大青溝自然保護區主要森林群落優勢種的葉性狀
2017-08-28 15:14:37劉貴峰劉玉平達福白乙拉程偉燕高學磊姜麗麗
生態學報 2017年14期
劉貴峰,劉玉平,達福白乙拉,程偉燕,高學磊,姜麗麗
1 內蒙古民族大學農學院, 通遼 0280002 內蒙古大青溝國家級自然保護區管理局, 甘旗卡 0280003 中國科學院青藏高原研究所,高寒生態與生物多樣性重點實驗室,北京 100101
大青溝自然保護區主要森林群落優勢種的葉性狀
劉貴峰1,*,劉玉平1,達福白乙拉2,程偉燕1,高學磊1,姜麗麗3
1 內蒙古民族大學農學院, 通遼 0280002 內蒙古大青溝國家級自然保護區管理局, 甘旗卡 0280003 中國科學院青藏高原研究所,高寒生態與生物多樣性重點實驗室,北京 100101
植物功能性狀是近年來生態學研究的熱點。其中葉功能性狀與植株生物量和植物對資源的獲得、利用及利用效率的關系最為密切。大青溝森林植物群落分布于科爾沁沙地,生境條件非常特殊,在沙溝里存在著一片茂密的森林,與周圍浩瀚無垠的沙坨景觀形成極為鮮明的對照。從溝底到溝頂,雖然海拔高度僅相差六、七十米,但由于距離溝底水源不同,土壤條件差異大,形成了不同的森林植物群落類型。大果榆群落、蒙古櫟群落和水曲柳群落分別分布在大青溝自然保護區的溝頂、溝中和溝底。為了對大青溝自然保護區,不同環境梯度下的森林植物群落葉片功能性狀進行研究,以大青溝自然保護區大果榆、蒙古櫟、水曲柳3種主要森林群落為研究對象,分別測定不同群落優勢種的葉厚度、比葉面積、葉干物質含量、葉大小和葉干重等5 項葉功能性狀,研究葉功能性狀之間的關系,并對不同生長型、不同群落葉功能性狀進行比較。相關分析結果表明,葉厚度與比葉面積呈極顯著負相關,與葉大小和葉干重呈極顯著正相關;比葉面積與葉干物質含量、葉干重呈極顯著負相關,與葉大小呈顯著正相關;葉干物質含量與葉大小呈極顯著負相關,與葉干重呈極顯著正相關;葉大小與葉干重呈極顯著正相關。不同生長型植物葉片性狀的分析表明,草本植物的葉干物質含量比喬木和灌木低,而其比葉面積高于喬木和灌木;對不同群落葉功能性狀進行比較發現,大果榆群落和蒙古櫟群落喬木、灌木葉厚度與干物質含量顯著高于水曲柳群落的葉厚度與干物質含量,二者比葉面積顯著低于水曲柳群落。大果榆群落和蒙古櫟群落從葉功能性狀的角度,它們具有較高的葉干物質含量和較低的比葉面積,體現出適應干旱生境葉片的特征。水曲柳群落呈現出低葉干物質含量、高比葉面積的特征,體現出適應濕潤、土壤水分較好生境的特征。不同群落通過調節自身的物種組成,形成不同的功能性狀組合來適應環境。
大青溝自然保護區;森林群落;優勢種;葉性狀;生長型
植物功能性狀是在形態、生理、物候等方面表征植物的生態策略,決定植物如何應對環境因素,并且進一步影響其他營養級和生態系統特性的植物性狀[1],如生長型、最大高度、木材密度、比葉面積、葉干物質含量、光合能力、固氮能力、葉片氮磷含量、果實類型、種子大小和散布方式等植物形態、生理和物候等特征[2]。它反映了植物在表征生態系統功能方面的生態指示作用,強調其與生態系統過程和功能的關系[3]。植物的光合作用是陸地生態系統初級生產力的主要來源,是大多數生態系統中食物鏈的基礎。同時,植物利用光合作用中獲得的能量從環境中吸收營養物質,并支撐整個植株的新陳代謝。而葉片是植物光合作用的主要場所,植物葉的功能性狀直接影響光合作用的速率和持久性,因而對植物個體的生存與繁殖極為重要[4- 6]。植物的葉性狀能夠反映植物適應環境的策略,同時深刻影響著生態系統的生產力水平、能量流動和物質循環。對植物葉性狀、葉性狀之間的關系的研究一直是植物生態學的一個重要領域[7- 9]。葉功能性狀分為結構型性狀和功能型性狀。結構型性狀指葉片的生物化學結構特征,包括葉面積、比葉面積、葉厚度、葉干物質含量等,此性狀相對較穩定并可以很好地反映植物為獲得最大化碳收獲而采取的生存適應策略。功能型性狀主要包括凈光合速率、呼吸速率和氣孔導度等,此性狀體現葉片的生長代謝指標,并隨時間和空間的變異相對較大,因此難以深入解釋植物在長期進化過程中的適應機理[10- 12]。
