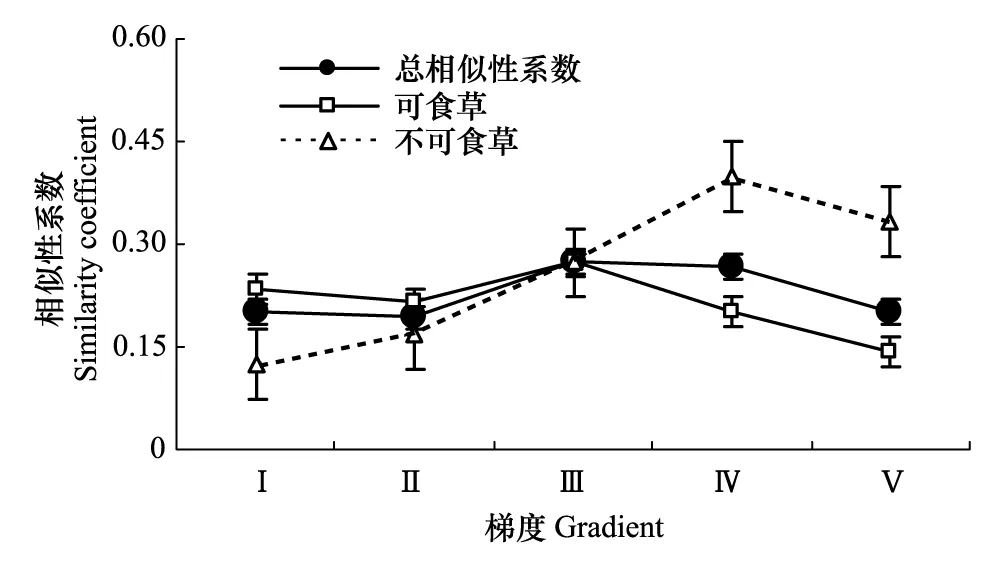
圖1 土壤種子庫與地上植被功能群的相似性 Fig.1 The similarity of plant functional groups composition of soil seed bank
由圖1可以看出,祁連山北坡高寒干旱草原土壤種子庫與植被的相似性隨著植被梯度的變化而變化,土壤種子庫和地上植被的不同功能群也表現出不同的特點,可食草土壤種子庫和地上植被相似性在梯度上減少,并在梯度Ⅲ后相似性明顯遞減。不食草相似性在梯度上出現明顯的遞增。造成這一結果的原因是地上植被與土壤種子庫共有物種存在較大差異,不食草物種在植被退化梯度上共有物種增加,可食草共有物種減少。
綜合統計表明,在植被梯度下,地上植被與土壤種子庫群落組成的相似性程度較低(0.192—0.273)。不食草相似性的增加與毒雜草的穩定出現有關而可食草相似性的降低與物種漸趨單一和物種變動較大有關。可食草地上植被與土壤種子庫的相似性系數差異不顯著(F=0.229,P=0.645),不食草地上植被與土壤種子庫的相似性系數存在顯著差異(F=6.713,P=0.018)。另外,對比分析圖中趨勢線還可發現:在Ⅰ—Ⅲ梯度下,可食草相似性系數高于不食草,說明草地的穩定性較強,而Ⅲ—Ⅴ梯度草地的穩定性相對減弱。
2.2.2 土壤種子庫與植被的功能群變化
在植被梯度下,土壤種子庫與地上植被的功能群物種組成差異很大,各物種在土壤種子庫和植被中的優勢度值變化也不盡相同。圖2為土壤種子庫與地上植被功能群在植被梯度下的物種存現關系。地上植被物種共調查出15種,土壤種子庫檢測出植物18種。總體來看,土壤種子庫中可食草的物種數多于不食草(梯度Ⅴ除外),地上植被中可食草的物種數亦多于不食草(梯度Ⅲ除外),土壤種子庫的總物種數均不小于地上植被中的物種數。
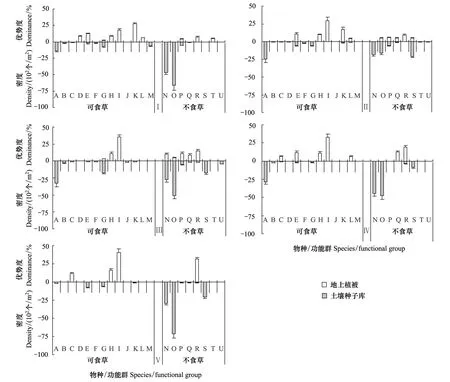
圖2 土壤種子庫與植被的功能群變化Fig.2 The change of functions in the soil seed bank and vegetation 可食草(A-M):A草地早熟禾;B灰綠藜;C鶴虱;D陰山扁蓿豆;E扁穗冰草;F堿韭;G小蘭花棘豆;H 賴草;I阿爾泰針茅;J巴天酸模;K星毛委陵菜;L冷蒿;M獨行菜;不食草(N-U):N麥瓶草;O柴胡;P 狼毒;Q阿爾泰狗哇花;R披針葉黃華;S火絨草;T甘肅馬先蒿;U秦艽
地上植被中,可食草功能群優勢度前5個物種阿爾泰針茅、賴草、星毛委陵菜(P.aclaulis)、扁穗冰草和鶴虱(L.myosotis),其值分別為31.08%、11.36%、8.98%、6.90%和3.65%,然而僅有扁穗冰草的種子在土壤種子庫中被檢測到,即使可食草物種阿爾泰針茅和賴草穩定出現在每個梯度;不食草功能群優勢度前5個物種披針葉黃華、麥瓶草、柴胡、阿爾泰狗娃花和狼毒,其值分別為16.48%、10.11%、9.65%、5.25%和4.17%,其種子則全在土壤種子庫中檢測到。
存在于地上植被中而未在土壤種子庫中檢測到的可食草物種有阿爾泰針茅和賴草,存在于土壤種子庫中但地上植被中未被發現的物種有早熟禾、灰綠藜、堿韭、酸模、獨行菜和秦艽。由此可見,研究區草原土壤種子庫中保存了較多的植物物種,這對維持當地的植物多樣性和植被的穩定起到了一定的作用。
