高寒沙區生物土壤結皮覆蓋土壤碳通量對模擬降水的響應
2017-09-01 00:42:26賈曉紅楊占武楊德福趙雪彬
生態學報 2017年13期
關鍵詞:生物
辜 晨,賈曉紅,*,吳 波,成 龍,楊占武,楊德福,趙雪彬
1 中國林業科學研究院荒漠化研究所,北京 1000912 青海省農林科學院,西寧 8100163 青海共和荒漠生態系統國家定位觀測研究站,共和 813000
高寒沙區生物土壤結皮覆蓋土壤碳通量對模擬降水的響應
辜 晨1,賈曉紅1,*,吳 波1,成 龍1,楊占武2,楊德福3,趙雪彬3
1 中國林業科學研究院荒漠化研究所,北京 1000912 青海省農林科學院,西寧 8100163 青海共和荒漠生態系統國家定位觀測研究站,共和 813000
生物土壤結皮作為干旱半干旱地區重要的地表覆蓋類型和景觀特征之一,其自身具備的光合與呼吸能力對荒漠生態系統地表與大氣界面中的碳交換與循環產生了重要影響。水分是干旱半干旱地區許多生態過程中的主要限制因子,能夠影響生物土壤結皮的光合與呼吸過程,進而影響生物土壤結皮覆蓋土壤的碳通量規律。針對高寒沙區蘚類和藻類結皮為主的生物土壤結皮覆蓋土壤,設置了1、2、5、10mm以及0(對照)的模擬降水梯度,利用LI- 8100土壤碳通量測定系統,對模擬降水后結皮覆蓋土壤的碳通量進行測定,以探討不同結皮種類和不同強度降水對碳通量的影響。結果表明:(1)降水對生物土壤結皮覆蓋土壤的凈碳通量、暗呼吸均有激發作用,使碳通量在極短時間內到達峰值且與對照差異顯著,但各降水量之間差異不顯著。兩種不同結皮覆蓋類型相比,蘚類結皮覆蓋土壤在降水后的碳通量峰值和受降水激發的有效時間均顯著高于藻類結皮。(2)兩種結皮覆蓋土壤在模擬降水后的48h累積碳釋放隨降水量的增加而增加且與對照差異顯著。同時蘚類結皮覆蓋土壤累計碳釋放顯著高于藻類結皮覆蓋土壤。(3)兩種生物土壤結皮覆蓋土壤的碳通量和土壤水分體現出顯著的相關性,凈碳通量和暗呼吸均隨水分的增加而增加。因此,在降水條件下生物土壤結皮覆蓋土壤表現出明顯的碳源效應,其碳通量以及碳釋放量都有顯著的改變,在研究干旱半干旱地區碳交換規律時應該考慮不同生物結皮的覆蓋和降水事件的影響。
高寒沙區;碳通量;生物土壤結皮;模擬降水
生物土壤結皮廣泛分布于干旱半干旱地區,結構上主要由兩個部分組成,上層為微生物和隱花植物例如藻類、蘚類、地衣等,下層為覆蓋的土壤顆粒,二者通過菌絲、假根和分泌物等的緊密膠結成為一個整體,是具有代謝活性的特殊地表覆蓋層。在受非生物因素調控和脅迫的荒漠生態系統中,生物土壤結皮作為荒漠生態系統常見的地表覆被類型,是荒漠生態系統重要的組成部分和景觀特征之一,廣泛分布于干旱、半干旱區,地表覆蓋度到達60%以上甚至更高[1-2]。在荒漠景觀地區有機和無機成分的循環與交流中,生物土壤結皮扮演著聯系、指示和影響的角色,對許多生態過程產生直接或間接的影響。作為具有光合固碳活性的土壤與大氣系統界面層,生物土壤結皮中的藻類和蘚類植物以及下層土壤能夠通過呼吸作用釋放CO2,同時在有光條件下開始光合固碳過程,固定空氣中的CO2,釋放出氧氣。因此生物土壤結皮是干旱、半干旱地區荒漠系統碳通量的重要影響因素[2-4]。
在干旱半干旱區荒漠生態系統中,水分是主要的調控和限制因子,能夠直接影響生物土壤結皮的生理活動[5]。生物土壤結皮的重要組分藻類、地衣及蘚類植物作為變水植物對水分的變化尤其敏感,在頻繁多變的水分條件下有著很強的適應能力,缺水時自身開始脫水進入休眠狀態,而在水分條件適宜的情況下其生理活性才會逐漸恢復并顯現。因此生物土壤結皮的碳通量和碳釋放量受水分影響較為明顯。受全球氣候變化影響,未來干旱半干旱地區的降水格局將產生變化,降水頻率和幅度都將增加,這將導致許多自然資源和地表覆蓋類型產生巨大的空間異質性,地表植被的分布不均和自然資源的時空變異使我們在評估生物土壤結皮的碳源匯角色時增加了不確定性[6-9]。
目前國外有關研究集中在極地和苔原、科羅拉多高原、熱帶荒漠、亞熱帶草原、地中海中歐草原等,國內主要集中在黃土高原、騰格里沙漠、古爾班通古特沙漠、毛烏素沙地、科爾沁沙地等地。生物土壤結皮在降水影響下的碳通量交換規律的研究相對較少[4,10],在高寒沙區條件下進行的相關研究更是略顯不足。本研究在青海省共和盆地中西部典型高寒沙區人工植被恢復區,進行不同水平模擬降水下生物土壤結皮覆蓋土壤的凈碳通量(NCF)、呼吸速率(DR)、光合速率(Pn)以及相關環境因子進行動態監測,探尋生物土壤結皮覆蓋土壤碳通量對不同水平降水的響應規律,揭示了土壤碳通量與降水、土壤水分之間的關系,以期能夠為該地區生物土壤結皮的碳交換研究做出參考,也為整個高寒干旱半干旱地區的生物研究提供有益的背景資料。
