不同氮水平下玉米苗期根系形態和氮吸收量的QTL定位
2017-09-03 09:34:50王祎湯繼華付延磊許恒譚金芳韓燕來
植物營養與肥料學報 2017年4期
關鍵詞:檢測
王祎,湯繼華,付延磊,許恒,譚金芳,2,韓燕來*
(1河南農業大學資源與環境學院,河南鄭州450002;2河南省糧食作物協同創新中心,河南鄭州450002;3省部共建小麥玉米國家重點實驗室,河南鄭州450002)
不同氮水平下玉米苗期根系形態和氮吸收量的QTL定位
王祎1,2,3,湯繼華3,付延磊1,許恒3,譚金芳1,2,韓燕來1,2,3*
(1河南農業大學資源與環境學院,河南鄭州450002;2河南省糧食作物協同創新中心,河南鄭州450002;3省部共建小麥玉米國家重點實驗室,河南鄭州450002)
【目的】玉米的根系形態與氮素吸收能力關系密切,利用單片段代換群體對玉米苗期根系形態相關性狀和植株氮吸收量進行QTL定位,可為進一步精細定位并克隆控制玉米低氮下優異根系形態和氮吸收的主效QTL奠定基礎。【方法】以氮效率差異顯著的兩親本許178和綜3構建的150個玉米單片段代換系(SSSL)群體作為研究材料,進行水培試驗。以Ca(NO3)2作為氮源,設置高氮(4mmol/L NO3–)和低氮(0.05mmol/L NO3
–)兩個處理,培養20d后分根、冠收獲植株,測定生物量和氮含量。通過WinRHIZO根系分析系統獲得根系的總根長、根表面積、根體積、根直徑和根尖數等指標。根據代換系與親本許178表型值的T-test結果,利用該群體SSR遺傳連鎖圖譜,在P≤0.001條件下定位所調查性狀的QTL。【結果】高氮條件下SSSL群體除了根直徑與總根長和根尖數沒有顯著相關性以外,其它各性狀之間均顯著或極顯著正相關,并且植株氮吸收量也與根系各性狀呈顯著或極顯著正相關;低氮條件下,除了根直徑以外,植株氮吸收量與其他根系性狀均呈極顯著正相關,并且地上部和根部氮累積量均與根表面積的相關性最大。在高氮條件下共檢測到102個QTL位點,包括40個根形態相關QTL、34個植株生物量QTL和28個氮吸收量QTL;在低氮條件下共檢測到85個QTL位點,包括47個根形態QTL、22個植株生物量QTL和16個氮吸收量QTL。所檢測到的根形態相關QTL與生物量和氮積累量QTL成簇存在,同一QTL區間多同時檢測到根形態QTL和氮吸收量QTL。高氮條件下,在代換系1428、1376、1282、1266和1473的代換區間上檢測到高氮特異的QTL簇,同時包括多個根形態和氮吸收量QTL,貢獻率從–43%到84%。