2.3 地上生物量與土壤種子庫的關系
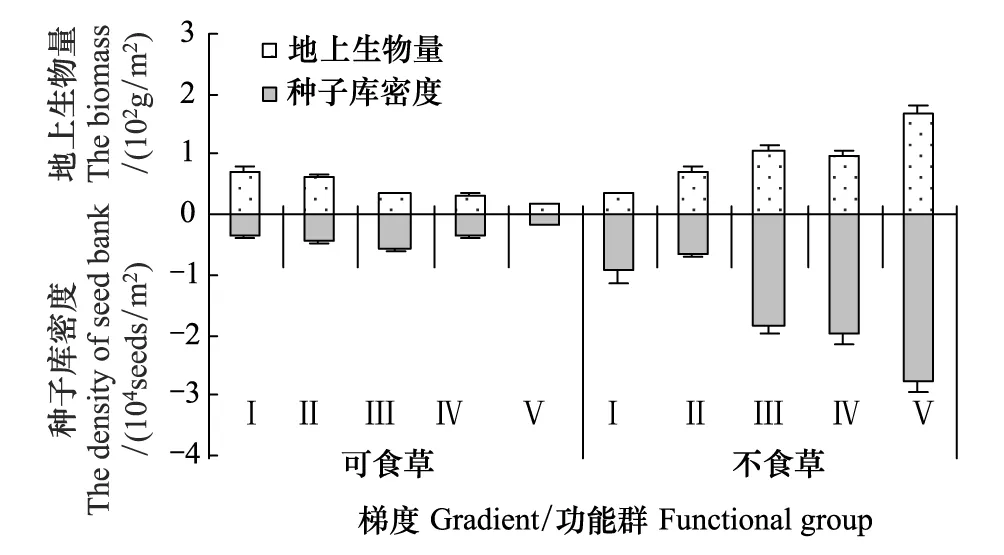
圖3 地上生物量與土壤種子庫的關系Fig.3 The relationship between biomass and the soil seed bank
在植被梯度下,不同功能群的地上植被生物量與土壤種子庫密度變化明顯(圖3):可食草物種的生物量呈遞減趨勢(y=-0.134x+ 0.842,R2=0.934),不食草物種的生物量呈遞增趨勢(y=30.417x+ 4.715,R2=0.992),兩植被功能群占地上總生物量的比值在植被梯度下趨勢亦相反;可食草土壤種子庫密度呈一個先增大后減少的單峰波動,不食草土壤種子庫密度則表現出明顯的遞增(y=0.505x+ 0.106,R2=0.862)。
相關分析表明,在植被梯度下,可食草與不食草的生物量的變化呈顯著負相關(r=-0.940,P=0.018),它們的土壤種子庫密度變化則相關性不顯著(r=-0.476,P=0.418);可食草生物量變化與其土壤種子庫密度變化相關性不顯著(r=0.318,P=0.602),不食草生物量變化與其土壤種子庫密度的增加顯著正相關(r=0.902,P=0.036)。
在植被梯度下,土壤種子庫中未檢測到的可食草物種阿爾泰針茅和賴草生物量百分含量總體較低,平均百分含量分別為7.205%和5.897%,不食草功能群優勢物種披針葉黃華和阿爾泰狗娃花生物量平均百分含量分別為52.755%和7.326%,而在土壤種子庫中檢測的數量百分含量卻很低,分別為0.586%和0.435%。另據分析可知,可食草地上植被生物量與土壤種子庫種子數量差異不顯著(F=0.272,P=0.616),不食草地上植被生物量與土壤種子庫不可食草種子數量差異亦不顯著(F=2.269,P=0.170)。
3 討論與結論
物種組成被認為是生態系統穩定性、生產力、營養動態等功能的重要決定因子[29]。一些研究認為土壤種子庫基本上反映了地表植被的組成[30-31],而更多的研究則發現土壤種子庫和地表植被的物種組成差異很大[32]。研究區高寒干旱草原植物群落功能群物種在植被梯度下變化明顯,地上植被和土壤種子庫的功能群物種組成表現出明顯的差異,可食草的物種數量減少,而土壤種子庫的緩沖作用使其中的物種和地表植被的物種具有相對較高的相似性,但隨著植被梯度的增加,植被中的可食草物種趨于單一化,土壤種子庫種子數量明顯減少,加之不食草在植被和土壤種子庫中的穩定存在,導致土壤種子庫與地表植被的相似性降低。祁連山高寒干旱草原土壤種子庫和地表植被間的物種相似性與區域環境及其所受干擾和脅迫程度有直接的關系。土壤種子庫與地上植被相似性較低的另一個原因是一些植物僅存在于地上植被中(可食草阿爾泰針茅和賴草等),而土壤種子庫中不存在或很少存在該物種的種子。