大青溝自然保護區是國家級自然保護區,是我國乃至世界罕見的沙地溝壑植物生態系統,是科爾沁沙地中僅存的一塊原始森林植物群落。這條小小沙溝里存在的一片茂密森林與周圍浩瀚無垠的沙坨景觀形成鮮明對照,溝內溪水常年不凍,生境條件非常特殊。大青溝森林植物群落是經過長期的環境變遷殘留下來的森林生態系統類型,對于正確認識該地區的環境、植被及氣候的演變過程有重要意義[13]。在大青溝自然保護區,從溝底到溝頂,雖然海拔高度僅相差六、七十米,但由于距離溝底水源不同,土壤條件差異大,形成了不同的群落類型。在大青溝自然保護區的溝頂、溝中和溝底,分別分布著大果榆(UlmusmacrocarpaHance.)群落、蒙古櫟(QuercusmongolicaFisch.)群落和水曲柳(FraxinusmandshuricaRupr.)群落。目前關于葉功能性狀的研究很多,研究目標通常是在宏觀尺度上揭示植物對環境的適應機制[14-16]。但在某些特殊的生境中,較小的空間尺度下也存在較大的環境梯度差異,例如沙地溝壑生境。然而目前這種特殊生境的生態學研究較少,小尺度范圍內的群落結構變化及其內在機制還缺乏系統研究。為了對大青溝自然保護區,不同環境梯度下的森林植物群落葉片功能性狀進行研究,本文選取大青溝這3 種具有代表性的森林群落作為研究對象,通過測定優勢種葉厚度、比葉面積、葉干物質含量、葉大小和葉干重5 項葉功能性狀,探討大青溝自然保護區不同森林群落優勢種葉功能性狀之間的關系;不同生長型葉性狀間的差異;比較不同群落葉功能性狀的差異。本文在相對較小的尺度,從群落水平開展研究,為大青溝自然保護區的資源保護利用、生態工程建設及生態環境的保護提供依據。
1 研究區概況
大青溝自然保護區位于內蒙古自治區東部通遼市科爾沁左翼后旗甘旗卡西南24 km處,地理位置為122°13′—122°15′E,42°45′—42°48′N,總面積達8183 hm2。在保護區內分布有大、小青溝兩大溝壑,大青溝長20 km,溝深40—50 m,溝寬平均250 m,溝坡平均36°。小青溝長10 km,溝深50—70 m,溝寬平均200—300 m,坡度平均28°。兩溝內水流匯合后流入柳河,溝內水面寬2—4 m,水深不超過1 m。海拔高度溝上225—253 m,溝下173—200 m,為該區海拔最低的地方。溝外為一望無際,此起彼伏的沙丘,沙丘一般高度在10 m 以下[17- 18]。
2 研究方法
2.1 野外調查和固定樣地設置
2013年6—9月進行野外群落學調查,在對大青溝自然保護區進行全面踏查的基礎上,每種群落類型設置5 個20 m×20 m固定樣地,共設置15 個20 m×20 m樣地。將每個20 m×20 m固定樣地劃分為16 個5 m×5 m的小樣方,在其中進行木本植物的調查,分別測量喬木坐標、胸徑、樹高、冠幅、枝下高,測量灌木高度、基徑、冠幅、蓋度,草本植物的株數、高度和蓋度。在上述調查完成后還測定各樣地的坡向、坡度、坡位、海拔高度等環境因子。溫濕度數據使用HOBOProTemp/RH dataloggers U23-001采集,每個400m2樣地內各放置1個記錄儀,每隔1h自動測定并記錄空氣溫度和相對濕度。生長季指的是5—9月。研究地點概況見表1。
2.2 葉片采集
計算每個樣地喬木層、灌木層、草本層各物種的重要值,選取重要值占優勢(大于0.1的)的物種進行葉片采集。確定優勢種后,每個樣地中,每一優勢種選擇5株或10株(喬木、灌木為5株,草本為10株)生長成熟、長勢良好的個體,采集完全展開、沒有病蟲害且未被遮光(一些陰生植物除外)的葉片。使用5米桿可伸縮高枝剪并請當地擅長爬樹的人員協助,從每個個體冠層陽面采集至少3個帶全部現存葉片的末端小枝,包括至少20片健康的成熟葉片或復葉的小葉,將采集的枝葉裝入自封袋帶回駐地,當天掃描葉片。
2.3 葉性狀的測定
測定葉厚度(cm)、比葉面積(m2/kg)、葉干物質含量(mg/g)、葉大小(cm2)和葉干重(g)等5 項指標。
