海水中藻菌共培養體系對碳氮磷的吸收轉化
2017-08-28 15:14:37張艷敏王江濤譚麗菊
生態學報 2017年14期
張艷敏,王江濤,譚麗菊
中國海洋大學化學化工學院, 青島 266100
海水中藻菌共培養體系對碳氮磷的吸收轉化
張艷敏,王江濤,譚麗菊*
中國海洋大學化學化工學院, 青島 266100
海洋環境中,細菌和微藻之間的物質交換是生源要素在自然界中遷移轉化的重要方式。為進一步了解生源要素的生物地球化學循環,在實驗室模擬條件下,研究了共培養體系中營養鹽和有機物在細菌和微藻之間的轉換。通過純培養中肋骨條藻(Skeletonemacostatum)、東海原甲藻(Prorocentrumdonghaiense)、天然海水中的細菌以及藻菌混合培養,分析了營養鹽和有機物隨藻菌生物量的變化情況,并計算了溶解有機碳(DOC)和溶解有機氮(DON)的濃度比值[(DOC/DON)a]。結果發現,在共培養體系中,細菌對中肋骨條藻的生長有抑制作用,對東海原甲藻影響不明顯;中肋骨條藻有利于細菌生長,東海原甲藻抑制細菌生長,這種不同可能與微藻的粒徑有關。海洋細菌在2種藻的指數生長均期均會促進微藻吸收氨氮(NH4-N),但在生長末期NH4-N以釋放為主。硝氮(NO3-N)的濃度與藻的生長呈負相關,但在衰亡期NO3-N略有增加,表明NO3-N再生所需時間較長。細菌對硝氮的吸收量較少,但對其再生有貢獻。細菌和中肋骨條藻對磷酸鹽(PO4-P)的吸收存在競爭,但與東海原甲藻的競爭關系不明顯。不同培養體系中DOC濃度變化不同,在藻菌共培養體系中增加較快,純藻培養體系中增加緩慢,在純菌培養體系中緩慢減少。通過對DOC與DON濃度比值的分析,發現用判斷顆粒有機碳(POC)來源的方法可以分析DOC的來源。
細菌;共培養體系;生源要素;溶解有機碳
細菌和微藻在自然界水體中廣泛存在,它們對促進水環境中的物質循環、維持水生環境的生態平衡有非常重要的作用[1]。細菌和微藻有著獨特的生態關系,藻、菌之間的相互作用對物質在微食物網中的生物化學循環起著關鍵作用[2-3]。藻類的水華是以消耗水體中的營養鹽為代價的[4],藻類將海水中的營養元素轉化為自身生長所需物質,通過細胞分泌、尸體自溶和細胞吞噬等方式向環境釋放有機物,同時,溶解有機物又可以為浮游細菌提供食物來源[5-6],細菌吸收分解有機物后以無機或其他形式釋放回海水中,為藻類提供生長所必須的營養物質[7],完成生源要素的微循環。作為主要的分解者,細菌對藻類的作用主要集中在營養改善(生源要素的提供、生長因子的轉化)、信息素調節和協同保護[8-9]。藻類的生長、增殖和死亡,除了與環境中的溫度、光照、鹽度、營養物質等理化因子有密不可分的關系外,還與海洋細菌有密切的關系[10]。
微生物驅動的碳、氮循環過程是全球生物地球化學循環的重要組成部分。海洋浮游生物對全球碳循環有深遠的影響,光合微生物是形成海洋食物鏈的基礎,但單細胞真核生物與有機體的相互作用對碳循環的影響遠超出光合微生物的影響[11]。有研究發現,營養鹽對異養細菌分解溶解有機碳的影響不大[12],細菌濾液對藻細胞增殖的影響與細菌濃度、細菌的種屬特性以及藻細胞自身的生長階段有關,細菌濃度較高對藻細胞生物量具有明顯的抑制作用,而濃度較低則顯示了一定程度的促進作用[13]。細菌對含有不同生源要素有機物的分解利用存在差異,具有相同結構但不同生源要素有機物的礦化速率不同[14]。氮是細胞生物量中第4個最豐富的元素,生物固氮對海洋氮循環具有重要作用,生物固氮效率的高低不僅會造成生態系統從氮限制向磷限制轉變,還會對寡營養海域的新生產力和磷循環產生重要的影響[15-16]。