低氮下,在代換系1419和1314的代換區間上同時檢測到低氮特異的多個根形態和氮吸收量QTL,貢獻率從–32%到55%。【結論】單片段代換系1419和1314所包含的代換片段bnlg182—bnlg2295和umc1013—umc2047檢測到多個低氮特異的QTL,并且這兩個區間在前人的研究中均有玉米氮效率相關QTL檢測到,說明該區間包含有玉米根系形態和氮吸收量的主效QTL,在玉米氮高效吸收中可能起重要作用。
玉米;單片段代換系;根系形態;QTL;低氮脅迫
玉米是全球最重要的多用途作物之一,廣泛應用于糧食、飼料、化工、生物能源等行業。為保證國家糧食安全,我國在《國家糧食安全中長期規劃綱要》中明確提出到2020年需要新增糧食1000億斤,其中需要新增玉米650億斤,占全國新增糧食的65%,因此玉米在我國未來糧食生產中的地位舉足輕重。我國玉米高產育種已取得了重大成就,超高產記錄不斷突破。不容忽視的是,目前的玉米高產記錄都是高肥高水條件下獲得,同樣為提高玉米產量,我國農田的氮肥投入量大、氮肥利用率低,不但嚴重制約我國玉米產量和農業經濟效益的進一步提高,而且成為導致河流和湖泊富營養化的重要因素之一[1]。因此,提高玉米的氮素利用率、培育氮高效品種,從而在適當降低施氮量的條件下維持較高產量水平,對實現農業可持續發展和環境保護意義重大[2]。
研究表明,在玉米籽粒形成中54.5%~60.6%的氮來自營養體的再運轉[3]。因此,植株前期氮吸收效率對于后期提高玉米籽粒產量至關重要。植物主要通過根系從外界吸收礦質營養,有效的根系形態對培育氮吸收高效的玉米基因型、減少氮淋失非常重要[4]。許多研究表明,增加根系體積,包括根干重、根長和根密度,可以提高玉米氮吸收能力和籽粒產量[5–6]。玉米的根系形態受土壤有效氮含量的強烈影響,植株通過改變根系形態更有效地從土壤中獲取氮素營養。在低氮條件下,玉米的節根數目減少、總根長增加,而在氮充足的土壤側根易發生和伸長[7–8]。因此,可以利用玉米根系形態對土壤氮水平響應的可塑性來提高玉米的氮吸收效率。目前數量性狀位點(QTL)作圖已成為研究復雜性狀遺傳位點甚至確定候選基因的有力工具。許多與玉米氮效率相關性狀的QTL已經被定位,包括氮吸收效率、氮利用效率[9]、花后氮吸收、氮再利用[1],以及氮同化、代謝等性狀[9]。同樣,諸多控制玉米根系形態的QTL也在不同的群體中被定位和分析[10–14]。但是,這些研究多數都是對正常生長條件下玉米根系形態相關性狀進行QTL定位,對低氮脅迫下玉米根系形態相關性狀的QTL定位相對缺乏。