2.3.1 葉厚度的測定方法
將一株樹上的5片葉子疊放在一起,沿著主脈的方向均勻選3個點,選用精度為0.02 mm的游標卡尺,測量葉片主脈一側約0.25 cm處的厚度,測得的3個厚度值做平均再除以5即該組葉片的平均厚度,每株樹測5組,其平均值即該株樹葉片的平均厚度,每種樹測5株,其平均值即該種樹的葉片平均厚度[11,19]。
2.3.2 葉大小、葉干重和比葉面積的測定方法
用HP G3110掃描儀掃描采集的優勢樹種的葉面積,每株樹掃描5組,用Note葉面積計算軟件計算出每組的葉面積,除以相應的葉片個數,得到的平均值就是葉大小(cm2);將掃描后的葉片放入信封中,帶回實驗室,將葉片放入80℃烘箱內烘干24 h后取出稱重,測定干重,葉干重=干重/葉片個數;SLA=葉片面積(m2)/葉片干重()[19- 20]。
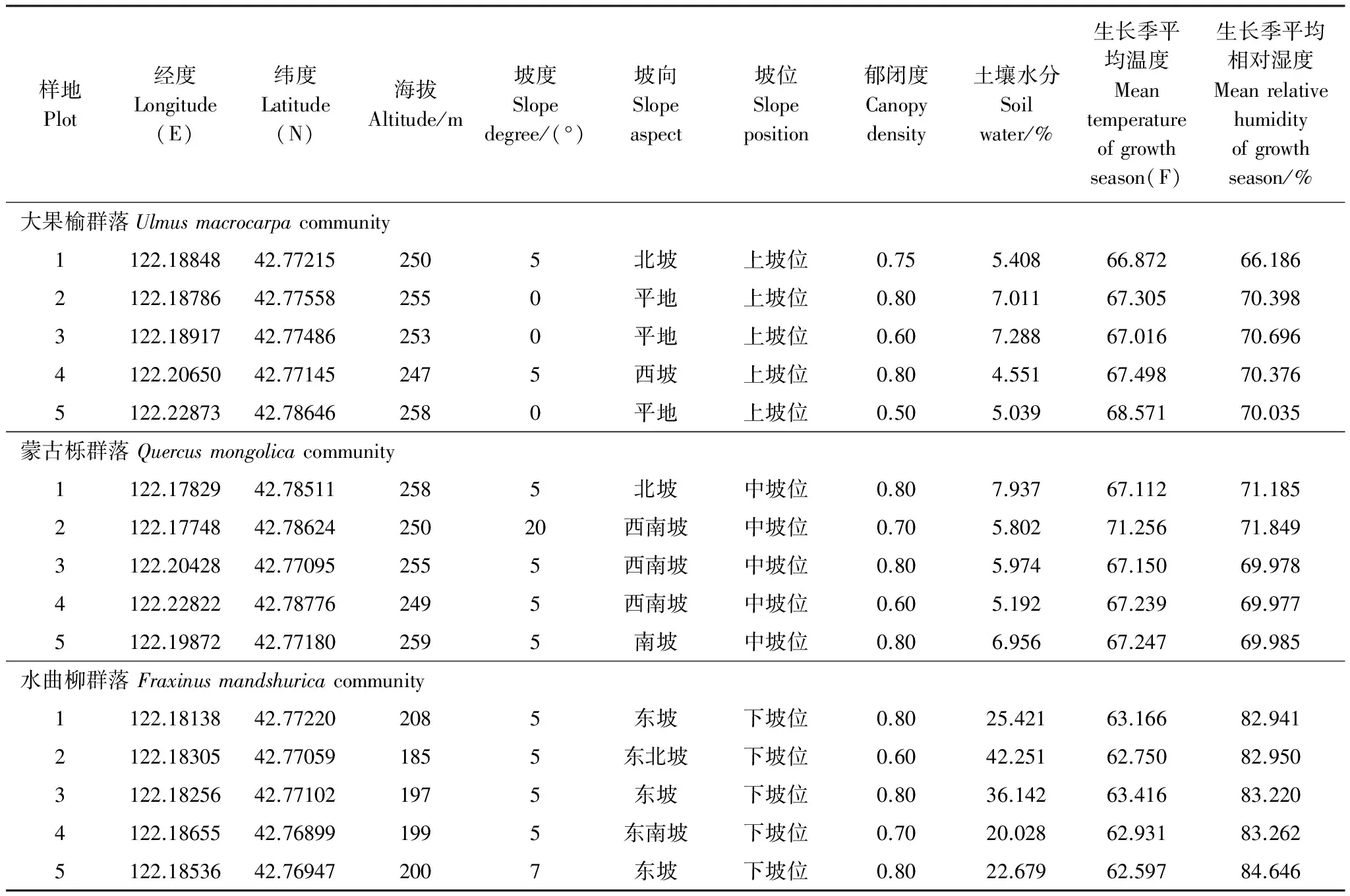
表1 研究地點概況
2.3.3 葉干物質含量的測定方法
將葉片放在清水中置于黑暗處浸泡 12 h后,取出后迅速用濾紙吸干葉片表面的水分,在1/10000的電子天平上稱重(飽和鮮重)。將葉片放入80℃烘箱內烘干24 h后取出稱重(干重)。LDMC=葉片干重(mg)/葉片飽和鮮重(g)[21]。
2.4 數據處理方法
2.4.1 重要值的計算
選取重要值大于0.1的物種進行葉片采集和測定。喬木重要值=(相對密度+相對顯著度+相對頻度)/300;灌木重要值=(相對密度+相對蓋度+相對頻度)/300;草本重要值=(相對高度+相對蓋度+相對頻度)/300[19- 20]。各樣地優勢種見附錄1。
2.4.2 Pearson相關分析
首先對各樣地優勢樹種的葉性狀進行測定,測定的葉性狀值經過匯總處理用SPPS 19.0進行葉功能性狀間的Pearson相關檢驗。
2.4.3 不同生長型葉性狀的比較
將所有優勢種分為喬木、灌木、草本3種生長型,進行葉性狀的比較。采用One-way ANOVA分析,并用Duncan法對各參數平均數進行顯著性檢驗,用Excel 2010完成作圖。
2.4.4 群落間葉功能性狀的比較
將每個群落,測定的葉功能性狀值,分為不同生長型,結合相應的重要值進行加權平均,然后把同一群落、不同生長型葉功能性狀加權值進行平均,從而對不同群落、不同生長型葉性狀值進行比較[19- 20]。采用One-way ANOVA分析,并用Duncan法對各參數平均數進行顯著性檢驗,用Excel 2010完成作圖。
3 結果與分析
3.1 葉功能性狀間的相關分析
從表2可以看出,大青溝自然保護區所有優勢種葉性狀之間的關系:葉厚度與比葉面積呈極顯著負相關,與葉大小和葉干重呈極顯著正相關;比葉面積與葉干物質含量、葉干重呈極顯著負相關,與葉大小呈顯著正相關;葉干物質含量與葉大小呈極顯著負相關,與葉干重呈極顯著正相關;葉大小與葉干重呈極顯著正相關。也就是說,葉子越厚,葉大小、葉干重也越大;比葉面積越大,葉干物質含量和葉干重越小;葉大小越大,葉干重也越大。

表2 大青溝自然保護區所有優勢種葉性狀Pearson相關分析
**P<0.01; *P<0.05;N=745

圖1 不同生長型葉功能性狀Fig.1 Leaf functional traits in different growth forms不同小寫字母代表不同生長型葉性狀之間具有顯著差異(P<0.05); 圖中數據為平均值±標準誤差
3.2 不同生長型植物葉片性狀的分析
從圖1可以看出,葉厚度灌木和草本之間沒有顯著差異,二者與喬木之間具有顯著差異,喬木葉厚度最大;比葉面積和葉干物質含量變化趨勢正好相反,喬木和灌木的比葉面積、葉干物質含量均無顯著差異,二者均與草本的比葉面積和干物質含量具有顯著差異,草本的比葉面積最大,葉干物質含量最小;喬木和草本的葉大小無顯著差異,二者與灌木葉大小具有顯著差異,灌木葉大小最小;喬木、灌木、草本葉干重均具有顯著差異,呈現出喬木>灌木>草本的趨勢。
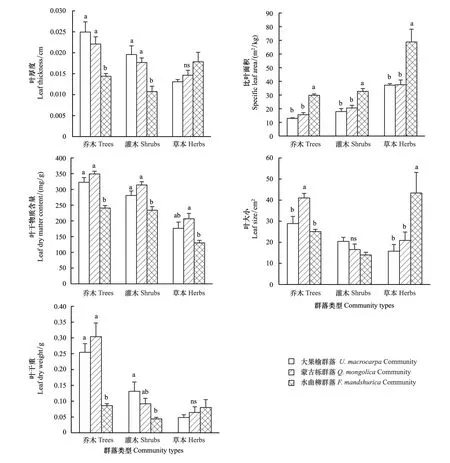