磷是浮游植物和細菌生長的必需元素,磷的重新分布可以造成微生物群落重塑[17-18]。由此可見,海洋浮游細菌的存在及生命活動決定著水域食物鏈基本環節的發展[19-20]。近幾年來,關于自然環境中碳氮磷循環的研究、微生物對有機物的礦化作用研究以及藻類對營養鹽的吸收利用研究越來越多[21-22],但關于營養鹽和有機物在藻菌之間轉換關系的研究較少。本文通過室內模擬實驗,在共培養條件下對營養鹽和有機物在藻菌之間的轉化關系進行探討,以期了解海洋微生物在生源要素轉換過程中的作用,進一步探索生源要素的生物地球化學循環規律。
1 材料和方法
1.1 水樣采集和處理
水樣于2015年7月采集于青島近海,采集深度5m。水樣采集后,先用孔徑為0.45μm的聚碳酸酯濾膜過濾,除去浮游藻類,濾液即為實驗所用海水。
1.2 藻種及培養條件
實驗選取中國東海最常見藻種——中肋骨條藻(Skeletonemacostatum)和東海原甲藻(Prorocentrumdonghaiense)進行培養,均來自中國海洋大學海洋污染生態化學實驗室藻種庫,用f/2培養液培養到指數生長期時開始接種。細菌來自天然海水中的混合菌種,主要包括:變形桿菌門(Proteobacteria)、藍細菌門(Cyanobacteria)、擬桿菌門(Bacteroidetes)和浮霉菌門(Planctomycetes)4個門類,其中α-Proteobacteria為優勢菌,其次為藍細菌門和擬桿菌門[23]。培養體系的細菌初始密度約為3.1×104細胞/mL。
設置6組培養體系。分別為菌純培養(BPC);中肋骨條藻純培養(APC-S);東海原甲藻純培養(APC-P);中肋骨條藻-細菌混合培養(MC-S);東海原甲藻-細菌混合培養(MC-P)和空白對照組。其中,對照組為高溫滅菌海水,其他實驗組添加微生物情況見表1。所有實驗組均在聚四氟乙烯桶中,置于光照培養箱中培養,溫度為20℃,光照周期為12(L)∶12(D),光照度4000lx,光強70μmol光子 m-2s-1。中肋骨條藻的初始藻密度約為3×104細胞/mL;東海原甲藻的初始藻密度約為1.3×103細胞/mL。
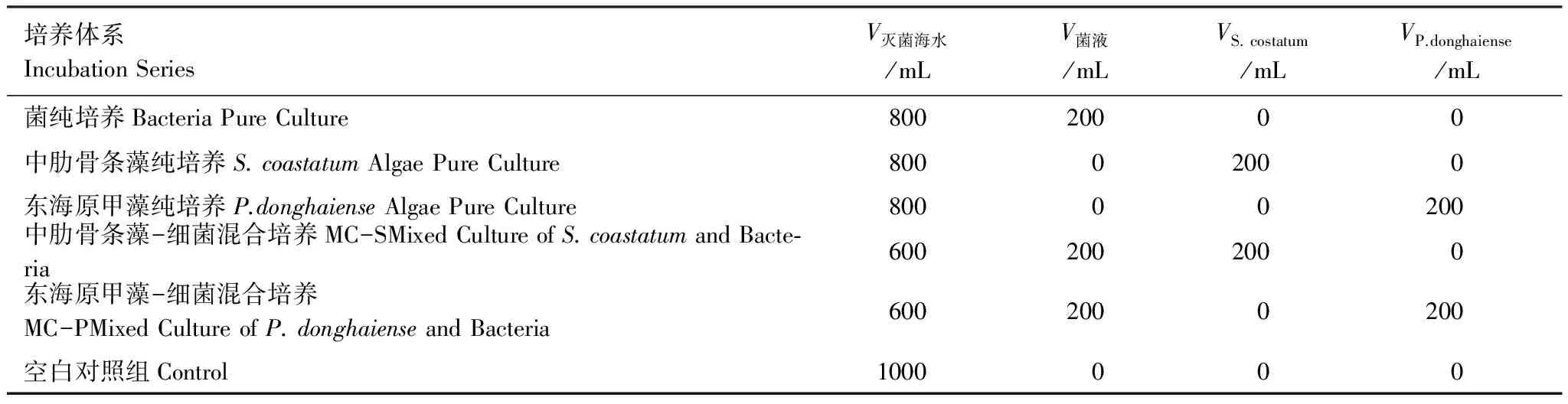
表1 培養體系
V滅菌海水:滅菌海水的體積the volume of sterilized sea water;V菌液:含細菌的溶液體積the volume of solution containing bacteria;VS.costatum:中肋骨條藻藻液the volume of S.costatum;VP.donghaiense:東海原甲藻藻液the volume of P.donghaiense
1.3 生物量計數
細菌計數采用DAPI(4′6-二脒基- 2-苯基吲哚)染色,熒光顯微鏡(Leica DM4000,德國萊卡公司)計數的方法[14]。細菌豐度的表達式為:
BB(個/mL)=A×S1/(S2×V)[14]
式中,BB為細菌豐度;A為10個視野的平均細菌數;S1為濾膜的有效過濾面積;S2為視野面積;V為過濾水樣體積。微藻生物量在加入魯格試劑后,用電子顯微鏡直接計數[14]。
1.4 營養鹽添加和測定
向培養體系中加入修改后的f/2營養配方(添加后NO3-N濃度為18μmol/L,PO4-P的濃度為1.5μmol/L,氨氮及其他元素濃度不變),測試不同營養條件菌藻混合培養對生源要素轉移的影響。采用TechniconTMAA-Ⅲ型營養鹽自動分析儀測定濾液中營養鹽濃度[24]。