1 材料與方法
1.1 研究區自然概況
研究區位于青海省林業廳下屬沙珠玉鄉治沙實驗站(98°45′—101°30′E,35°03′—35°40′N)。該實驗站地處共和盆地中西部,位于青海南山以南,是青海省荒漠化土地集中分布區之一,研究區總面積為5.28×102km2,沙地面積為1.74×102km2,海拔為2600—3400m。年均氣溫為2.4 ℃,1月平均氣溫為-12.5℃,7月平均氣溫為14.6 ℃。年均降水量僅為246.3mm,主要集中生長季,年內分配不均。降水多發生在夜間,歷時短且降水量少。年均潛在蒸發量為1716.7mm。
沙珠玉地區海拔較高,氣溫度較低,太陽輻射較強,水資源稀缺,生態環境脆弱;近幾年來,草場和林地被破壞的情況增多,使得該區域環境更加惡化,土地沙漠化程度逐年提高。從上世紀下半葉開始,沙珠玉地區逐漸開始對荒漠化土地進行有效治理,通過人工植被培育,減緩甚至遏制了沙漠化發展趨勢,穩固了沙丘,形成了具有一定規模的人工植被恢復區。本研究選擇1977年的植被恢復區為實驗樣地,該樣地位于固定沙丘的頂部,樣地內生物結皮發育良好,且分布廣泛,主要有藻結皮、蘚類結皮2種類型。土壤為棕鈣土和栗鈣土,非地帶性土壤為草甸土、鹽土和風沙土。研究區內的植被種類較少,群落結構簡單。自然植被中多為草本群落,如短花針茅(StipabrevifloraGriseb)和芨芨草(Achnatherumsplendens(Trin.)Nevski )。人工植被中,喬木主要有青楊(PopuluscathayanaRehd.)、河北楊(PopulushopeiensisHu et Chow)和小葉楊(PopulussimoniiCarr.),灌木主要有檸條錦雞兒(CaraganakorshinskiiKom.)、沙棘(HippophaerhamnoidesLinn.)、檉柳(TamarixchinensisLour.)等。藻結皮主要包含4類藻種,分別為漂浮克里藻(Klebsormidiumfluitans)、微小色球藻(Chroococcusminutus)、窩形席藻(Phormidicumfoveolarum)和溪生須藻(Homoeothrixfluviatillis)[11]。對該樣地內的結皮蓋度、厚度、結皮生物量等指標進行調查。由于結皮無法用烘干法測干物質的生物量,因此在本研究中藻結皮、蘚結皮的生物量均測定葉綠素a含量,用來表示其生物量。測定方法參照蘇延桂等的方法[12]。采集生物土壤結皮樣本放于土壤盒中(直徑4.5cm,表面積約15.9cm2),在實驗室中測定生物結皮葉綠素a含量。
用蒸餾水沖洗結皮樣本后放入研缽中同1.5mL乙醇(濃度98%)和少量石英砂、碳酸鈣進行研磨。完全研磨后,再加入7mL相同濃度乙醇進行2次各30min的離心(4℃,14000r/min)。完成后把提取液轉移至25mL容量瓶中,利用相同濃度的乙醇定容,之后將樣品放于25mL試管中。葉綠素a用紫外分光光度計(UV- 2450; Shimadzu, Kyoto, Japan)測定,測定使用波長為649nm、665nm。計算使用的公式如下:
Chla=(13.95×A665-6.88×A649)×25/15.9
(1)
式中,Chla為葉綠素a含量(mg/cm2),A665、A649分別表示波長為649nm、665nm時吸光值A,25表示稀釋體積(mL),15.9表示結皮的垂直投影面積(cm2)。為了防止葉綠素a的降解,樣品研磨、提取、測定均在相對黑暗的環境中進行。
實驗使用生物結皮樣品的特征如表1所示。

表1 生物結皮概況
生物結皮生物量用葉綠素a含量表示
1.2 研究方法
1.2.1 樣地布置
在研究區內坡度小于5°的叢間地上選擇發育良好的以蘚類結皮、藻類結皮為主的生物土壤結皮覆蓋土壤為研究樣地,保證結皮覆蓋度達到90%,結構完整,種類統一。
對于蘚類、藻類結皮,分別設置4個10m×10m的樣方,每個樣方之間相距5m以上。在4個大樣方中隨機選擇3個,設置4個50cm×50cm的小樣方,每個小樣方內分別放置1個直徑為20cm的土壤環,進行1mm、2mm、5mm、10mm的模擬降水處理,即P1、P2、P5、P10,剩余一個大樣方設置3個小樣方,進行0mm(對照,CK)降水。土壤環使用PVC材料自制,在測定之前3d設置于樣地內,以便內外充分平衡,放置時保證樣環內結皮完整。整個樣方設置分為2種結皮處理,5種降水處理,3個重復,共30個測定基座。