劉宗華等[15]前期研究結果表明,在大田不施氮肥條件下,綜3的產量降低了54%,而許178的產量僅降低了1.2%,表明許178對低氮脅迫不敏感,而綜3對低氮脅迫表現敏感。本研究以綜3為供體親本,許178為輪回親本構建的染色體單片段代換系(single segment substitution lines,SSSLs)[16]作為研究材料,通過營養液培養對群體在高氮和低氮條件下根系形態相關性狀,以及氮吸收量的QTL進行了定位。研究分析了低氮脅迫下玉米苗期根形態與氮吸收效率之間的遺傳機制,為通過分子標記輔助選擇培育低氮下具有優異根系形態的玉米基因型提供參考。
1 材料與分析
1.1 供試材料
以綜3為供體親本、許178為受體親本,經過4個世代回交和3個世代自交,結合SSR分子標記輔助選擇,共獲得了239份純合的單片段代換系材料,SSSL平均代換片段長度為47.67cM,導入片段總長11394.22cM,覆蓋玉米基因組67.6%[15]。利用其中的150個單片段代換系進行玉米苗期不同氮水平下根系形態QTL檢測,同時結合各株系表型值及片段所在位置,對相應根系形態QTL進行初步定位。
1.2 幼苗培養
取150個單片段代換系及親本各30粒飽滿完整的種子,以10%H2O2浸泡消毒30min,蒸餾水沖洗3~4次,置于濕潤的紗布上催芽,然后移入干凈的石英砂中育苗,光照培養室日間溫度為25℃,夜間溫度為20℃。用蒸餾水培養至兩葉,挑取發育健壯、長勢一致的幼苗共12株,分別移入低氮和高氮營養液中培養,每個氮水平6株。幼苗定植于穿孔直徑為2cm、厚0.5cm的塑料板上,用海綿固定幼苗,培養容器采用黑色方形塑料盒(38cm×28cm× 12cm),每盆培養幼苗12株,內盛營養液10L,3天更換一次營養液。營養液配方參考文獻[17],具體如下:Ca(NO3)2·4H2O2mmol/L;K2SO40.75mmol/L;MgSO4·7H2O0.65mmol/L;KCl:0.1mmol/L;KH2PO40.25mmol/L;H3BO30.001mmol/L;MnSO4·H2O 0.001mmol/L;CuSO4·5H2O0.0001mmol/L;ZnSO4·7H2O0.001mmol/L;(NH4)6MoO24·4H2O5× 10–6mmol/L;FeSO4·7H2O0.1mmol/L;Na2EDTA 0.1mmol/L;CaCl22mmol/L(低氮處理時加入,用以補平Ca離子濃度)。其中低氮處理氮濃度為0.05 mmol/L,高氮水平含氮4mmol/L,共培養20天。