1.5 DOC、TN的測定
培養液中的溶解有機碳(DOC)、總氮(TN)采用高溫燃燒法,用有機碳自動分析儀(Shimadzu TOC-VCPH)測定[25]。為避免可能存在的污染,樣品從培養桶中直接取樣到測樣瓶中[14](含藻的實驗組需經0.45μm濾膜過濾)。DOC含量包括浮游細菌的碳含量。DOC的分析精度±1μmol/L,變異系數CV為1%—2%。空白對照,精度為±0.3μmol/L,CV<1%。TN的分析精度±1μmol/L,變異系數CV<2%。
2 結果和討論
2.1 微藻和細菌生物量的變化
2.1.1 微藻生物量的變化
不同體系中肋骨條藻和東海原甲藻生物量的變化曲線如圖1所示。在其他條件一致的情況下,中肋骨條藻的生物量在細菌是否存在條件下的生長趨勢比較類似,即:在培養初期藻細胞密度指數增長,達到最大值后,趨于穩定然后下降。但在MC-S體系中,最大藻密度低于APC-S體系,說明混合培養過程中,藻的生長受到了細菌的抑制作用。由于細菌的比表面積較大,使其對營養鹽有更強的親和力,導致細菌吸收了相對較多的營養鹽,造成微藻可利用的營養鹽減少,從而抑制了微藻的生長[26]。對于東海原甲藻,細菌對其沒有明顯影響。已有研究發現,在低磷酸鹽濃度的培養液中,細菌存在時,外源碳的加入會刺激細菌對磷酸鹽的吸收,對東海原甲藻細胞吸收磷產生競爭,從而限制藻的生長;無外源碳加入時,細菌的存在對東海原甲藻的生長影響不明顯[27],本實驗培養體系中無外源碳加入,從而細菌對東海原甲藻的生長沒有明顯影響。
關于中肋骨條藻和東海原甲藻對營養鹽的競爭過程及結果的研究發現,由于中肋骨條藻對營養鹽的利用方式及較高的生長率[28],中肋骨條藻是完全的優勢藻,且兩種藻的競爭結果并不依賴于初始接種密度比例[29]。而在營養鹽相對匱乏條件下,東海原甲藻會在中肋骨條藻的競爭中取得優勢[30]。說明,實驗條件以及微藻自身利用營養鹽的方式及效率等因素影響藻的生長,這也是相同密度的細菌對中肋骨條藻和東海原甲藻生長影響不同的原因。
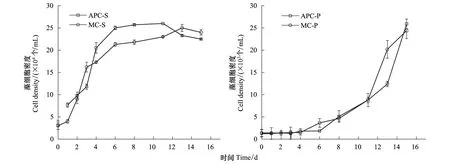
圖1 不同培養體系中藻的生物量變化曲線Fig.1 Biomass of algae in different cultivating systemsAPC-S:中肋骨條藻純培養S. coastatum Algae Pure Culture;MC-S:中肋骨條藻-細菌混合培養Mixed Culture of S. coastatum and Bacteria;APC-P:東海原甲藻純培養P. donghaiense Algae Pure Culture;MC-P:東海原甲藻-細菌混合培養Mixed Culture of P. donghaiense and Bacteria
2.1.2 細菌生物量的變化
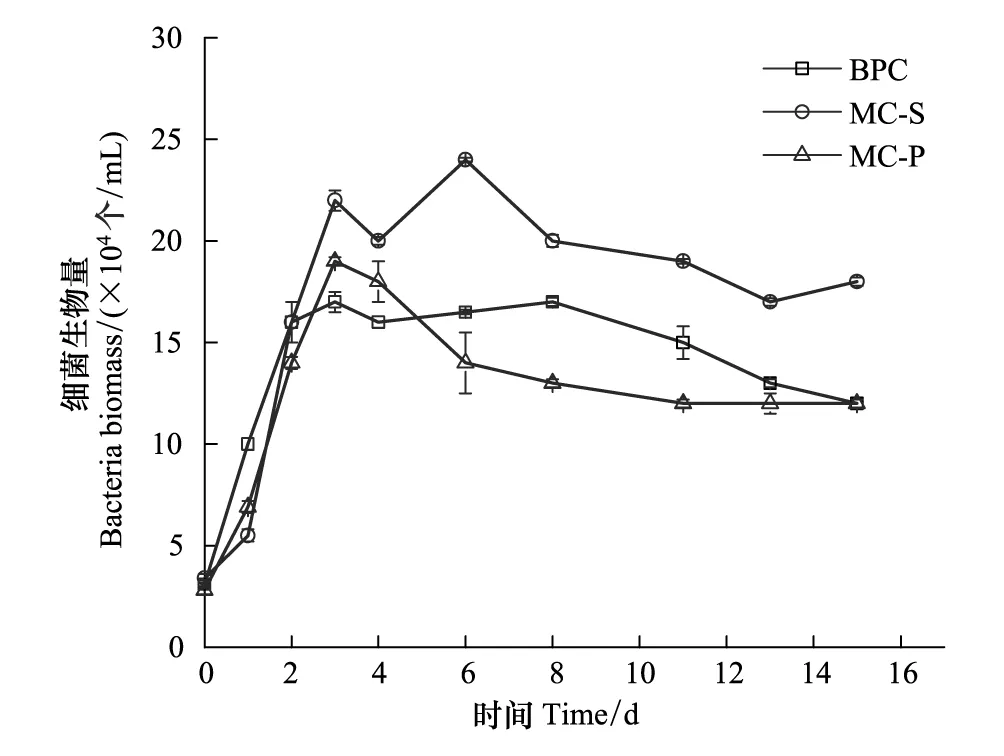
圖2 不同培養體系中細菌生物量變化曲線Fig.2 Biomass of bacteria in different cultivating systemsBPC為菌純培養Bacteria Pure Culture