植物群落優勢種群的更替是天然草地退化的一種主要表現形式,能夠指示自然環境中的某些組成成分的變化[33]。當一種功能群的物種數量或生產力下降時,會為其它功能群物種提供更大的空間,使之占據更多的資源,隨之出現優勢度增加的趨勢。研究區放牧過程中,牲畜啃食、踐踏等行為抑制了優勢種阿爾泰針茅和賴草等可食草的繁衍和競爭優勢,這為毒雜草物種的入侵提供了機會。在一定的干擾強度下,部分優良禾本科牧草依靠營養繁殖形成聚集的群體,生物量有所增加,這補償了地上植被可食草消失物種減少的量,但不食草較強的生境適應性促使其占有更多的生長資源,這又會顯著增加其相對生物量,因而草原植被生長狀況良好,地表并未嚴重裸露。在超載放牧和氣候變化影響下,盡管毒雜草群落物種處于不斷的波動之中,但某些關鍵(功能群)物種對于整個生態系統的維持具有決定作用[34]。阿爾泰針茅、冰草和草地早熟禾等可食草對環境的適應性生長是目前維系研究區草地利用的物質基礎,但它們種子庫的存量并不樂觀。在植被梯度下,以披針葉黃華為代表的不食草優勢度和生物量的遞增,以及可食草優勢度的降低和生物量的變化,特別是禾本科物種阿爾泰針茅和賴草種子庫中種子的缺失,表明草原優良牧草競爭、更新能力受到干擾,植物群落結構上產生了較大變化,草原不食草功能群更加適應當前草原環境。
本研究對披針葉黃華群落從植物功能群角度進行研究得出功能群生物量與種子數量出現明顯的相關性,但具體到功能群中特定的種群它并沒有一致性關系,如不食草優勢種披針葉黃華和阿爾泰狗娃花生物量很大,而在土壤種子庫中檢測的種子數量百分含量卻很低。這與仲延凱等[35]對種群的生物量與種子數量的研究結果相一致。
土壤種子庫是植被潛在更新的物質基礎,在植被自然恢復過程中起著重要的作用。Bakker等[36]研究表明,雖然地上植被76%的物種在土壤種子庫中沒有出現,土壤種子庫對植被恢復仍具有重要意義,但不能作為植被恢復的主要措施;Kalamees等[37]、Roovers等[38]和Blomqvist等[39]則認為,在退化草地上利用土壤種子庫進行植被恢復的可能性極小。在進行植被恢復時,土壤種子庫與地上植被相似性較高且退化程度較低的草地,可以優先考慮利用土壤種子庫中的潛在資源進行植被恢復,而相似性較低且退化程度較高的草地,則需加以必要的人工輔助措施,才能達到較好的恢復效果。補播是植被恢復的主要措施之一,許多試驗都證實補播目標物種尤其是本地物種容易成功,可以恢復草地植被,縮短草地生態系統自然恢復進程[40]。研究區因草原利用方式及多種自然因素的影響,草地植物群落可食草物種生產力降低,規模退縮,特別是原生禾草物種(如阿爾泰針茅)的衰退以及不食草物種規模擴大,生產力增強,使草原出現了以披針葉黃華為代表的不食草逐步取代阿爾泰針茅等優良牧草的現象,形成了草原毒雜草斑塊景觀。
[1] 付華, 王彥榮, 吳彩霞, 塔拉騰. 放牧對阿拉善荒漠草地土壤性狀的影響. 中國沙漠, 2002, 22(4): 339- 343.
[2] 張自和. 無聲的危機——荒漠化與草原退化. 草業科學, 2000, 17(2): 10- 12.
[3] 內蒙古農牧學院. 草原管理學. 北京: 農業出版社, 1981: 237- 257
[4] 張起鵬, 趙成章, 王倩, 王媛,王海鵬. 高寒干旱退化草地披針葉黃華群落土壤種子庫. 干旱區地理, 2015, 38(2): 305- 311.
[5] Thompson K, Grime J P. Seasonal variation in the seed banks of herbaceous species in ten contrasting habitats. Journal of Ecology, 1979, 67(3): 893- 921.