1.3 根系性狀的測定
培養20天后,每個重復取生長一致的4株幼苗,蒸餾水沖洗干凈,用剪刀剪下根系,WinRHIZO根系分析系統分別測定單株玉米的總根長(TRL,cm)、根尖數(RTN)、根體積(RV,cm3)、根表面積(RSA, cm2)、根直徑(RAD,mm)。將新鮮根和莖葉分別置于105℃烘箱殺青30min,然后80℃烘干,測定地上部干重(SDW)、根干重(RDW)。
1.4 氮含量的測定
植株烘干粉碎后,采用H2SO4–H2O2進行消化,通過流動注射儀測定氮含量。植株氮累積量計算:
地上部氮累積量(SNU,mg)=地上部干重(g)×地上部氮含量(mg/g);
根氮累積量(RNU,mg)=根部干重(g)×根部氮含量(mg/g)。
1.5 QTL 定位
利用SPSS19.0軟件對各染色體片段代換系和雙親本根系相關數據進行統計分析。將輪回親本的觀察值作為對照,通過方差分析和T檢驗比較各SSSL與許178之間的差異,以P≤0.001時,認為該代換片段上有相關QTL存在;當P>0.001時,認為在該代換片段上沒有相關QTL的存在[18]。參照Eshed和Zamir的方法[19]估算各QTL的加性效應值和加性效應貢獻率,加性效應值=(SSSL表型值–許178表型值)/2,加性效應貢獻率=加性效應值/許178表型值×100%。
QTL命名遵循Mc Couch等[20]制定的原則,QTL名稱中前面的字母為鑒定性狀的英文詞首的縮寫,第1個數字為QTL所在染色體的編號,第2個數字為該性狀在該染色體上的QTL序號。
1.6 QTL 代換作圖
參照Paterson等[20]的方法,如果在含有重疊代換片段的不同染色體片段代換系中同時檢測到同一性狀的QTL,且遺傳效應方向一致,則認為該QTL存在于代換片段的重疊區段上;如果在一個染色體片段代換系中檢測到QTL的存在,而在代換片段具有重疊關系的另一個染色體片段代換系中未檢測到,則認為該QTL位于這2個代換系代換片段的非重疊區段上。
1.7 數據處理
采用Microsoft Excel2010和SPSS19.0軟件對數據進行整理和統計分析。

圖1 高氮 (HN) 和低氮 (LN) 水平下許 178 和綜 3的根系形態Fig. 1 Root morphology of Xu178 and Zong3 under high and low nitrogen conditions
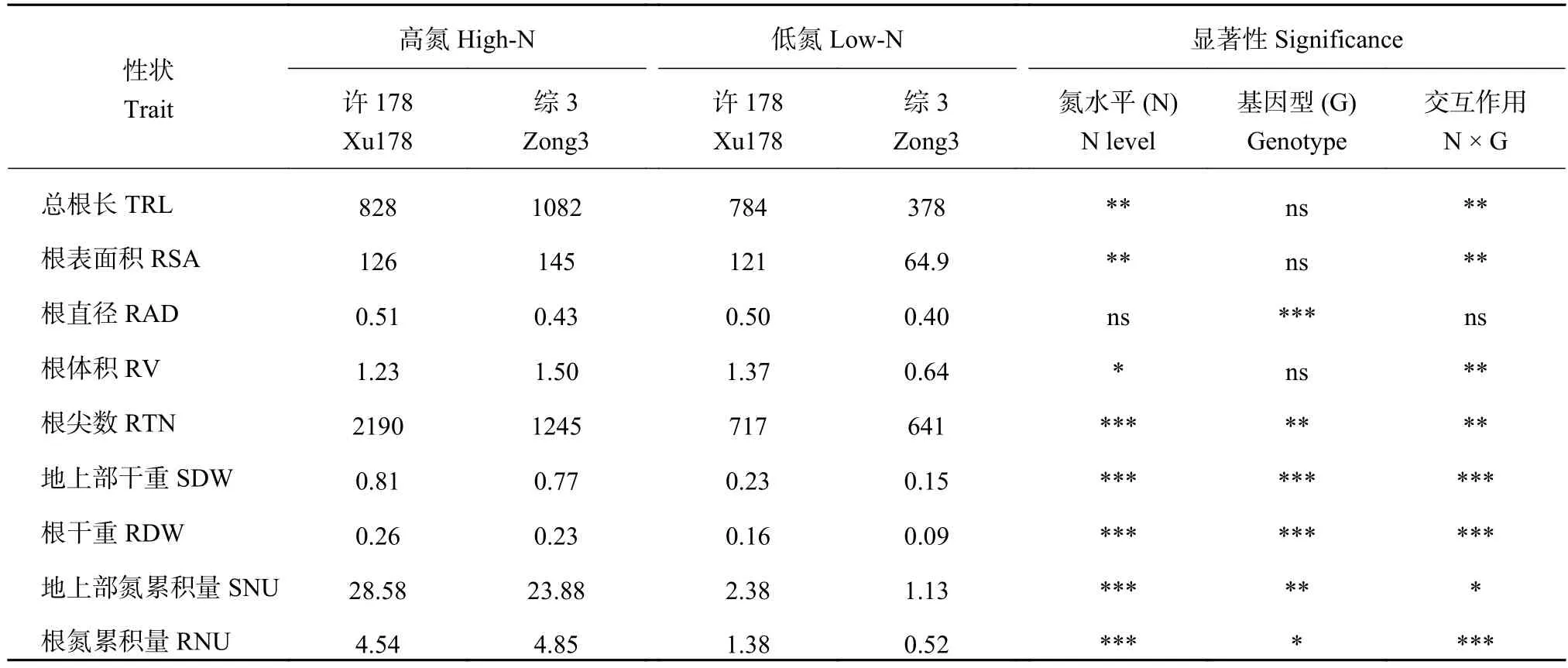