圖2 不同群落葉功能性狀Fig.2 Leaf functional traits in different communities不同小寫字母代表不同群落不同生長型葉性狀之間具有顯著差異(P<0.05); ns表示不同群落不同生長型葉性狀之間無顯著差異; 圖中數據為平均值±標準誤差
3.3 不同群落葉功能性狀的比較
從圖2可以看出,各群落喬木,葉厚度大果榆群落和蒙古櫟群落無顯著差異(P>0.05),二者均與水曲柳群落具有顯著差異,水曲柳群落葉厚度最小;比葉面積大果榆群落和蒙古櫟群落無顯著差異(P>0.05),二者均與水曲柳群落具有顯著差異,水曲柳群落比葉面積最大;葉干物質含量蒙古櫟群落和大果榆群落沒有顯著差異(P>0.05),二者均與水曲柳群落具有顯著差異,水曲柳群落葉干物質含量最小;葉大小大果榆群落和水曲柳群落無顯著差異,二者均與蒙古櫟群落具有顯著差異,且蒙古櫟群落葉大小最大;葉干重蒙古櫟群落和大果榆群落無顯著差異(P>0.05),二者均與水曲柳群落具有顯著差異,且水曲柳群落葉干重最小。各群落之間的灌木,葉大小無顯著差異(P>0.05),葉厚度、比葉面積和干物質含量變化趨勢均與喬木一樣;葉干重只有大果榆群落和水曲柳群落有顯著差異(P<0.05),其他均無顯著差異,水曲柳群落葉干重最小。各群落的草本,葉厚度和葉干重均無顯著差異(P>0.05),比葉面積變化趨勢與喬木和灌木一樣;葉干物質含量只有蒙古櫟群落和水曲柳群落有顯著差異,其他均無顯著差異(P>0.05);葉大小水曲柳群落最大,且與蒙古櫟和大果榆群落具有顯著差異。
4 結論與討論
4.1 葉功能性狀間的關系
許多研究表明,葉干物質含量與比葉面積之間呈負相關關系[22- 24],本研究結果與其一致。葉干物質含量的增加,即比葉面積的減小,使葉片內部水分向葉片表面擴散的距離或阻力增大,降低植物內部水分散失[22]。葉片厚度通常被作為一個非常有價值的特性,它在植物生長方面扮演著重要角色,其與植物獲取資源、利用資源的策略都極其相關[25- 27]。Reich等以及Witkowski等研究發現養分貧瘠環境中,植物葉片較厚[28- 29]。Reich等認為比葉面積較低的植物葉常形成厚度較大而面積較小的葉片[28]。李宏偉等[20]對黃土高原子午嶺不同森林群落葉功能性狀研究表明,LT與SLA呈極顯著負相關。楊建軍等[30]對云南地區高山栲(CastanopsisdelavayiFranch)葉性狀研究表明,葉厚度與比葉面積呈極顯著負相關。宋光等[31]對陜北黃土高原刺槐植物功能性狀研究表明,刺槐植物功能性狀相互之間都有一定的相關關系,比葉面積(SLA)與葉厚度(LT)呈極顯著負相關關系。Riva等[23]研究表明,比葉面積與葉厚度呈極顯著負相關。這些研究與本文葉厚度與比葉面積之間呈負相關的關系一致。
4.2 不同生長型植物葉片性狀的分析
本研究表明,草本植物的葉干物質含量比喬木和灌木低,而其比葉面積高于喬木和灌木(圖1,圖2)。寶樂等[19]對東靈山地區不同森林群落葉功能性狀研究表明,草本植物的葉干物質含量普遍比喬木和灌木低,而其比葉面積普遍高于喬木和灌木。施宇等[32]對黃土丘陵區延河流域植物功能性狀研究表明,不同生長型功能性狀在環境梯度下存在差異,表現為草本植物相對木本植物有更大的比葉面積。呂金枝等[33]對山西霍山30種不同功能型植物葉性特征的研究表明,草本植物的比葉面積最大,灌木次之,喬木最小。Chen[34]等對大青溝保護區147物種的SLA、N、P、C/N、C/P、N/P進行研究表明,草本植物比葉面積高于灌木和喬木。Wright等[35]的研究表明,平均來說,與草本相比,喬木和灌木有更低的比葉面積。本文研究結果與以上研究結果一致。也就是說,不同生長型植物,對環境的適應能力明顯不同,這是由于不同生長型物種對葉片氮的分配策略不同。喬木和灌木往往分配較多的生物量和氮于細胞壁,以增強葉片韌性,同時積累較多的光合產物為越冬和第2年的生長做準備,因而其比葉面積,分配到光合器中的氮較少,以致其光合能力較低。而草本植物分配較多的有機氮于類囊體和RuBP羧化酶中使其具有較高的光合能力,植物生長加快,尤其是地上部分生長迅速,葉片較薄,葉面積增大,SLA較高[19,36]。