[6] Harper J L. Population Biology of Plants. London: Academic Press, 1977: 256- 263.
[7] 柳妍妍, 胡玉昆, 王鑫, 公延明, 李凱輝. 輪葉馬先蒿群落土壤種子庫特征研究. 自然資源學報, 2011, 26(1): 48- 57.
[8] 沈有信, 劉文耀. 長久性紫莖澤蘭土壤種子庫. 植物生態學報, 2004, 28(6):768- 772.
[9] 沈有信, 趙春燕. 持久性土壤種子庫種子萌發的個體競爭能力會衰減嗎?——以紫莖澤蘭為例. 植物生態學報, 2012, 36(8): 754- 762.
[10] Tedder M J, Kirkman K P, Morris C D, Trollopec W S W, Bonyongo M C. The influence ofPechuel-Loeschealeubnitziae(wild sage) on grass sward and soil seed bank composition. African Journal of Range & Forage Science, 2012, 29(3): 101- 107.
[11] 杜晶, 趙成章, 宋清華, 史元春. 基于地統計學的退化草地狼毒種群土壤種子庫空間異質性. 生態學雜志, 2015, 34(1): 94- 99.
[12] 尚占環, 龍瑞軍, 馬玉壽, 丁路明. 青藏高原“黑土灘”次生毒雜草群落成體植株與幼苗空間異質性及相似性分析. 植物生態學報, 2008, 32(5): 1157- 1165.
[13] 趙成章, 張起鵬. 祁連山退化草地狼毒群落土壤種子庫的空間格局. 中國草地學報, 2010, 32(1): 79- 85.
[14] Kelt D A, Valone T J. Effects of grazing on the abundance and diversity of annual plants in Chihuahuan desert scrub habitat. Oecologia, 1995, 103(2): 191- 195.
[15] Hager H A, Rupert R, Quinn L D, Newman J A. EscapedMiscanthussacchariflorusreduces the richness and diversity of vegetation and the soil seed bank. Biological Invasions, 2015, 17(6): 1833- 1847.
[16] Swanton C J, Shrestha A, Knezevic S Z, Roy R C, Ball-Coelho B R. Influence of tillage type on vertical weed seedbank distribution in a sandy soil. Canadian Journal of Plant Science, 2000, 80(2): 455- 457.