表1 不同氮水平下親本各表型值和雙因素顯著性分析Table 1 Mean values and significance of two-factor ANOVA analysis for the nine traits in the parents
2 結果與分析
2.1 不同氮水平下親本和 SSSL 群體各表型性狀值分析
高氮條件下,親本許178和綜3的根系形態并沒有顯著差異,但是低氮脅迫顯著增加了許178的根長,其根系遠大于綜3(圖1),表明在低氮條件下許178具有明顯的根系優勢。
數據統計結果顯示,高氮下兩親本除了根尖數有顯著差異外,其他根系性狀均無顯著差異(表1)。低氮條件下,氮高效親本許178的生物量顯著高于綜3,并且許178的根系性狀(總根長、根表面積、根體積、根直徑和根尖數)大于綜3,最終導致許178植株總氮吸收量是綜3的2.3倍。對親本9個性狀進行雙因素主效應分析發現,除了根直徑外,其他8個性狀氮水平的主效應差異均達到顯著或極顯著差異,而兩親本的基因型差異在總根長、根表面積和根體積上沒有達到顯著水平,但是除了根直徑外,氮水平和基因型的交互作用在兩親本間的差異均達到顯著或極顯著水平(表1)。
在兩個氮水平下,SSSL群體的9個性狀均表現連續變異,接近正態分布(表2)。這些性狀的變異系數從5.6%~33.9%。根直徑的變異系數較小,其他性狀的變異系數均大于20%。在兩個氮水平下,每個性狀均觀察到超親現象。在SSSL群體中,基因型對9個性狀的主效應作用均達到顯著或極顯著差異。除了根干重外,氮水平及其與基因型的交互作用對其他8個性狀的作用均達到顯著水平。這些結果表明,研究所選用的SSSL群體對氮脅迫敏感,并且群體基因型差異性顯著,有利于我們利用該群體對根性狀和氮吸收相關QTL進行定位和進一步深入分析。
2.2 不同氮水平下 SSSL 群體各表型性狀的相關性分析
高氮條件下SSSL群體除了根直徑與總根長和根尖數沒有顯著相關性以外,其它各性狀之間均呈顯著或極顯著正相關,并且各根系性狀與植株生物量均呈極顯著正相關,植株氮吸收量也與根系各性狀呈顯著或極顯著正相關(表3)。在低氮條件下,根直徑與總根長、根表面積、地上部生物量之間均沒有顯著相關性,其他根系性狀之間均顯著相關。此外,低氮條件下,除了根直徑,植株氮吸收量與其他根系性狀均呈極顯著正相關性。在兩個氮水平下,均是根體積與根表面積的相關性最大(高氮條件r= 0.913,P<0.001;低氮條件r=0.927,P<0.001)。高氮條件下,地上部氮累積量與根表面積相關性最高,根系氮累積量則與根體積的相關性最高,而在低氮條件下,地上部和根系氮累積量均與根表面積的相關性最大。