1.2.2 土壤碳通量測定
參考當地多年降水資料,雨季集中在每年6月—9月,降水基本在10mm以下以小降水事件為主。因此實驗于2015年9月進行,設置1、2、5、10mm4種降水處理。在晴朗的天氣條件下,每天日出之后,約7點左右開始人工模擬降水,使用噴壺噴灑,保證水流速度,使水滴均勻撒在地表,并保證不能形成地表徑流,控制整個降雨過程在3min之內,對不同的降水量使用完全相同的操作方法。在降水后的0、1、2、4、6、8、12、24、48h進行凈碳通量(NCF)和呼吸速率(DR)的測定。測定時間分別為第一日07:00、08:00、09:00、11:00、13:00、15:00、19:00、第2日07:00、第3日07:00,每次測定都在天亮時進行。碳通量的測定使用Li- 8100土壤碳通量系統,配以便攜測量室8100- 103以及長期監測室8100- 104C。凈碳通量即為在光照條件下(使用長期檢測室8100- 104C,該氣室使用透明材料制作,透光率>99%),測定通過空氣-土壤界面層的CO2通量值,為生物土壤結皮的光合作用、呼吸作用與下層土壤的呼吸作用的總和。呼吸速率即為在遮光條件下(使用便攜測量室8100- 103,該氣室外罩使用金屬材質,保證測定在完全黑暗條件下進行),測定通過界面的CO2通量值,為生物土壤結皮呼吸作用與亞表層土壤的呼吸作用的總和。使用Li- 8100測定碳通量時,設定每次測定90s,儀器自動對變化曲線進行擬合得出斜率,即為通量值。根據NCF和DR即可計算得到光合速率(Pn)。
Pn=DR -NCF
(1)
式中,Pn為正并且數值越大時,表示光合速率越高,碳吸收能力越強。
土壤體積含水量(SVWC) 采用輔助傳感器測定,測量土壤深度為5cm,與土壤碳通量測定同時進行。
1.2.3 數據處理與分析
對模擬降水后的蘚類覆蓋土壤和藻類覆蓋土壤的凈碳通量、呼吸速率和光合速率進行了分析,同時對模擬降水后48h內的碳通量值進行了平均得到平均碳通量,并且對兩種不同結皮覆蓋土壤48h內的碳通量變化規律曲線進行了積分,得到累積碳量。
數據分析和圖表繪制分別在SPSS、Origin和Excel中進行。使用重復測量方差分析進行土壤覆蓋類型對碳通量影響效果的分析。雙因素方差分析用來對比不同降水處理之間的碳通量規律間的區別。回歸方程用來模擬碳通量和土壤水分溫度之間的關系。回歸方程的顯著性檢驗通過方差分析檢驗。
2 結果與分析
2.1 生物土壤結皮覆蓋土壤碳通量對模擬降水的響應
2.1.1 生物土壤結皮覆蓋土壤凈碳通量的動態變化
降水量對生物土壤結皮覆蓋土壤的NCF有極顯著影響(P<0.01)。降雨前兩種生物土壤結皮覆蓋土壤的NCF分別是1.17,2.51μmol m-2s-1。降水后二者均表現出大量的碳釋放,其NCF在短時間內到達峰值,然后隨時間逐漸下降,在1h左右重新上升,并在6h時到達又一峰值后再次開始下降,呈現出先快后慢,先上升后下降的趨勢(圖1)。蘚類結皮在24h左右回到原始水平,而藻類結皮在12h左右基本回到原始水平。
對于蘚類結皮,五種處理下的NCF的最大值分別為0.33、5.16、9.72、10.61、9.41μmol m-2s-1,即P5> P2>P10>P1>CK。藻類結皮覆蓋土壤的NCF最大值分別為0.29、2.61、3.03、4.40、3.34μmol m-2s-1。NCF在兩種結皮類型中差異顯著(P<0.01),而4種降水處理與CK對比差異顯著(P<0.01),降水量之間對比不顯著(P>0.01)(表2)。
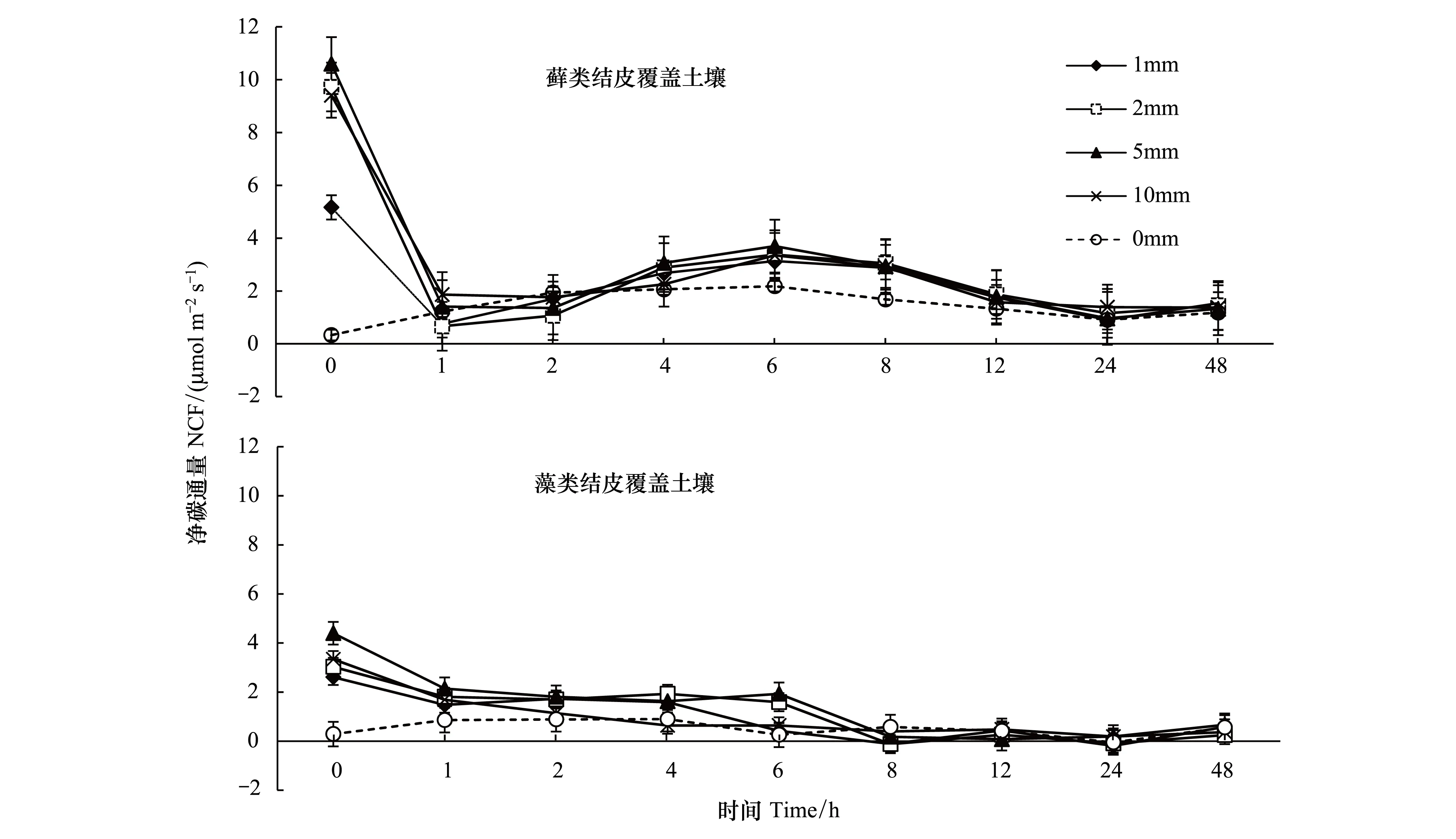
圖1 生物土壤結皮覆蓋土壤凈碳通量對模擬降水的響應Fig.1 Response of net carbon flux in biological-soil crusted soil to simulated precipitation
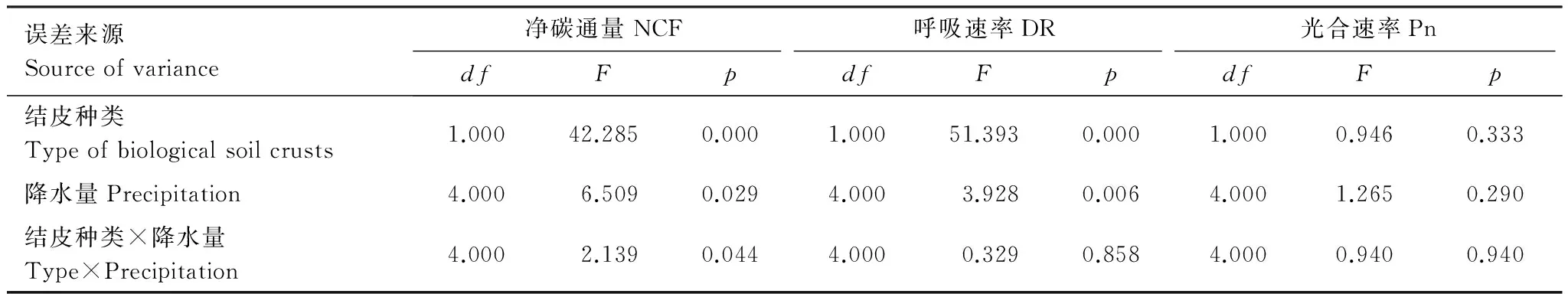
誤差來源Sourceofvariance凈碳通量NCF呼吸速率DR光合速率PndfFpdfFpdfFp結皮種類Typeofbiologicalsoilcrusts1.00042.2850.0001.00051.3930.0001.0000.9460.333降水量Precipitation4.0006.5090.0294.0003.9280.0064.0001.2650.290結皮種類×降水量Type×Precipitation4.0002.1390.0444.0000.3290.8584.0000.9400.940