[17] Jarvis J C, Moore K A. Influence of environmental factors onVallisneriaamericanaseed germination. Aquatic Botany, 2008, 88(4): 283- 294.
[18] 黨偉光, 高賢明, 王瑾芳, 李愛芳. 紫莖澤蘭入侵地區土壤種子庫特征. 生物多樣性, 2008, 16(2): 126- 132.
[19] Zhu L, Sun O J, Sang W G, Li Z Y, Ma K P. Predicting the spatial distribution of an invasive plant species (Eupatoriumadenophorum) in China. Landscape Ecology, 2007, 22(8): 1143- 1154.
[20] Gioria M, Py?ek P. The legacy of plant invasions: changes in the soil seed bank of invaded plant communities. BioScience, 2016, 66(1): 40- 53.
[21] 閆巧玲, 劉志民, 李榮平. 持久土壤種子庫研究綜述. 生態學雜志, 2005, 24(8): 948- 952.
[22] 李國旗, 李淑君, 蒙靜, 武東波. 土壤種子庫研究方法評述. 生態環境學報, 2013, 22(10): 1721- 1726.
[23] 尚占環, 龍瑞軍, 馬玉壽, 張黎敏, 施建軍, 丁玲玲. 黃河源區退化高寒草地土壤種子庫: 種子萌發的數量和動態. 應用與環境生物學報, 2006, 12(3): 313- 317.
[24] 傅家瑞. 種子生理. 北京: 科學出版社, 1985: 66- 76.
[25] 王正文, 邢福, 祝廷成, 李憲長. 松嫩平原羊草草地植物功能群組成及多樣性特征對水淹干擾的響應. 植物生態學報, 2002, 26(6): 708- 716.
[26] 楊澤新, 蔡維湘. 灌叢草地放牧山羊的牧草適口性與嗜食性及山羊采食率研究. 草業科學, 1995, 12(2): 20- 21, 24- 24.
[27] 林麗. 石羊河上游退化草地植物功能群特征分析[D]. 蘭州: 甘肅農業大學, 2007.
[28] Guo Q F, Rundel P W, Goodall D W. Structure of desert seed banks: comparisons across four North American desert sites. Journal of Arid Environments, 1999, 42(1): 1- 14.
[29] Bengtsson J. Which species? What kind of diversity? Which ecosystem function? Some problems in studies of relations between biodiversity and ecosystem function. Applied Soil Ecology, 1998, 10(3): 191- 199.
[30] Yuan L Y, Liu G H, Li W, Li E H. Seed bank variation along a water depth gradient in a subtropical lakeshore marsh, Longgan Lake, China. Plant Ecology, 2007, 189(1): 127- 137.
[31] Van der Valk A G, Davis C B. The role of seed banks in the vegetation dynamics of prairie glacial marshes. Ecology, 1978, 59(2): 322- 335.
[32] Smith L M, Kadlec J A. Seed banks and their role during drawdown of a North American marsh. Journal of Applied Ecology, 1983, 20(2): 673- 684.
[33] 劉成林, 樊任華, 武建軍, 閆峰. 錫林郭勒草原植被生長對降水響應的滯后性研究. 干旱區地理, 2009, 32(4): 512- 518.
[34] Walker B. Conserving biological diversity through ecosystem resilience. Conservation Biology, 1995, 9(4): 747- 752.
[35] 仲延凱, 包青海, 孫維, 張海燕. 割草干擾對典型草原土壤種子庫種子數量與組成的影響. Ⅳ群落生物量的組成與種子數量組成的比較. 內蒙古大學學報: 自然科學版, 2001, 32(5): 551- 556.
[36] Bakker C, De Graaf H F, Ernst W H O, van Bodegom P M. Does the seed bank contribute to the restoration of species-rich vegetation in wet dune slacks?. Applied Vegetation Science, 2005, 8(1): 39- 48.
[37] Kalamees R, Zobel M. Soil seed bank composition in different successional stages of a species rich wooded meadow in Laelatu, western Estonia. Acta Oecologica, 1998, 19(2): 175- 180.
[38] Roovers P, Bossuyt B, Igodt B, Hermy M. May seed banks contribute to vegetation restoration on paths in temperate deciduous forest?. Plant Ecology, 2006, 187(1): 25- 38.
[39] Blomqvist M M, Bekker R M, Vos P. Restoration of ditch bank plant species richness: the potential of the soil seed bank. Applied Vegetation Science, 2003, 6(2): 179- 188.