不同體系中細菌生物量的變化曲線如圖2所示。在培養的前3d,細菌都處于指數生長期,在短暫的穩定期后,進入緩慢的衰亡期。MC-S體系的細菌密度最大,BPC體系的細菌生物量居中,MC-P體系的細菌密度相對略小,說明藻的存在影響細菌的生長狀況,中肋骨條藻的存在可以促進菌的生長,但東海原甲藻對細菌的生長略有抑制作用。營養鹽限制條件下,細菌的生長主要受可利用的氮、磷營養鹽的限制[31]。東海原甲藻與細菌混合培養體系中不添加外源碳,細菌只能依賴微藻光合作用產生的有機碳,對磷酸鹽的競爭力減弱,從而其生長受到了抑制[22]。
從生理學角度看,小細胞的比表面積較大,有利于低分子量代謝物通過細胞膜擴散,因此粒徑較小的浮游植物對DOC的釋放速率較高[32-33],DON的釋放可能是通過膜被動運輸的,較小微藻的DON釋放效率更高[34]。因此,較小的中肋骨條藻(粒徑<20μm)[35]對DOC和DON的釋放速率比東海原甲藻(22μm<粒徑<70μm)的釋放速率高,這可能是中肋骨條藻促進細菌生長而東海原甲藻抑制細菌生長的原因。
2.2 營養鹽的濃度變化
2.2.1 氨氮
氨氮在培養體系中的濃度變化如圖3所示。不同培養體系中的NH4-N濃度均是先升高,后降低。其中,細菌單獨培養體系NH4-N變化幅度最小,藻純培養體系NH4-N變化幅度最大,混合培養體系處于中間水平,但混合培養體系的末期NH4-N濃度有小幅的升高。氨氮是有機氮礦化的第一個無機產物[36],浮游植物吸收硝酸鹽同化成氨或DON[37],因此,氨氮通常與細菌分解有關。已有研究表明,雖然中肋骨條藻對氨氮更具親和力,但由于中肋骨條藻對磷酸鹽的需求很高,在磷酸鹽很低的情況下,藻的生長受到限制,也影響了它對氨氮的吸收[38]。已有類似研究發現,小球藻在氨氮較高時,對NH4-N具有較好的去除能力[39],但細菌的存在[30]和營養結構的改變會使小球藻去除氮、磷能力存在較大差異[40]。
本論文的結果表明,細菌的存在對中肋骨條藻吸收氨氮的能力有促進作用,在生長周期結束時,氨氮以釋放為主。對于東海原甲藻,由于生長周期較長,在指數生長期時對氨氮的去除能力有促進,但指數生長末期,氨氮以增加為主,反映了此時細菌已經開始轉化有機物為氨氮。有研究表明,22℃溫度時,浮游生物將50%的有機氮分解為NH4-N時,需3—7d;將90%的有機氮分解為NH4-N時,需9—23d[36],可見,此時的氨氮來源于細菌的分解。
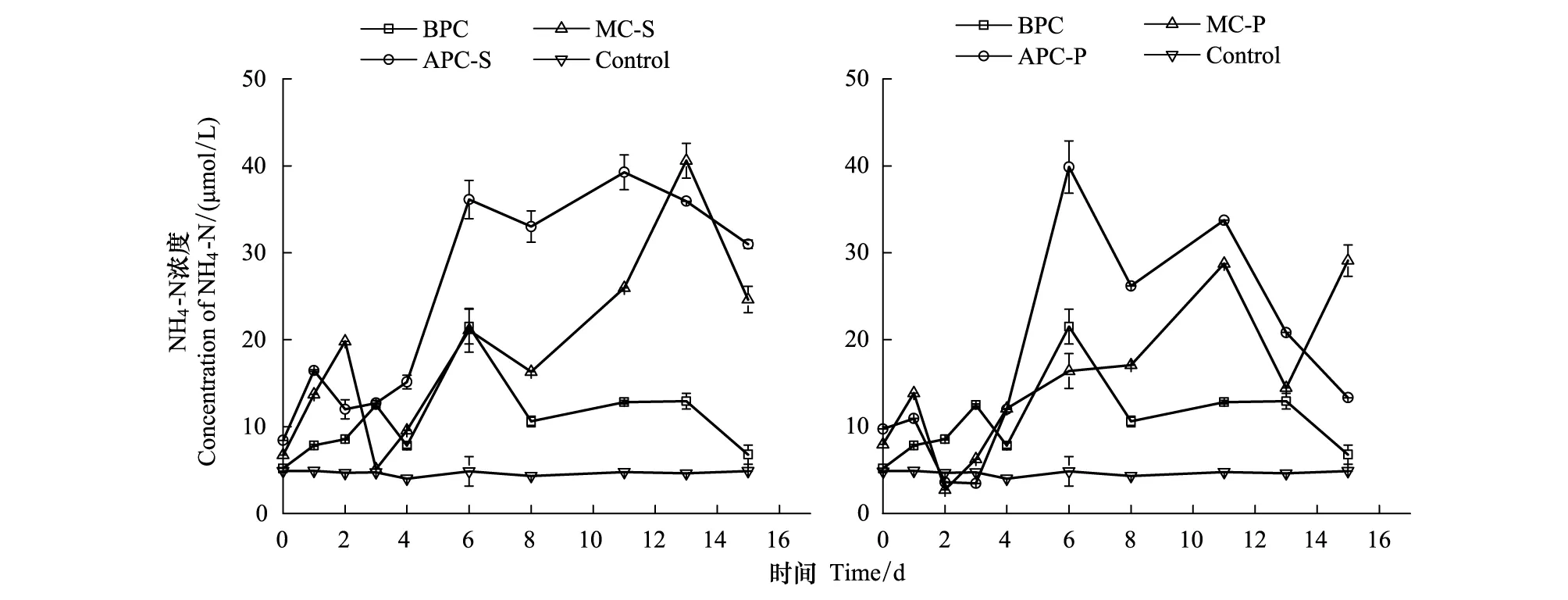
圖3 培養體系中NH4-N的濃度變化Fig.3 The concentration variation of NH4-N
2.2.2 硝氮
在培養過程中,NO3-N的濃度變化如圖4所示。細菌純培養時NO3-N濃度變化很小,表明細菌在生長過程中對NO3-N的需求量非常少。有研究發現,膠州灣海域異養浮游細菌對硝酸鹽的吸收作用要小于其釋放作用[41],異養浮游細菌在生長過程中優先利用有機氮(氨基酸)和銨鹽作為其氮源[41-42]。藻菌混合培養和微藻單獨培養時,NO3-N的濃度均大幅降低,由于中肋骨條藻進入指數生長期較早,所以NO3-N消耗的更快。另外,從3種形態無機氮(NH4-N, NO3-N, NO2-N)的含量變化來看,只有NO3-N濃度同藻密度存在一定程度的負相關,因此微藻的生長主要消耗NO3-N,與文獻研究結果一致[43]。NO3-N可能還與異養細菌分解有機質再生營養鹽有關[19],研究發現膠州灣中肋骨條藻赤潮過后無機氮恢復較快[43],說明異養浮游細菌對NO3-N的再生有貢獻。
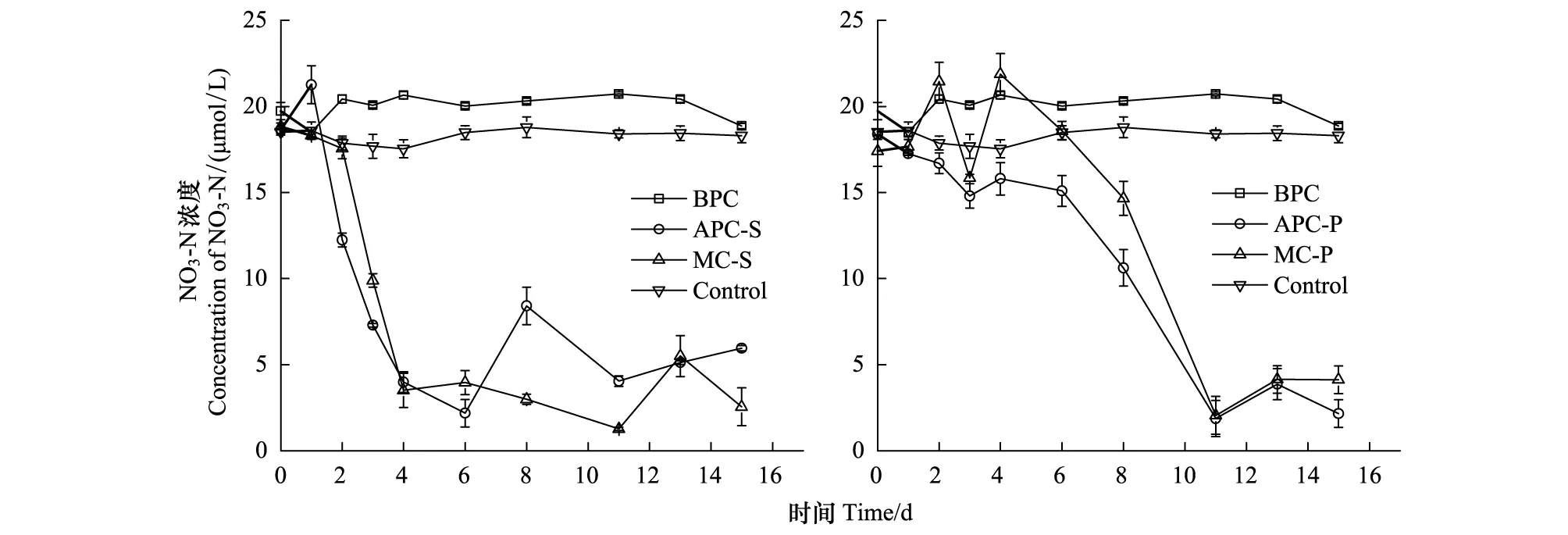
圖4 培養體系中NO3-N的濃度變化Fig.4 The variation of concentration of NO3-N