2.1.2 生物土壤結皮覆蓋土壤呼吸速率的動態變化
降水量對生物土壤結皮覆蓋土壤的DR也有著顯著影響(P<0.01)。降雨前兩種生物土壤結皮覆蓋土壤的DR分別是1.16,0.79μmol m-2s-1。降水后兩種生物土壤結皮覆蓋土壤的DR十分迅速的達到最高值,然后隨時間逐漸下降。在峰值時蘚類結皮DR分別為0.95、3.66、4.78、5.25、8.93μmol m-2s-1,表現為P10>P5>P2>P1>CK,P10顯著高于其他處理,P5、P2和P1顯著高于CK。蘚類結皮的DR在24h后回到正常水平。
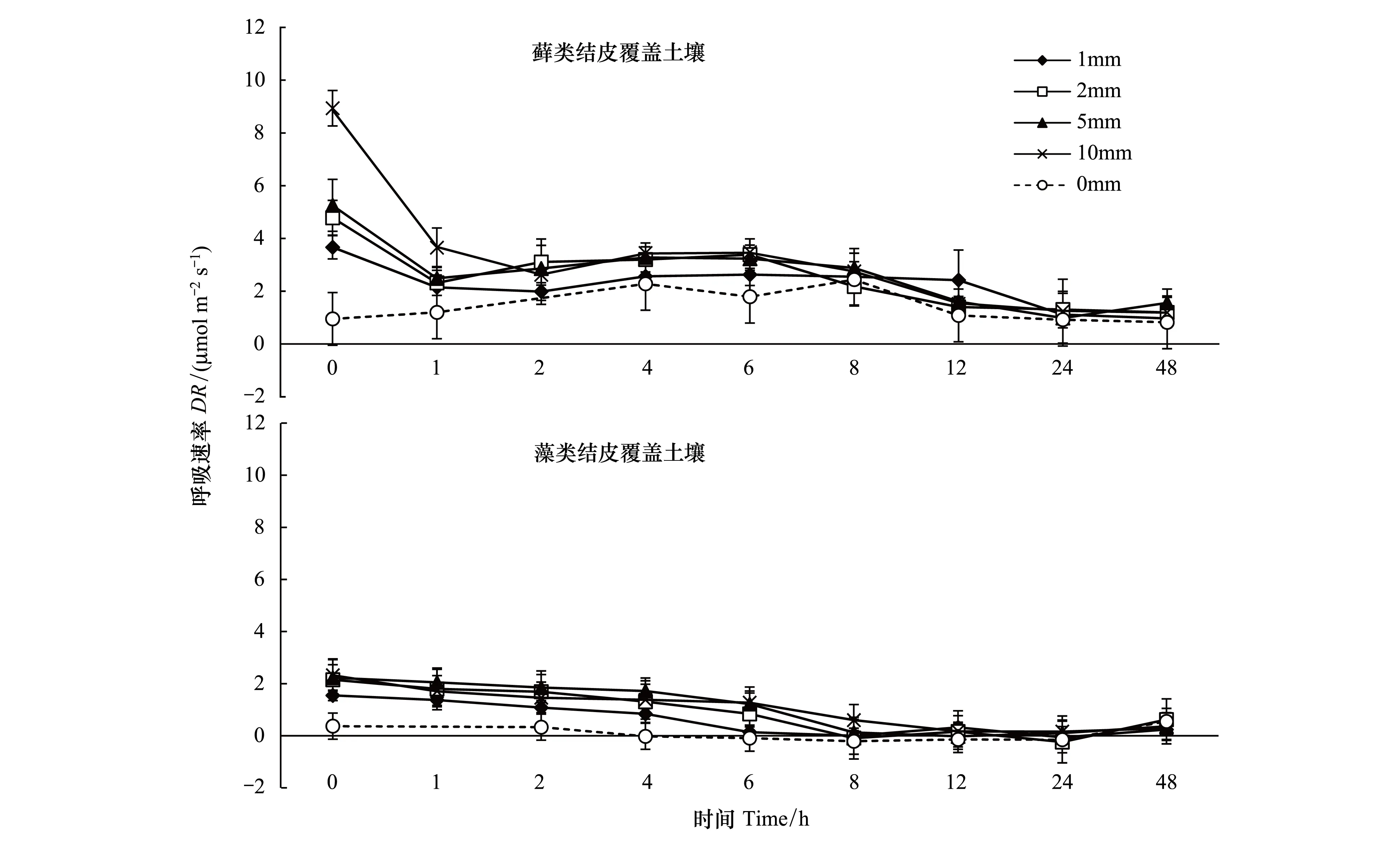
圖2 生物土壤結皮覆蓋土壤呼吸速率對模擬降水的響應Fig.2 Response of respiration rate in biological-soil crusted soil to simulated precipitation
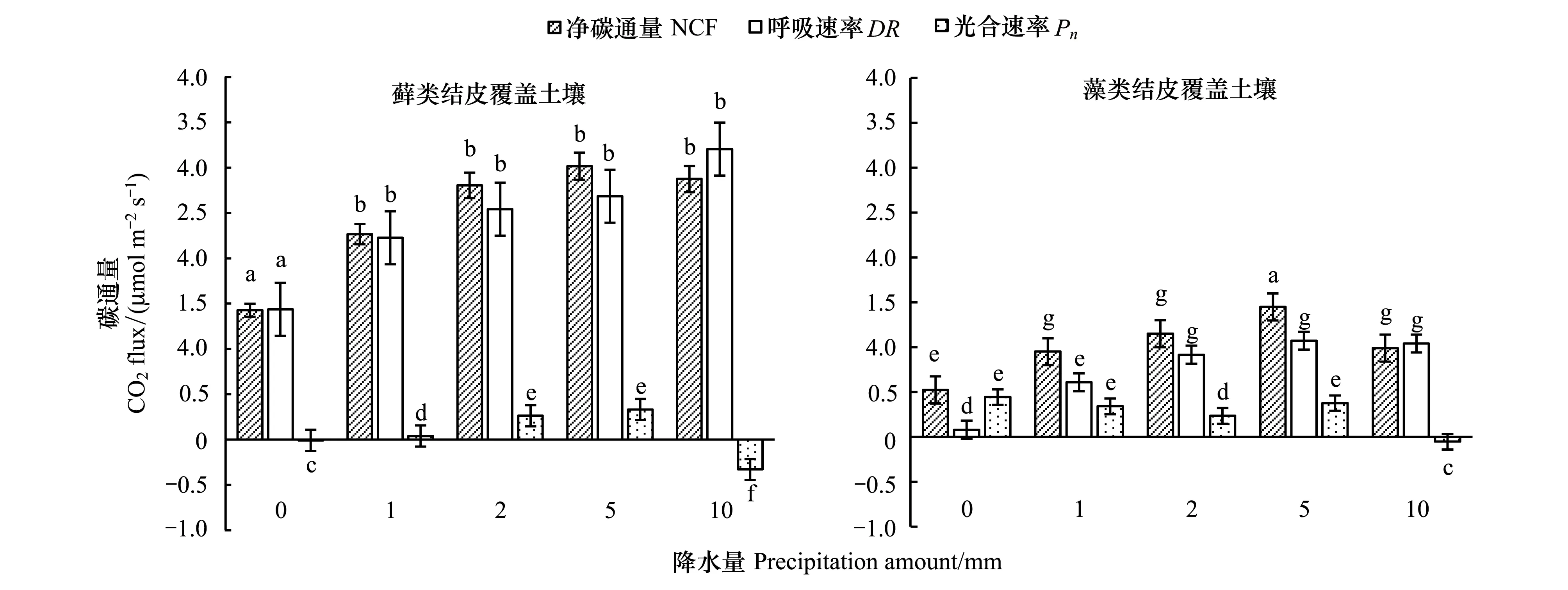
圖3 48h內平均碳通量和降水的關系Fig.3 The relationships between the average CO2 flux and simulated rainfall amount over the first 48 h (AVR)a,b,c,d,e,f代表不同的顯著性水平
藻類結皮覆蓋土壤的DR最大值分別為0.37、1.55、2.15、2.23、2.32μmol m-2s-1,表現為P5> P10> P2> P1,均顯著高于CK,不同降水處理間有顯著差異(P<0.01)。對于藻結皮,除P10外其他降水處理均在24h左右回到正常水平。兩種結皮相對比,蘚類結皮覆蓋土壤的DR受到模擬降水的影響較明顯,且蘚類結皮的DR最大值高于藻類結皮。
隨著降水量的增大,兩種結皮覆蓋土壤的48h內平均碳通量也隨之增加,蘚類結皮覆蓋土壤的NCF分別為1.42、2.66、2.80、3.01、2.87μmol m-2s-1,DR分別為1.43、2.22、2.54、2.68、3.20μmol m-2s-1。
2.1.3 生物土壤結皮光合速率的動態變化
根據圖4所示,降水后蘚類結皮的Pn在降水后總體呈現出逐漸上升的趨勢。初始值分別為-1.50、-4.94、-5.35、-0.47、0.62μmol m-2s-1,在2h左右到達光合速率最大值,此時為0.26、2.04、1.50、0.86、0μmol m-2s-1,并在12h之后基本保持平穩。在整個測定過程中,蘚類結皮的4種降水處理與對照之間差異顯著(P<0.01),而每個降水量之間的差異不顯著(P>0.01)。
藻類結皮Pn的變化趨勢較之蘚類表現的并不明顯。藻類結皮在開始時分別為-1.06、-0.88、-2.17、-1.02、0.08μmol m-2s-1,其中P10隨著時間其Pn逐漸提高并在4h至6h左右到達最大值,并在降水后24h后恢復正常值。其他降水處理并沒有表現出明顯的光合速率增加趨勢。蘚類結皮光合速率表現出的最高值為藻類結皮的2.76倍,而藻類結皮的光合速率也比蘚類結皮更早回到正常水平。
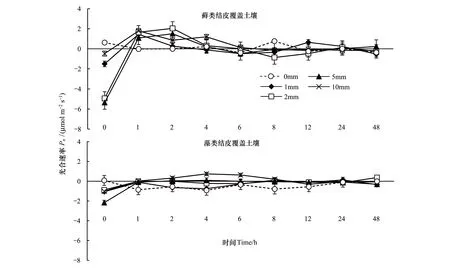
圖4 光合速率對模擬降水的響應Fig.4 Response of photosynthetic rate in biological-soil crusts to simulated precipitation
2.2 模擬降水對生物土壤結皮碳累積釋放量的影響
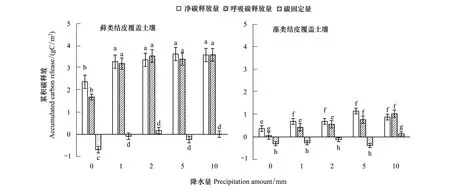
圖5 模擬降水與累積碳釋放量的關系Fig.5 The relationships between the total accumulated carbon relaease and simulated rainfall amounta,b,c,d,e,f代表不同的顯著性水平
如圖5所示,降水后,蘚類結皮48h內累計碳凈釋放量(有光條件下NCF累計量)隨降雨量變化相對于CK明顯增加,分別為1.68、3.19、3.54、3.39、3.59 gC/m2。表現為P10> P2> P5> P1,各降水處理之間差異不顯著(P>0.01)。藻類結皮分別為0.37、0.69、0.69、1.15、0.88 gC/m2。各個降水處理之間差異不顯著(P>0.01),但是與CK之間差異顯著(P<0.01)。蘚類結皮覆蓋土壤凈累積碳釋放明顯多于藻類結皮,分別為藻類結皮的3.78、4.58、5.13、2.93、4.05倍。蘚類呼吸累積碳釋放量(DR累計量)同樣隨降雨量的增大而增大,分別為1.68、3.20、3.54、3.40、3.59 gC/m2,表現為P5> P10> P2> P1,差異不顯著(表3)。藻類呼吸累積釋放量分別為0.06,0.43,0.56,0.77,1.03 gC/m2,隨著降雨量的增加而增加。