4.3 不同群落間葉功能性狀的比較
葉片干物質含量和比葉面積是植物功能生態學研究中最常用的性狀,它們在植物葉片功能中發揮著重要作用,能反映植物對資源獲取和利用情況[2],研究指出葉片干物質含量高,比葉面積小的植物資源獲取能力強,更能在逆境中占優勢[22,37]。Hodgson 等[38]認為比葉面積是最適合研究的植物功能性狀,它能代表土壤的肥力,比葉面積是葉干物質含量與葉厚度綜合作用的結果。
大果榆群落和蒙古櫟群落分別分布在溝頂、溝中,距離水源相對較遠,具有葉厚度較大、比葉面積較小、葉干物質含量較大的葉片功能性狀。水曲柳群落分布在溝底,距離水源較近,具有葉厚度較小、比葉面積較大、葉干物質含量較小的葉片功能性狀。以水曲柳為主的群落在長期的自然演替過程中已達到與生境條件相互適應的動態平衡過程,它僅僅是一種小生境條件作用下的隱域性穩定植被系統。以蒙古櫟為主的群落在當地處于穩定狀態,并與當地的大氣候條件相適應,以大果榆為主的群落則處于相對不穩定的狀態中[39]。蒙古櫟群落和大果榆群落從葉功能性狀的角度,它們具有較高的葉干物質含量和較低的比葉面積,體現出適應干旱生境葉片的特征。水曲柳群落呈現出低葉干物質含量、高比葉面積的特征,體現出適應濕潤、土壤水分較好生境的特征。大青溝自然保護區,3 種主要森林植物群落,通過調節自身的物種組成,形成不同的功能性狀組合來適應環境。Riva等[23]的研究表明,群落水平的植物功能性狀之間的關系要強于物種水平的,支持了主要物種更適合當地環境條件,形成植被功能群的假設。
[1] Pérez-Harguindeguy N, Díaz S, Garnier E, Lavorel S, Poorter H, Jaureguiberry P, Bret-Harte M S, Cornwell W K, Craine J M, Gurvich D E, Urcelay C, Veneklaas E J, Reich P B, Poorter L, Wright I J, Ray P, Enrico L, Pausas J G, de Vos A C, Buchmann N, Funes G, Quétier F, Hodgson J G, Thompson K, Morgan H D, ter Steege H, van der Heijden M G A, Sack L, Blonder B, Poschlod P, Vaieretti M V, Conti G, Staver A C, Aquino S, Cornelissen J H C. New handbook for standardised measurement of plant functional traits worldwide. Australian Journal of Botany, 2013, 61(3): 167- 234.
[2] Cornelissen J H C, Lavorel S, Garnier E, Díaz S, Buchman N, Gurvich D E, Reich P B, ter Steege H, Morgan H D, van der Heijden M G A, Pausas J G, Poorter H. A handbook of protocols for standardised and easy measurement of plant functional traits worldwide. Australian Journal of Botany, 2003, 51(4): 335- 380.
[3] 孟婷婷, 倪健, 王國宏. 植物功能性狀與環境和生態系統功能. 植物生態學報, 2007, 31(1): 150- 165.