2.2.3 磷酸鹽
培養過程中,無機PO4-P濃度變化如圖5。空白對照組中PO4-P濃度變化不大。細菌純培養的培養液中PO4-P消耗量較多,大約占PO4-P初始量的50%。中肋骨條藻實驗組中,相同時間內,混合培養體系消耗的PO4-P濃度少于或等于藻類單獨培養體系。說明在低PO4-P濃度下,細菌與藻存在PO4-P競爭,細菌的存在影響了藻類消耗PO4-P的速率。實驗末期,PO4-P濃度都低于檢測限。混合培養體系中,中肋骨條藻的生長低于純培養體系中藻的生長,也說明細菌的存在,影響了藻類吸收PO4-P,與文獻結果一致[37,44]。東海原甲藻實驗組中,相同時間內,混合培養體系消耗的PO4-P濃度與藻類單獨培養體系沒有明顯差異,且混合培養體系中,東海原甲藻的生長與純培養體系中藻的生長也沒有明顯差異。

圖5 培養體系中磷酸鹽的濃度變化Fig.5 The concentration variation of PO4-P
2.3 培養過程中DOC的濃度變化
不同培養體系中DOC的濃度變化如圖6所示。由圖可見,在APC-S和APC-P體系,即微藻單獨培養時,DOC濃度與藻密度有一定的正相關性。在藻菌混合體系,即MC-S和MC-P,在培養初始的2—6d內,體系中DOC濃度的增加速率大于APC-S/P體系。分析原因為:在細菌存在時,細菌對藻的抑制作用包括溶藻酶類的產生[7,45],分解死亡藻細胞從而釋放更多的有機物。藻細胞釋放和分泌的易降解有機物占總的溶解有機物的一小部分[26],細菌分解易降解的有機物,大部分不易降解的有機物得到累積使有機物的濃度增加。另有研究發現,細菌的生物量變化滯后于有機物濃度變化10—20h[12],DOC在細菌單獨存在時,前3天濃度降低明顯,之后緩慢降低,結合圖2,表明細菌在指數生長期分解DOC較快,之后分解速率沒有明顯變化。
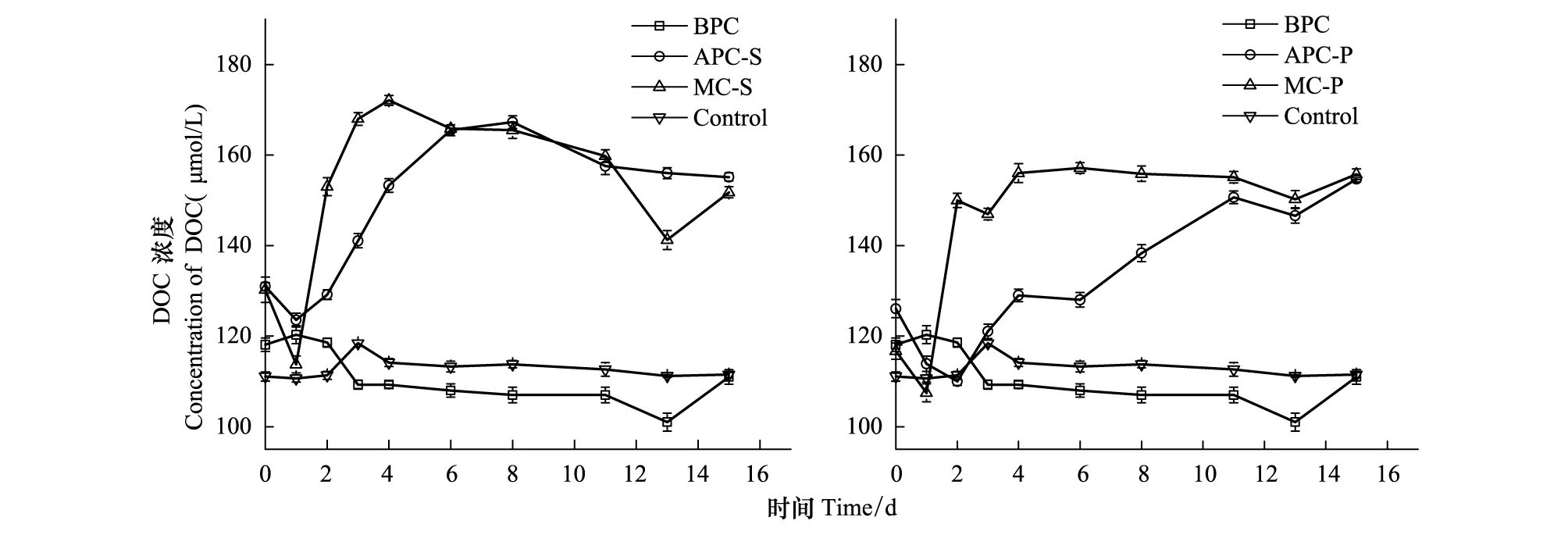
圖6 培養過程中DOC的濃度變化Fig.6 The concentration variation of DOC during the culture process
2.4 培養過程中DOC/DON的濃度變化
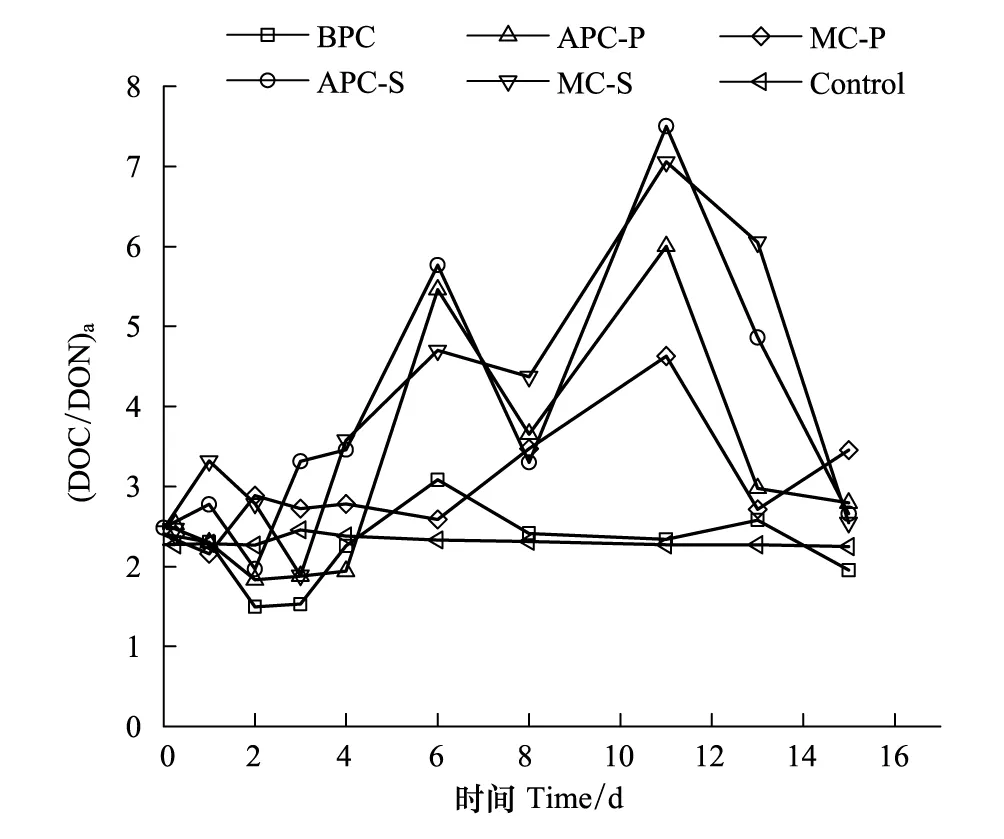
圖7 培養體系中(DOC/DON)aFig.7 The Ratio of (DOC/DON)a in ambient solution
海洋是全球最大的有機碳儲蓄庫之一,有機碳主要以DOC和POC的形式存在[45-46]。POC在水體中的含量變化與微藻的初級生產力有關,是評價海區初級生產力水平的一個重要參數[33]。海洋微藻通過光合作用合成有機碳是DOC的重要來源之一[20,25]。水體中DON是微生物的重要氮源,主要由微藻釋放[20,25]。根據POC與PON的摩爾比((POC/PON)a),可以判斷海區的POC來源[47],對研究藻類與異養細菌之間的相互作用以及生物匯集碳的速率和機制提供理論。Cifuentes等人認為,(POC/PON)a在2.6—4.3之間POC來源為細菌;7.7—10.1之間為浮游植物;>50為高等植物,即為陸源[48]。本文根據相同的原理,研究了DOC/DON的摩爾比((DOC/DON)a)用于判斷DOC來源的可能性。培養過程中(DOC/DON)a的變化情況如圖7所示。