2.3 模擬降水后生物土壤結皮覆蓋土壤碳通量和土壤水分、溫度的關系
為了分析土壤水分與碳通量的關系,使用對數方程分別對蘚類結皮P5和藻結皮P10降水處理下的NCF與DR進行擬合。擬合方程分別為y=0.485ln(x) + 3.1885(蘚類結皮覆蓋土壤呼吸速率,P<0.01),y=2.2226ln(x)+6.7728(蘚類結皮覆蓋土壤凈碳通量,P<0.01),y=33.225ln(x) + 36.684(藻類結皮覆蓋土壤呼吸速率,P<0.01)以及y=40.833ln(x) + 44.08(藻類結皮覆蓋土壤凈碳通量,P<0.01)。兩種結皮覆蓋土壤的NCF和DR均表現出了隨水分增大的趨勢(圖6,圖7)。
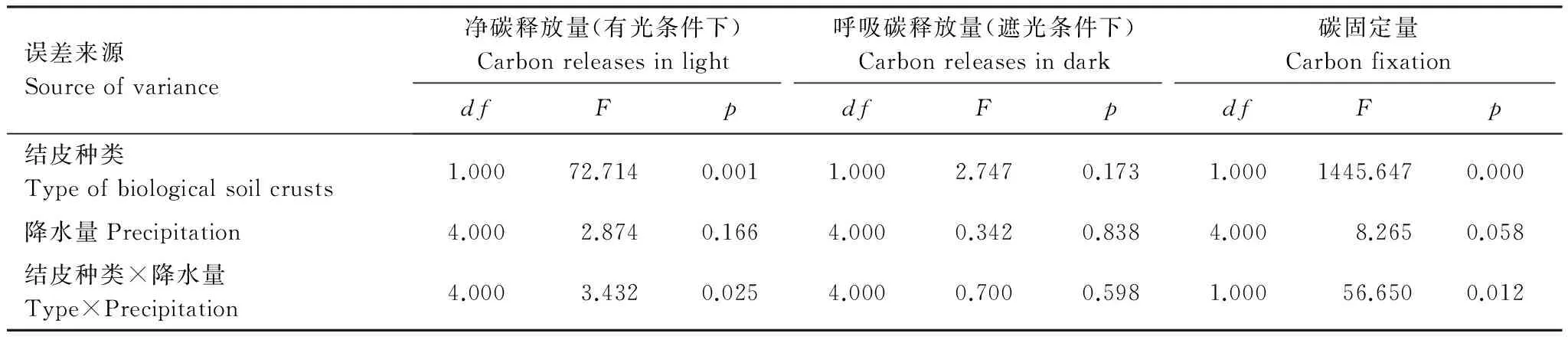
表3 結皮類型和降水量對48h內累計碳釋放的影響

圖6 5mm降水處理下蘚類結皮覆蓋土壤凈碳通量和呼吸速率與土壤水分的關系Fig.6 The relationships between the NCF, DR and SVWC in moss-crusted soil under 5mm precipitation

圖7 10mm降水處理下藻類結皮覆蓋土壤凈碳通量和呼吸速率與土壤水分的關系Fig.7 The relationships between the NCF, DR and SVWC in algea-crusted soil
對蘚類和藻類為主結皮覆蓋土壤的NCF和DR與土壤溫度的相關分析表明,兩類結皮覆蓋土壤的NCF和DR均與溫度無明顯的相關關系(圖8,圖9)。
3 討論
在沙漠中,土壤有效水分是決定植被種類,植物生理活性和其他相關生理過程的關鍵因素[13-14]。對于生物土壤結皮,水分能夠直接影響其光合與呼吸作用。而土壤水分與降水直接相關,在干旱半干旱地區土壤中的水分絕大多數來自于降水。本研究表明降水會刺激蘚類結皮和藻類結皮的碳釋放,并且隨著降水量的增加,碳釋放量也有一定程度的增加,這與前人的研究一致[15-17]。
研究中,生物土壤結皮覆蓋土壤的凈碳通量在模擬降水后迅速到達峰值然后下降,1h至2h后再次上升,之后隨著時間的推移逐漸下降至自然水平。對于這種現象,可能有以下幾個原因。首先,降水前土壤水分處于干旱狀態,土壤空隙較大,降雨后水分滲入土壤空隙將土壤空隙中的CO2排出,這就增加了土壤碳排放[18-19];其次,在干燥條件下土壤中可能累積一定的活性碳和呼吸底物,水分的增加可能影響了土壤微生物活性從而增加了土壤碳排放[20]。隨著時間推移這一部分的碳排放會迅速減少,生物土壤結皮因為之前處于干燥狀態而對水分的響應存在一定的延遲與滯后,在一段時間后才能增加呼吸速率和光合速率,所以在圖中表現出了降水一段時間后碳通量的二次上升現象。兩種結皮的凈碳通量、呼吸速率、光合速率以及累積碳釋放量在降水后均有明顯的增加,但在數值上有較大差異,蘚類結皮的凈碳通量、呼吸速率和光合速率更高。這不僅是因為兩種結皮自身存在著生理活性的差異,蘚類結皮擁有著比藻類結皮更強的呼吸與光合能力,除此之外還存在其他影響因素。一方面,蘚類結皮能夠保持較長時間的水分有效性,因此能夠在降水后長時間內保持濕潤,減緩水分蒸發和下滲,保持自身活性。藻類結皮的保水持水能力較差,在降水事件,尤其是小降水事件后的20h之內回到原始水平。另一方面,兩種結皮覆蓋土壤的性狀也有所區別,蘚類結皮覆蓋土壤孔隙度較大,養分含量相對較高,而藻類結皮覆蓋土壤的土壤穩定性較差、粘粒含量很少。兩種結皮的覆蓋對水分小循環的影響和對土壤性狀的改變所表現出的差異也符合生物土壤結皮從藻類到蘚類、從低級到高級的演替規律[21]。
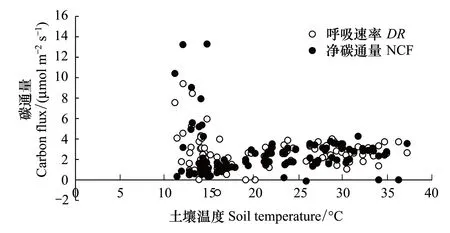
圖8 土壤溫度與蘚類結皮覆蓋土壤碳通量的關系 Fig.8 The relationships between the CO2 flux and the soil temperatures in moss-crusted soil
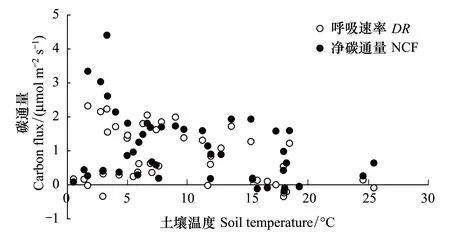