[40] Le Floc′H E, Neffati M, Chaieb M, Floret C, Pontanier R. Rehabilitation experiment at Menzel Habib, Southern Tunisia. Arid Soil Research and Rehabilitation, 1999, 13(4): 357- 368.
Relationships between the soil seed bank and above-ground vegetation of aThermopsislanceolatacommunity in degraded alpine arid grasslands
ZHANG Qipeng1,3, WANG Jian1,ZHAO Chengzhang2,*, ZHANG Zhigang1, FENG Wanwan3, WANG Ke4
1SchoolofGeographicScience,NanjingNormalUniversity,Nanjing210023,China2CollegeofGeographyandEnvironmentalScience,NorthwestNormalUniversity,Lanzhou730070,China3GansuNormalUniversityforNationalities;Hezuo747000,China4HuTaiMiddleSchoolofXining,Xining810001,China
The main objective of this study was to address questions with regard to how the soil seed banks and vegetation change along a habitat gradient. In this article, we chose theThermopsislanceolatacommunity in degraded alpine arid grasslands as our research subject and determined the relationship between the alpine arid grassland soil seed bank and above-ground vegetation in the Qilian Mountains using field investigation and laboratory analysis. Fifteen species were present in the above-ground vegetation and 18 species were present in the soil seed bank. Dominant above-ground vegetation species included edible grasses such asStipakrylovii,AneurolepidiumdasystachysandAgropyroncristatum, and non-edible grasses such asThermopsisLanceolataandHeteropappusaltaicus; however, no other species was observed to exhibit any distinct dominance. Moreover, the degree of similarity between the above-ground vegetation and the soil seed bank was distinctive. The Sorenson similarity index was low between the above-ground vegetation and the soil seed bank along the vegetation gradient. The Sorenson similarity index between the soil seed bank and observed vegetation functional groups exhibited varying characteristics. The number of edible grasses was greater than that of non-edible grasses in the soil seed bank (except V gradient). The number of species of vegetation was also greater than that of non-edible grasses (except III gradient). The number of species in the soil seed bank was not lower than that of the above-ground vegetation species.S.kryloviiandA.dasystachyswere founded in the above-ground vegetation but not in the soil seed bank, whereasPoaannua,Chenopodiumglaucum,Alliumpolyrhizum,Rumexpatientia,Lepidiumapetalum, andGentianamacrophyllawere found in the soil seed bank but not in the vegetation. The proportion of biomass attributable to gramineous plants was low for edible grasses, whereas the biomass of poisonous weeds was significantly greater in the vegetation gradient than in the soil seed bank. The plant population biomass had no unanimous relation with seed amount. Edible grass and non-edible grass were significantly negatively correlated (r=-0.940,P=0.018) along the vegetation gradient, but changes in the soil seed bank density were not correlated. The relationship between the biomass of edible grass and the soil seed bank density was uncorrelated. The biomass of non-edible grass and the soil seed bank density were significantly correlated along the vegetation gradient (r=0.902,P=0.036). Our research indicated that the seed bank could retain and restore several species in the process of grassland degradation, and these species could be valuable for vegetation succession and grassland restoration.
arid grasslands;Thermopsislanceolata; patch distribution; vegetation;similarity of community
國家自然科學基金項目(91125014)
2016- 04- 10; 網絡出版日期:2017- 03- 02
10.5846/stxb201604100654
*通訊作者Corresponding author.E-mail: zhaocz@nwnu.edu.cn
張起鵬,王建,趙成章, 張志剛,馮婉婉, 王珂.高寒干旱草原披針葉黃華植物群落土壤種子庫與地上植被的關系.生態學報,2017,37(14):4619- 4626.
Zhang Q P, Wang J, Zhao C Z, Zhang Z G, Feng W W, Wang K.Relationships between the soil seed bank and above-ground vegetation of aThermopsislanceolatacommunity in degraded alpine arid grasslands.Acta Ecologica Sinica,2017,37(14):4619- 4626.
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2024年12期)2024-12-02 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
中學生博覽(2022年7期)2022-06-21 21:48:14
大科技·百科新說(2021年8期)2021-11-03 10:55:16
學苑創造·A版(2021年5期)2021-06-28 19:51:42
少兒美術(快樂歷史地理)(2020年9期)2020-03-19 05:10:56
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