由圖7可見,培養過程中,空白對照組的(DOC/DON)a始終低于3,變化不明顯。對于其他培養體系,在初期(0—5d),各體系(DOC/DON)a的波動變化,整體呈上升趨勢,比值介于2—5,說明實驗初期,細菌礦化作用較明顯,DOC主要來源于細菌。培養中后期(6—14d),除第8天,含藻培養體系中C/N比值高于5,甚至超過7,說明此過程中細菌和藻相互作用,而藻的作用要顯著于細菌的礦化作用,DOC既來源于細菌也來源于微藻。培養末期(第15天),藻衰亡,所以C/N比值有所下降,細菌分解藻釋放DOC,來源于微藻的DOC較少。培養過程中,含藻體系的C/N比值高于含細菌體系的C/N比值,與理論C/N比值相符。而且APC-S的C/N比值最大,其次是MC-S、APC-P、MC-P,最后是BPC。以上分析表明,可以根據判斷POC來源的方法判斷DOC的來源,與實際情況相符。
通過以上結果可知,細菌的存在可以影響中肋骨條藻的生長、對營養鹽的吸收以及對DOC的釋放[26]。細菌的存在對東海原甲藻的生長和營養鹽的吸收沒有明顯的影響,與文獻[27]研究結果有差異,可能是細菌的生物量、藻細胞密度、藻粒徑大小以及營養鹽濃度不同導致的。Carlson通過對比添加NH4-N、PO4-P前后,細菌生物量和細菌生長效率等的變化發現,NH4-N和PO4-P不是影響細菌生長的主要因素,影響細菌生物量的主要因素是可利用有機物的量[12]。有機物礦化作用受很多因素的影響,比如異養細菌的生物量,微生物群落結構,海水溫度,有機物的組成等。自養生物分泌的有機物影響細菌對有機物的利用性,影響海洋生態系統中的營養動力學和生物地球化學循環[25]。研究發現,生物群落的組成及其對NO3-N的利用程度影響微生物吸收利用有機碳[47]。
3 結論
本文通過實驗室混合培養海洋微藻和細菌,發現細菌和微藻共存時,細菌通過對營養鹽的競爭限制某些微藻的生長,微藻通過對有機物的釋放或對營養鹽的競爭促進或抑制細菌的生長。培養液中DOC的濃度變化與藻密度密切相關,并提出可以用(DOC/DON)a來判斷DOC的來源。
[1] 王少沛, 曹煜成, 李卓佳, 楊鶯鶯, 陳素文. 水生環境中細菌與微藻的相互關系及其實際應用. 南方水產, 2008, 4(1): 76- 80.
[2] 王新, 周立紅, 鄭天凌, 寧修仁. 塔瑪亞歷山大藻藻際細菌溶藻過程. 生態學報, 2007, 27(7): 2864- 2871.
[3] Azam F. Microbial control of oceanic carbon flux: the plot thickens. Science, 1998, 280(5364): 694- 696.
[4] 劉浩, 潘偉然. 營養鹽負荷對浮游植物水華影響的模型研究. 水科學進展, 2007, 19(3): 345- 351.
[5] 曲長鳳, 宋金明, 李寧, 李學剛, 袁華茂, 段麗琴. 海水中沙海蜇消亡對水體碳、氮、磷的釋放與補充. 應用生態學報, 2016, 27(1): 299- 306.
[6] Condon R H, Steinberg D K, del Giorgio P A, Bouvier T C, Bronk D A, Graham W M, Ducklow H W. Jellyfish blooms result in a major microbial respiratory sink of carbon in marine systems. Proceedings of the National Academy of Sciences of the United States of America, 2011, 108(25): 10225- 10230.