圖9 土壤溫度與藻類結皮覆蓋土壤碳通量的關系 Fig.9 The relationships between the CO2 flux and the soil temperatures in algea-crusted Soil
雖然有研究認為生物土壤結皮能夠通過光合作用固定大氣中的CO2,是干旱半干旱區潛在的碳匯,但本次研究結果表明模擬降水后生物土壤結皮覆蓋土壤扮演著碳源的角色。兩種生物土壤結皮覆蓋土壤在模擬降水后的凈碳通量都表現為碳釋放,這個過程中沒有觀測到生物土壤結皮的碳吸收,說明結皮覆蓋土壤在模擬降水的刺激下始終保持著碳源效應。這與前人的研究具有相同之處[22,23]。這可能是由于生物土壤結皮的光合速率較低,不能抵消土壤呼吸所導致的。研究中發現蘚類結皮的光合速率最大達到2.04μmol m-2s-1,下降后基本在0至-1.5μmol m-2s-1之間波動。藻類為優勢種的結皮中光合速率通常為1 μmol m-2s-1或更少[24-26]。研究地區頻繁的干旱脅迫導致生物土壤結皮經常保持在休眠狀態,而在干旱休眠狀態時的生物土壤結皮基本不具備光合能力。休眠中的生物土壤結皮在有了適當的水分時并不能立即完全表現出其光合生理活性,需要一定時間響應。所以這有可能是實驗選擇的小降雨量引起的生物土壤結皮光合速率尤其是藻類結皮覆蓋土壤光合速率較低且變化不明顯的原因。
本研究中雖然根據自然降水情況設置了模擬降水,但是沒有連續長時間的在自然條件下監測,同時也沒有考慮某些特殊環境條件,例如冰雪融水、極端降水等,生物土壤結皮在干旱區碳循環中的功能以及碳循環中的碳源匯角色有待深入研究。
4 結論
(1)降水對生物土壤結皮覆蓋土壤的凈碳通量和呼吸有極顯著激發作用。兩種生物土壤結皮覆蓋土壤的凈碳通量、呼吸、平均碳通量和累積碳釋放均隨著降水的增加而增加,各降水處理相比于對照差異顯著。兩種結皮覆蓋土壤的光合速率在降水后也有增加的趨勢,其中蘚類結皮覆蓋土壤的光合速率增加趨勢較明顯,各降水處理與對照差異顯著。藻類結皮覆蓋土壤的光合速率變化不明顯,只有在高降水量處理下才有較明顯的增加。
(2)兩種生物土壤結皮覆蓋土壤在模擬降水后的凈碳通量、呼吸和光合速率變化規律差異顯著。蘚類結皮覆蓋土壤在模擬降水后的凈碳通量、呼吸和光合速率的極值顯著高于藻類結皮,同時蘚類結皮覆蓋土壤碳通量受降水激發的有效時間也高于藻類結皮覆蓋土壤。兩種生物土壤結皮覆蓋土壤在模擬降水后48h內的累積碳釋放隨著降水量的增加而增加,相對于對照差異顯著,且蘚類結皮覆蓋土壤的48h累計碳釋放顯著高于藻類結皮。
(3)兩種生物土壤結皮的凈碳通量和呼吸速率與土壤水分有著較明顯的相關性,表現出隨土壤水分增大而增大的趨勢,但與土壤溫度沒有明顯的相關關系。
[1] Su Y G, Wu L, Zhou Z B, Liu Y B, Zhang Y M. Carbon flux in deserts depends on soil cover type: a case study in the Gurbantunggute Desert, North China. Soil Biology and Biochemistry, 2013, 58: 332- 340.
[2] Belnap J, Lange O L. Biological Soil Crusts: Structure, Function, and Management. Berlin: Springer-Verlag, 2001: 239- 257.
[3] Wassmann R, Neue H U, Lantin R S, Aduna J B, Alberto M C R, Andales M J, Tan M J, van der Gon H A C D, Hoffmann H, Papen H, Rennenberg H, Seiler W. Temporal patterns of methane emissions from wetland rice fields treated by different modes of N application. Journal of Geophysical Research-Atmospheres, 1994, 991(D8): 16457- 16462.
[4] 李新榮, 賈玉奎, 龍利群, 王新平, 張景光. 干旱半干旱地區土壤微生物結皮的生態學意義及若干研究進展. 中國沙漠, 2001, 21(1): 4- 11.