[4] Kikuzawa K. A cost-benefit analysis of leaf habit and leaf longevity of trees and their geographical pattern. The American Naturalist, 1991, 138(5): 1250- 1263.
[5] Li L, Mccormack M L, Ma C G, Kong D L, Zhang Q, Chen X Y, Zeng H, Niinemets ü, Guo D L. Leaf economics and hydraulic traits are decoupled in five species-rich tropical-subtropical forests. Ecology Letters, 2015, 18(9): 899- 906.
[6] Reich P B, Walters M B, Ellsworth D S. From tropics to tundra: global convergence in plant functioning. Proceedings of the National Academy of Sciences of the United States of America, 1997, 94(25): 13730- 13734.
[7] Díaz S, Kattge J, Cornelissen J H C, Wright I J, Lavorel S, Dray S, Reu B, Kleyer M, Wirth C, Prentice I C, Garnier E, B?nisch G, Westoby M, Poorter H, Reich P B, Moles A T, Dickie J, Gillison A N, Zanne A E, Chave J, Wright S J, Sheremet′ev S N, Jactel H, Baraloto C, Cerabolini B, Pierce S, Shipley B, Kirkup D, Casanoves F, Joswig J S, Günther A, Falczuk V, Rüger N, Mahecha M D, Gorné L D. The global spectrum of plant form and function. Nature, 2016, 529(7585): 167- 171.
[8] Zhang J L, Poorter L, Cao K F. Productive leaf functional traits of Chinese savanna species. Plant Ecology, 2012, 213(9): 1449- 1460.
[9] Blonder B, Vasseur F, Violle C, Shipley B, Enquist B J, Vile D. Testing models for the leaf economics spectrum with leaf and whole-plant traits inArabidopsisthaliana. AoB Plants, 2015, 7: plv049.
[10] 張林, 羅天祥. 植物葉壽命及其相關葉性狀的生態學研究進展. 植物生態學報, 2004, 28(6): 844- 852.
[11] 劉金環, 曾德慧, Lee D K. 科爾沁沙地東南部地區主要植物葉片性狀及其相互關系. 生態學雜志, 2006, 25(8): 921- 925.
[12] 黃文娟, 李志軍, 楊趙平, 白冠章. 胡楊異形葉結構型性狀及其相互關系. 生態學報, 2010, 30(17): 4636- 4642.
[13] 趙學勇, 常學禮, 張志堅, 李玉山, 韓鳳樓. 大青溝國家自然保護區土地利用與覆蓋變化研究. 中國沙漠, 1999, 19(S1): 132- 136.
[14] Niinemets ü, Keenan T F, Hallik L. A worldwide analysis of within-canopy variations in leaf structural, chemical and physiological traits across plant functional types. New Phytologist, 2015, 205(3): 973- 993.
[15] Meng T T, Wang H, Harrison S P, Prentice I C, Ni J, Wang G. Responses of leaf traits to climatic gradients: adaptive variation versus compositional shifts. Biogeosciences Discussions, 2015, 12(18): 7093- 7124.
[16] Tian M, Yu G R, He N P, Hou J H. Leaf morphological and anatomical traits from tropical to temperate coniferous forests: mechanisms and influencing factors. Scientific Reports, 2016, 6: 19703.
[17] 鄭元潤. 大青溝森林植物群落物種多樣性研究. 生物多樣性, 1998, 6(3): 191- 196.
[18] 劉貴峰, 程偉燕, 劉玉平, 張淑娟. 大青溝自然保護區主要森林群落優勢種的種群結構與動態. 西北植物學報, 2014, 34(4): 843- 850.
[19] 寶樂, 劉艷紅. 東靈山地區不同森林群落葉功能性狀比較. 生態學報, 2009, 29(7): 3692- 3703.
[20] 李宏偉, 王孝安, 郭華, 王世雄, 夏菲. 黃土高原子午嶺不同森林群落葉功能性狀. 生態學雜志, 2012, 31(3): 544- 550.
[21] 周欣, 左小安, 趙學勇, 劉川, 呂朋. 科爾沁沙地中南部34種植物葉功能性狀及其相互關系. 中國沙漠, 2015, 35(6): 1489- 1495.
[22] Wilson P J, Thompson K, Hodgson J G. Specific leaf area and leaf dry matter content as alternative predictors of plant strategies. New Phytologist, 1999, 143(1): 155- 162.
[23] de la Riva E G, Tosto A, Pérez-Ramos I M, Navarro-Fernández C M, Olmo M, Anten N P R, Maraón T, Villar R. A plant economics spectrum in Mediterranean forests along environmental gradients: is there coordination among leaf, stem and root traits? Journal of Vegetation Science, 2016, 27(1): 187- 199.
[24] Wright I J, Cannon K. Relationships between leaf lifespan and structural defences in a low-nutrient, sclerophyll flora. Functional Ecology, 2001, 15(3): 351- 359.
[25] Agustí S, Enríquez S, Frost-Christensen H, Sand-Jensent K, Duarte C M. Light harvesting among photosynthetic organisms. Functional Ecology, 1994, 8(2): 273- 279.
[26] Vile D, Garnier E, Shipley B, Laurent G, Navas M L, Roumet C, Lavorel S, Díaz S, Hodgson J G, Lloret F, Midgley G F, Poorter H, Rutherford M C, Wilson P J, Wright I J. Specific leaf area and dry matter content estimate thickness in laminar leaves. Annals of Botany, 2005, 96(6): 1129- 1136.