[7] Wear E K, Carlson C A, Windecker L A, Brzezinski M A. Roles of diatom nutrient stress and species identity in determining the short-and long-term bioavailability of diatom exudates to bacterioplankton. Marine Chemistry, 2015, 177: 335- 348.
[8] 周進, 陳國福, 朱小山, 陳璐, 蔡中華. 赤潮過程中“藻-菌”關系研究進展. 生態學報, 2014, 34(2): 269- 281.
[9] Egan S, Harder T, Burke C, Steinberg P, Kjelleberg S, Thomas T. The seaweed holobiont: understanding seaweed-bacteria interactions. FEMS Microbiology Reviews, 2013, 37(3): 462- 476.
[10] 鄭天凌, 田蘊, 蘇建強, 王艷麗, 連玉武, 洪華生. 海洋赤潮生物與廈門海域幾種細菌的生態關系研究. 生態學報, 2002, 22(12): 2063- 2070.
[11] Worden A Z, Follows M J, Giovannoni S J, Wilken S, Zimmerman A E, Keeling P J. Rethinking the marine carbon cycle: factoring in the multifarious lifestyles of microbes. Science, 2015, 347(6223): 1257594.
[12] Carlson C A, Ducklow H W. Growth of bacterioplankton and consumption of dissolved organic carbon in the Sargasso Sea. Aquatic Microbial Ecology, 1996, 10(1): 69- 85.
[13] 蘇建強, 鄭天凌, 俞志明, 宋秀賢. 海洋細菌對赤潮藻生長及其產毒量的影響. 海洋與湖沼, 2003, 34(1): 44- 49.
[14] 譚麗菊, 肖慧, Carlson C A, 王江濤. 海水中天然細菌對不同生源要素有機物的礦化作用. 生態學報, 2016, 36(1): 77- 85.
[15] Stein L Y, Klotz M G. The nitrogen cycle. Current Biology, 2016, 26(3): R94-R98.
[16] 王艷君, 董園, 陳寅超, 周衛文, 李芊. 固氮作用對黑潮上游區域生態系統影響的模擬研究. 熱帶海洋學報, 2016, 35(1): 102- 111.
[17] 郭建國, 趙龍浩, 徐丹, 孫野青. 洪水對大凌河河流浮游細菌群落的影響. 生態學報, 2015, 35(14): 4777- 4783.
[18] Zhou Q X, Gibson C E, Zhu Y M. Evaluation of phosphorus bioavailability in sediments of three contrasting lakes in China and the UK. Chemosphere, 2001, 42(2): 221- 225.
[19] 白潔, 張昊飛, 李巋然, 張經, 劉東艷, 高冬梅, 易齊濤. 膠州灣冬季異養細菌與營養鹽分布特征及關系研究. 海洋科學, 2004, 28(12): 31- 34.
[20] Caron D A, Lim E L, Sanders R W, Dennett M R, Berninger U G. Responses of bacterioplankton and phytoplankton to organic carbon and inorganic nutrient additions in contrasting oceanic ecosystems. Aquatic Microbial Ecology, 2000, 22(2): 175- 184.
[21] 鮑林林, 陳永娟, 王曉燕. 河流沉積物氮循環主要微生物的生態特征. 微生物學通報, 2015, 42(6): 1141- 1150.
[22] 黃振華, 柯愛英, 劉俊峰, 葉深, 薛峰, 周朝生, 陳星星. 浙江樂清灣海域冬季顆粒有機碳的分布特征及影響因素. 福建水產, 2015, 37(3): 220- 226.
[23] 張艷. 山東近岸海域水體細菌多樣性研究[D]. 青島: 中國海洋大學, 2010.
[24] 潘建明, 周懷陽, 扈傳昱, 劉小涯, 董禮先, 張美. 夏季珠江口沉積物中營養鹽剖面分布和界面交換通量. 海洋學報, 2002, 24(3): 52- 59.
[25] Halewood E R, Carlson C A, Brzezinski M A, Reed D C, Goodman J. Annual cycle of organic matter partitioning and its availability to bacteria across the Santa Barbara Channel continental shelf. Aquatic Microbial Ecology, 2012, 67(3): 189- 209.
[26] 連子如, 王江濤. 不同濃度碳源下中肋骨條藻和海洋異養細菌對磷酸鹽的競爭吸收研究. 中國海洋大學學報, 2009, 39(S1): 237- 241.
[27] 連子如, 王江濤. 東海原甲藻和海洋異養細菌對磷酸鹽的競爭吸收. 水生生物學報, 2010, 34(3): 663- 668.
[28] 呂頌輝, 李英. 我國東海 4 種赤潮藻的細胞氮磷營養儲存能力對比. 過程工程學報, 2006, 6(3): 439- 444。
[29] 孫科, 丘仲鋒, 何宜軍, 尹寶樹. 東海原甲藻與中肋骨條藻的種間競爭數值模擬. 生態學報, 2014, 34(23): 6898- 6909.
[30] Zhu M Y, Xu Z J, Li R X, Wang Z L, Shi X Y. Interspecies competition for nutrients betweenProrocentrumdonghaienseLu andSkeletonemacostatum(Grev.) Cleve in mesocosm experiments. Acta Oceanologica Sinica, 2009, 28(1): 72- 82.
[31] Rodrigues R M N V, Le B Williams P J. Heterotrophic bacterial utilization of nitrogenous and nonnitrogenous substrates, determined from ammonia and oxygen fluxes. Limnology and Oceanography, 2001, 46(7): 1675- 1683.
[32] 劉誠剛, 寧修仁, 郝鏘, 樂鳳鳳. 海洋浮游植物溶解有機碳釋放研究進展. 地球科學進展, 2010, 25(2): 123- 132.
[33] Bj?rrisen P K. Phytoplankton exudation of organic matter: why do healthy cells do it?. Limnology and Oceanography, 1988, 33(1): 151- 154.
[34] 高亞輝, 荊紅梅, 黃德強, 楊心寧. 海洋微藻胞外產物研究進展. 海洋科學, 2002, 26(3): 35- 38.
[35] 孫軍, 劉東艷, 鐘華, 張利永. 浮游植物粒級研究方法的比較. 青島海洋大學學報, 2003, 33(6): 917- 924.