[5] Grote E E, Belnap J, Housman D C, Sparks J P. Carbon exchange in biological soil crust communities under differential temperatures and soil water contents: implications for global change. Global Change Biology, 2010, 16(10): 2763- 2774.
[6] Schlesinger W H, Raikes J A, Hartley A E, Cross A F. On the spatial pattern of soil nutrients in desert ecosystems. Ecology, 1996, 77(2): 364- 374.
[7] Jasoni R L, Smith S D, Arnone J A III. Net ecosystem CO2exchange in Mojave Desert shrub lands during the eighth year of exposure to elevated CO2. Global Change Biology, 2005, 11(5): 749- 756.
[8] Wohlfahrt G, Fenstermaker L F, Arnone J A III. Large annual net ecosystem CO2uptake of a Mojave Desert ecosystem. Global Change Biology, 2008, 14(7): 1475- 1487.
[9] Xie J X, Li Y, Zhai C X, Li C H, Lan Z D. CO2absorption by alkaline soils and its implication to the global carbon cycle. Environmental Geology, 2009, 56(5): 953- 961.
[10] José J J S, Bravo C R. CO2exchange in soil algal crusts occurring in the trachypogon savannas of the Orinoco Llanos, Venezuela. Plant and Soil, 1991, 135(2): 233- 244.
[11] 陳正宏. 高寒草甸沙化過程中生物結皮藻類組成及分布的變化[D]. 蘭州: 蘭州理工大學, 2009.
[12] 蘇延桂, 李新榮, 陳應武, 崔艷, 魯艷. 溫度和CO2濃度升高對荒漠藻結皮光合作用的影響. 應用生態學報, 2010, 21(9): 2217- 2222.
[13] Song W M, Chen S P, Wu B, Zhu Y J, Zhou Y D, Li Y H, Cao Y L, Lu Q, Lin G H. Vegetation cover and rain timing co-regulate the responses of soil CO2efflux to rain increase in an arid desert ecosystem. Soil Biology and Biochemistry, 2012, 49: 114- 123.
[14] Sponseller R A. Precipitation pulses and soil CO2flux in a Sonoran Desert ecosystem. Global Change Biology, 2007, 13(2): 426- 436.
[15] Housman D C, Grote E E, Belnap J. Annual CO2flux from a biological soil crust system on the Colorado Plateau: effects of increased temperature and summer precipitation // Ecological Society of America Annual Meeting Abstracts. 2007.
[16] 趙蓉, 李小軍, 趙洋, 楊昊天, 陳棟. 固沙植被區兩類結皮斑塊土壤呼吸對不同頻率干濕交替的響應. 生態學雜志, 2015, 34(1): 138- 144.