[27] 羅璐, 申國珍, 謝宗強, 喻杰. 神農架海拔梯度上4種典型森林的喬木葉片功能性狀特征. 生態學報, 2011, 31(21): 6420- 6428.
[28] Reich P B, Walters M B, Ellsworth D S, Vose J M, Volin J C, Gresham C, Bowman W D. Relationships of leaf dark respiration to leaf nitrogen, specific leaf area and leaf life-span: a test across biomes and functional groups. Oecologia, 1998, 114(4): 471- 482.
[29] Witkowski E T F, Lamont B B. Leaf specific mass confounds leaf density and thickness. Oecologia, 1991, 88(4): 486- 493.
[30] 楊建軍, 蘇文華, 王玲玲, 楊銳, 楊波, 張博睿. 高山栲葉性狀種內變異及其與環境因子的關系. 廣東農業科學, 2015, 42(12): 153- 158.
[31] 宋光, 溫仲明, 鄭穎, 丁曼. 陜北黃土高原刺槐植物功能性狀與氣象因子的關系. 水土保持研究, 2013, 20(3): 125- 130.
[32] 施宇, 溫仲明, 龔時慧, 宋光, 鄭穎, 丁曼. 黃土丘陵區植物功能性狀沿氣候梯度的變化規律. 水土保持研究, 2012, 19(1): 107- 111, 116- 116.
[33] 呂金枝, 苗艷明, 張慧芳, 畢潤成. 山西霍山不同功能型植物葉性特征的比較研究. 武漢植物學研究, 2010, 28(4): 460- 465.
[34] Chen F S, Niklas K J, Zeng D H. Important foliar traits depend on species-grouping: analysis of a remnant temperate forest at the Keerqin Sandy Lands, China. Plant and Soil, 2011, 340(1): 337- 345.
[35] Wright I J, Reich P B, Westoby M, Ackerly D D, Baruch Z, Bongers F, Cavender-Bares J, Chapin T, Cornelissen J H C, Diemer M, Flexas J, Garnier E, Groom P K, Gulias J, Hikosaka K, Lamont B B, Lee T, Lee W, Lusk C, Midgley J J, Navas M L, Niinemets ü, Oleksyn J, Osada N, Poorter H, Poot P, Prior L, Pyankov V I, Roumet C, Thomas S C, Tjoelker M G, Veneklaas E J, Villar R. The worldwide leaf economics spectrum. Nature, 2004, 428(6985): 821- 827.
[36] 鄭淑霞, 上官周平. 不同功能型植物光合特性及其與葉氮含量、比葉重的關系. 生態學報, 2007, 27(1): 171- 181.
[37] 宋彥濤, 周道瑋, 王平, 李強. 松嫩草地66種草本植物葉片性狀特征. 生態學報, 2013, 33(1): 79- 88.
[38] Hodgson J G, Montserrat-Martí G, Charles M, Jones G, Wilson P, Shipley B, Sharafi M, Cerabolini B E L, Cornelissen J H C, Band S R, Bogard A, Castro-Díez P, Guerrero-Campo J, Palmer C, Pérez-Rontomé M C, Carter G, Hynd A, Romo-Díez A, de Torres Espuny L, Royo Pla F. Is leaf dry matter content a better predictor of soil fertility than specific leaf area? Annals of Botany, 2011, 108(7): 1337- 1345.
[39] 鄭元潤. 大青溝植物群落穩定性研究. 生態學報, 1999, 19(4): 578- 580.
Leaf traits of dominant plants of main forest communities in Daqinggou Nature Reserve
LIU Guifeng1,*, LIU Yuping1, BAIYILA Dafu2, CHENG Weiyan1, GAO Xuelei1, JIANG Lili3
1AgriculturalCollege,InnerMongoliaUniversityfortheNationalities,Tongliao028000,China2TheManagementBureauofDaqinggouNationalProtectionRegion,Ganqika028000,China3KeyLaboratoryofAlpineEcologyandBiodiversity,InstituteofTibetanPlateauResearch,ChineseAcademyofSciences,Beijing100101,China
Plant functional traits have been important issues in ecological research in recent years. Leaf functional traits have close relationships with the biomass, resource acquisition and utilization, and resource-use efficiency of plants. Daqinggou forest plant communities are located in the Horqin Desert, where the habitat conditions are very distinctive. There is a dense forest in the sand ditch, which forms a sharp contrast with the surrounding dune landscape. Although the difference in elevation from the bottom to the top of the ditch is only 60—70 m, different forest communities have developed because of differences in soil conditions that have arisen at different distance from the water source in the bottom of the ditch.Ulmusmacrocarpa,Quercusmongolica, andFraxinusmandshuricacommunities are respectively located in the top, middle and bottom of the ditch in Daqinggou Nature Reserve. In order to study the different environmental conditions in the Daqinggou Nature Reserve,U.macrocarpa,Q.mongolica, andF.mandshuricacommunities were selected as the three main research objects. We measured five functional traits of dominant species in the main forest communities, namely, leaf thickness, specific leaf area, leaf dry matter content, leaf size, and leaf dry weight. The relationships between the different leaf functional traits were studied, and the leaf functional traits of different growth forms and different communities were compared. Correlation analysis showed that leaf thickness was significantly negatively correlated with specific leaf area and significantly positively correlated with leaf size and leaf dry weight. Specific leaf area was significantly negatively correlated with leaf dry matter content and leaf dry weight, and positively correlated with leaf size. Leaf dry matter content was significantly negatively correlated with leaf size and significantly positively correlated with leaf dry weight, whereas leaf size was significantly positively correlated with leaf dry weight. Analysis of the different growth forms showed that the leaf dry matter content of herbs was lower than that of trees and shrubs. However, the specific leaf area of herbs was higher than that of trees and shrubs. Comparison of the functional traits of different communities showed that in theU.macrocarpaandQ.mongolicacommunities, the leaf thickness and leaf dry matter content of trees and shrubs were significantly higher than those in theF.mandshuricacommunity. In contrast, the specific leaf area of trees and shrubs in theU.macrocarpaandQ.mongolicacommunities were significantly lower than those in theF.mandshuricacommunity. The higher leaf dry matter content and lower specific leaf area of trees and shrubs in theU.macrocarpaandQ.mongolicacommunities reflect their adaptation to a more arid environment. Similarly, the lower leaf dry matter content and higher specific leaf area of trees and shrubs in theF.mandshuricacommunity reflect their adaptation to a moister environment. The species compositions in three main forest communities in the Daqinggou Nature Reserve have been regulated by the development of specific combinations of functional traits, which have enabled these species to adapt to the specific local environments.