[36] 水化學研究組. 膠州灣海水中氮的地球化學(續). 海洋湖沼通報, 1982, (4): 37- 46.
[37] 洪義國. 硝酸鹽氮氧穩定同位素分餾過程記錄的海洋氮循環研究進展. 地球科學進展, 2013, 28(7): 751- 764.
[38] 丁光茂. 中國近海三種典型赤潮生物氮營養生理研究[D]. 廈門: 廈門大學, 2009.
[39] 白潔, 易齊濤, 李佳霖. 海洋異養細菌對小球藻生長和無機營養鹽吸收的影響研究. 海洋科學, 2009, 33(11): 6- 10.
[40] Watanabe K, Takihana N, Aoyagi H, Hanada S, Watanabe Y, Ohmura N, SaikiH, Tanaka H. Symbiotic association inChlorellaculture. FEMS Microbiology Ecology, 2005, 51(2): 187- 196.
[41] NagaosaK, MorikawaK. Fluctuations in bacterial community and environmental factors at the urban spring Masugata. Japanese Journal of Limnology, 1999, 60(2): 177- 184.
[42] Middelburg J J, Nieuwenhuize J. Nitrogen uptake by heterotrophic bacteria and phytoplankton in the nitrate-rich Thames estuary. Marine Ecology Progress Series, 2000, 203: 13- 21.
[43] 霍文毅, 俞志明, 鄒景忠, 宋秀賢, 郝建華. 膠州灣中肋骨條藻赤潮與環境因子的關系. 海洋與湖沼, 2001, 32(3): 311- 318.
[44] Bai J, Li K R, Zhang J, Li Z Y, Gao H W, Zhang H F. Distribution of biomass of heterotrophic bacterioplankton in the Bohai Sea. Chinese Journal of Oceanology and Limnology, 2005, 23(4): 427- 432.
[45] Skerratt J H, Bowman J P, Hallegraeff G, James S, Nichols P D. Algicidal bacteria associated with blooms of a toxic dinoflagellate in a temperate Australian estuary. Marine Ecology Progress Series, 2002, 244: 1- 15.
[46] 譚麗菊, 張哲, 梁成菊, 王江濤. 青島鄰近海域海水中有機碳的分布特征. 地球科學進展, 2011, 26(4): 426- 432.
[47] Cifuentes L A, Coffin R B, Solorzano L, Cardenas W, Espinoza J, Twilley R R. Isotopic and elemental variations of carbon and nitrogen in a mangrove estuary. Estuarine, Coastal and Shelf Science, 1996, 43(6): 781- 800.
[48] Letscher R T, Knapp A N, James A K, Carlson C A, Santoro A E, Hansell D A. Microbial community composition and nitrogen availability influence DOC remineralization in the South Pacific Gyre. Marine Chemistry, 2015, 177: 325- 334.
Uptake and transformation of carbon, nitrogen and phosphorus in the co-culture system of algae and bacteria in seawater
ZHANG Yanmin, WANG Jiangtao, TAN Liju*
CollegeofChemistryandChemicalEngineering,OceanUniversityofChina,Qingdao266100,China
In the marine environment, the exchange of the substance between bacteria and algae is an important way of the migration and transformation of biogenic elements in the natural world. In order to obtain a further understanding of the biogeochemical cycles of biogenic elements, the transformation of nutrients (nitrogen and phosphorus) and organic matter between marine bacteria and two types of algae (SkeletonemacostatumandProrocentrumdonghaiense) were studied in a co-culture or single-culture system of algae and bacteria based on a laboratory-simulated experiment. The changes of nutrient and organic carbon were tracked and the biomass of bacteria and the two algae were calculated. In addition, the concentration ratio [(DOC/DON)a] of dissolved organic carbon (DOC) to dissolved organic nitrogen (DON) in different culture systems was analyzed to determine the sources of organic matter. The results indicated that bacteria may inhibit the growth ofS.costatum, but have little effects on growth ofP.donghaiensein the co-culture system. On the contratry,S.costatumaccelerated the growth of bacteria, whereasP.donghaiensesuppressed the growth of bacteria, which is probably related to the different sizes of the two algae. During the exponential growth phase of the two algae, bacteria promoted the uptake of ammonia (NH4-N) by the algae, however NH4-N was mainly released at the end stage of algae growth. The concentrations of nitrate (NO3-N) and the biomass of algae showed a nearly negative correlation, but NO3-N increased slightly during the decline phase of algae, indicating that the regeneration of NO3-N took a longer time. Bacteria uptook little NO3-N, but contributed to the regeneration of NO3-N. Bacteria andS.costatumcompeted for phosphate (PO4-P) and bacteria affected the uptake of PO4-P byS.costatum. There were not obvious different between concentrations of PO4-P in co-culture ofP.donghaienseand bacteria from that in single culture ofP.donghaiense, which was in contrast to the results of the culture withS.costatum. In the course of the culture, the concentration of DOC varied in different cultural systems. DOC increased rapidly in the co-culture systems of bacteria and algae, whereas the concentration of DOC increased slowly in the pure algal systems, and decreased slowly in the single-culture bacterial system. The analysis of the concentration ratio of DOC and DON showed that the method of tracing the source of particulate organic carbon (POC) can also be used to trace the source of DOC.
bacteria; co-culture system; biogenic elements; dissolved organic carbon
國家海洋局海洋生態環境科學與工程重點實驗室開放基金(MESE- 2014-03); 科技部科技支撐計劃(2012BAF14B04)
2016- 04- 22; 網絡出版日期:2017- 03- 02
10.5846/stxb201604220761
*通訊作者Corresponding author.E-mail: lijutan@ouc.edu.cn
張艷敏,王江濤,譚麗菊.海水中藻菌共培養體系對碳氮磷的吸收轉化.生態學報,2017,37(14):4843- 4851.
Zhang Y M, Wang J T, Tan L J.Uptake and transformation of carbon, nitrogen and phosphorus in the co-culture system of algae and bacteria in seawater.Acta Ecologica Sinica,2017,37(14):4843- 4851.
猜你喜歡
小讀者(2021年2期)2021-03-29 05:03:48
新世紀智能(數學備考)(2020年11期)2021-01-04 00:38:16
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
中國外匯(2019年17期)2019-11-16 09:31:14
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
現代企業(2015年1期)2015-02-28 18:43:18
新高考·高一物理(2014年1期)2014-09-18 01:26:07