[17] 趙洋, 齊欣林, 陳永樂, 張志山, 賈榮亮, 胡宜剛, 張鵬. 極端降雨事件對不同類型生物土壤結皮覆蓋土壤碳釋放的影響. 中國沙漠, 2013, 33(2): 543- 548.
[18] Anderson J M. Carbon dioxide evolution from two temperate, deciduous woodland soils. Journal of Applied Ecology, 1973, 10(2): 361- 378.
[19] Birch H F. The effect of soil drying on humus decomposition and nitrogen availability. Plant and Soil, 1958, 10(1): 9- 31.
[20] Gordon H, Haygarth P M, Bardgett R D. Drying and rewetting effects on soil microbial community composition and nutrient leaching. Soil Biology and Biochemistry, 2008, 40(2): 302- 311.
[21] Zaady E, Kuhn U, Wilske B, Sandoval-Soto L, Kesselmeier J. Patterns of CO2Exchange in biological soil crusts of successional age. Soil Biology and Biochemistry, 2000, 32(7): 959- 966.
[22] 吳林, 蘇延桂, 張元明. 模擬降水對古爾班通古特沙漠生物結皮表觀土壤碳通量的影響. 生態學報, 2012, 32(13): 4103- 4113.
[23] 賈曉紅, 辜晨, 吳波,李元壽,成龍,李新榮. 干旱沙區生物土壤結皮覆蓋土壤CO2通量對脈沖式降雨的響應. 中國沙漠, 2016(02):423- 432.
[24] Lange O L, Belnap J, Reichenberge H. Photosynthesis of the cyanobacterial soil-crust lichenCollematenaxfrom arid lands in southern Utah, USA: role of water content on light and temperature responses of CO2exchange. Functional Ecology, 1998, 12(2): 195- 202.
[25] Brostoff W N, Sharifi M R, Rundel P W. Photosynthesis of cryptobiotic soil crusts in a seasonally inundated system of pans and dunes in the western Mojave Desert, CA: field studies. Flora-Morphology, Distribution, Functional Ecology of Plants, 2005, 200(6): 592- 600.
[26] Li X R, Zhou H Y, Wang X P, Zhu Y G, O′Conner P J. The effects of sand stabilization and revegetation on cryptogam species diversity and soil fertility in the Tengger Desert, Northern China. Plant and Soil, 2003, 251(2): 237- 245.
Effect of simulated precipitation on the carbon flux in biological-soil crusted soil in alpine sandy habitats
GU Chen1, JIA Xiaohong1,*, WU Bo1, CHENG Long1, YANG Zhanwu2, YANG Defu3, ZHAO Xuebin3
1 Institute of Desertification Studies, Chinese Academy of Forestry, Beijing 100091, China2 Qinghai Academiy of Agriculture and f Forestry Science, Xining 810016, China3 Qinghai Gonghe Desert Ecosystem Research Station, Gonghe 813000, China
Biological-soil crusts, a community of cyanobacteria, lichens, and mosses that live on the soil surface, are a critical part of land cover types in desert ecosystems. They play a significant role in many biogeochemical processes, contributing to soil fertility, stability, and vascular plant establishment. The photosynthetic and respirometric activity of biological-soil crusts can affect carbon fluxes and exchange in desert ecosystems. Water is the major limiting factor of ecosystem functions and processes in arid and semi-arid regions. In environments with dry climates, scarce precipitation, and water shortages, water can affect the photosynthesis and respiration of biological-soil crusts, leading to changes in carbon fluxes and exchange in the soil. In the present study, to determine the effect of biological-soil crusts on soil carbon fluxes under different amounts of precipitation, we used the two dominant soil crusts types found in the Qinghai-Tibet Plateau alpine sandy vegetated areas: moss and algae. Five different simulated precipitation amounts (1, 2, 5, 10, and 0 mm) were applied, and the carbon flux of soil covered by biological crusts was measured using Li- 8100 in the dark as dark respiration and in the light as the net carbon flux. The results show that the: (1) net carbon flux and dark respiration of biological-soil crusted soil were stimulated by simulated precipitation, and increased rapidly to their maximum levels. There was a significant difference between the precipitation treatments and control, but no significant difference between different treatments. The maximum carbon flux and stimulated available time of moss-crusted soil were much higher and longer, respectively, than those of algae-crusted soil; (2) total accumulated carbon release of two crusted soils after the addition of simulated precipitation increased with increasing precipitation, and there was a significant difference between carbon release in the precipitation treatments and the control. The total accumulated carbon release of moss-crusted soil was significantly more than that of algae-crusted soil; (3) carbon flux of two crusted soils and the soil volumetric water content showed a significant correlation. The effects of soil water on net carbon flux and dark respiration were stimulative. The results of the analysis indicated that the biological-crusted soil showed an effect of the carbon source after the addition of simulated precipitation. Precipitation can directly influence the carbon flux and release, and the response of the carbon flux in biological-soil crusted soil to simulated precipitation should be considered when studying carbon exchange in arid and semi-arid regions.
alpine sandy lands; carbon flux; biological soil crusts; simulated precipitation
國家自然科學基金(41471096,41371093,41171077);中央級公益性科研院所基本科研業務費項目(CAFYBB2016ZD010)
2016- 03- 22; 網絡出版日期:2017- 02- 23
10.5846/stxb201603220510
*通訊作者Corresponding author.E-mail: jiaxiaohong@caf.ac.cn
辜晨,賈曉紅,吳波,成龍,楊占武,楊德福,趙雪彬.高寒沙區生物土壤結皮覆蓋土壤碳通量對模擬降水的響應.生態學報,2017,37(13):4423- 4433.
Gu C, Jia X H, Wu B, Cheng L, Yang Z W, Yang D F, Zhao X B.Effect of simulated precipitation on the carbon flux in biological-soil crusted soil in alpine sandy habitats.Acta Ecologica Sinica,2017,37(13):4423- 4433.
猜你喜歡
天天愛科學(2022年9期)2022-09-15 01:12:54
天天愛科學(2022年4期)2022-05-23 12:41:48
當代水產(2022年3期)2022-04-26 14:26:56
科學大眾(2021年9期)2021-07-16 07:02:54
軍事文摘(2020年20期)2020-11-28 11:42:50
航空世界(2020年10期)2020-01-19 14:36:20
小學科學(學生版)(2018年3期)2018-04-18 12:34:19
知識經濟·中國直銷(2017年10期)2017-11-07 02:39:52
知識經濟·中國直銷(2017年3期)2017-04-16 03:08:01
文學少年(有聲彩繪)(2017年2期)2017-03-06 08:10:54