Daqinggou Nature Reserve; forest community; dominant species; leaf traits; growth form

附錄1 各樣地優勢種組成
小葉樸CeltisbungeanaBl.;雞桑MorusaustralisPoir.;山杏Armeniacasibirica(Linn.) Lam.;葉底珠Securinegasuffruticosa(Pall.) Rehd.;榆樹UlmuspumilaL.;元寶楓AcertruncatumBunge稠李Padusracemosa(Lam.) Gilib.;春榆UlmusdavidianaPlanch var.;japonica(Rehd.)Nakai.;茶條槭AcerginnalaMaxim.山荊子Malusbaccata(Linn.) Borkh.;黃檗PhellodendronamurenseRupr.小葉鼠李RhamnusparvifoliaBunge.;山里紅CrataeguspinnatifidaBge.;鼠李RhamnusdavuricaPall.;衛矛Euonymusalatus(Thunb.)Sieb.;金銀木Loniceramaackii(Rupr.)Maxim.;羊草AneurolepidiumchinensisKitag.;黃精PolygonatumsibiricumRedoute.;大麻Cannabis sataiva L.;大籽蒿Artemisia sieversiana Ehrhart ex Willd.;抱莖苦荬菜IxerissonchifoliaBunge Hance.;益母草Leonurusjaponicushouttuyn.;細葉益母草LeonurussibiricusL.;蘆葦PhragmitescommunisTrin.;野艾蒿ArtemisialavandulaefoliaDC.;野古草Arundinellahirta(Thunb.) Tanaka.;蘿藦MetaplexisjaponicaMakino.;箭頭唐松草ThalictrumsimplexL.;萬年蒿ArtemisiasacrorumLedeb.;玉竹Polygonatumodoratum(Mill.)Druce var.;pluriflorum(Mig) Ohwi;野稗Echinochloacrusgalli(L.) Beauv.;紫苞鳶尾IrisruthenicaKer-Gawler.;虎尾草ChlorisvirgataSw.;龍牙草AgrimoniapilosaLedeb.;雞腿堇菜ViolaacuminataLedeb.;大花杓蘭CypripediummacranthumSw.;東北天南星ArisaemaamurenseMaxim.;鼠掌老鸛草GeraniumsibiricumL.;牛蒡子ArctiumlappaL.;牛尾菜SmilaxnipponicaMiq.;落新婦AstilbechinensisMaxim.;翼果苔草CarexneurocarpaMaxim.;水芹Oenanthejavanica(Blume.) DC;禾稈蹄蓋蕨Athyriumyokoscense(Franch.et.Sav.);羊乳Codonopsislanceolata(Sieb.;et Zucc.) Trautv.;透骨草PhrymaleptostachyaL.;subsp.;asiatica(Hara) Kitamura
國家自然科學基金(31260108;41301600);內蒙古自治區人才開發基金
2016- 03- 29; 網絡出版日期:2017- 03- 02
10.5846/stxb201603290575
*通訊作者Corresponding author.E-mail: liuguifeng1234@126.com
劉貴峰,劉玉平,達福白乙拉,程偉燕,高學磊,姜麗麗.大青溝自然保護區主要森林群落優勢種的葉性狀.生態學報,2017,37(14):4646- 4655.
Liu G F, Liu Y P, Baiyila D F, Cheng W Y, Gao X L, Jiang L L.Leaf traits of dominant plants of main forest communities in Daqinggou Nature Reserve.Acta Ecologica Sinica,2017,37(14):4646- 4655.
猜你喜歡
中華詩詞(2022年6期)2022-12-31 06:41:24
中國科技論壇(2017年7期)2017-07-25 08:49:53
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
媽媽寶寶(2017年2期)2017-02-21 01:21:24
爆笑show(2016年7期)2017-02-09 09:36:13
國際漢語學報(2016年1期)2017-01-20 08:21:20
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·兒童版(2015年1期)2015-07-07 04:12:52