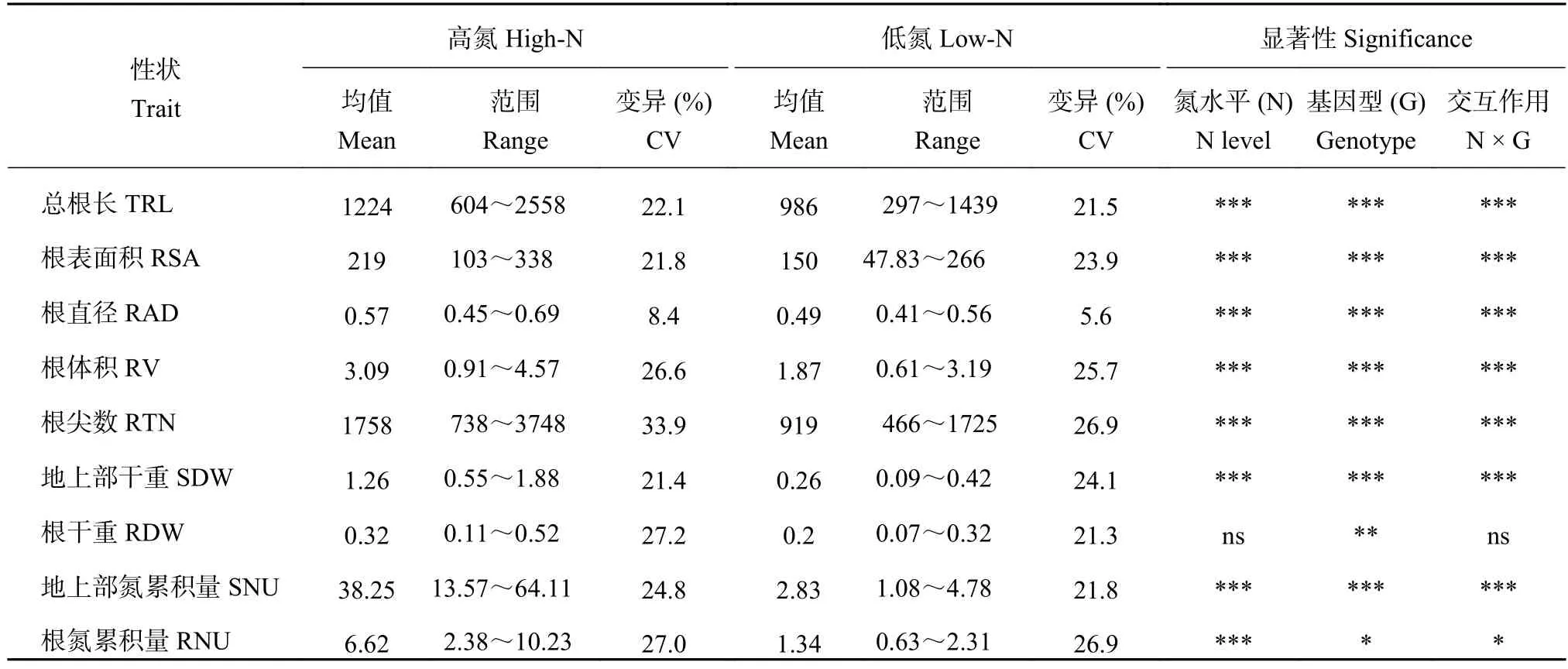
表2 不同氮水平下 SSSL 群體各表型值和雙因素顯著性分析Table 2 Mean values and significance of two-factor ANOVA analysis for the nine traits in the SSSL population
2.3 不同氮條件下玉米根系形態 QTL 分析
通過QTL代換作圖,對SSSL群體檢測到的根系形態QTL進行分析。高氮水平下,在P≤0.001條件下共檢測到40個根系形態QTL,其中8個總根長QTL,7個根表面積QTL,4個根直徑QTL,10個根體積QTL和11個根尖數QTL(表4)。總根長QTL主要分布在第1、3、5、7、9和10號染色體,第1染色體最多,其中qhnTRL1b表現為負向加性效應,其余均表現為正向加性效應。代換系1306包含的qhnTRL1c加性效應貢獻率最大(59.8%)。檢測到根表面積QTL與總根長類似,僅有一個QTL表現負向加性效應,其余均為正向,加性效應貢獻率為–23.5%~83.7%。檢測到的根直徑QTL的數量最少,均為正向加性效應,貢獻率從14.1%~17.4%。檢測到的根體積的QTL均為正向加性效應,且加性效應貢獻率很高,介于79.4%~125.1%,這表明在高氮水平下綜3片段的導入可能有利于代換系獲得更大的根系體積。與其他根系性狀不同,檢測到的10個根尖QTL中有9個表現負向加性效應,貢獻率為–23.2%~–36.5%,只有qhnRTN7a為正向加性效應。
低氮水平下,通過QTL代換作圖,在P≤0.001條件下共檢測到47個根系形態QTL,其中10個總根長QTL,5個根表面積QTL,5個根直徑QTL,12個根體積QTL和15個根尖數QTL(表5)。其中檢測到的總根長和根表面積QTL與高氮條件類似,
多為正向加性效應,各有一個QTL為負向加性效應。與高氮條件不同,低氮下檢測到的根直徑QTL均為負向加性效應,加性效應貢獻率為–5.0%~–8.9%。低氮下檢測到的根尖數QTL多為正向加性效應,僅qlnRTN4b表現為負向加性效應,并且加性效應的貢獻率低于高氮條件,這可能與低氮脅迫下植物根系的適應性反應有關。檢測到的12個根體積QTL中有3個為負向加性效應,其中在第1染色體檢測到5個QTL。

表3 群體各表型性狀在高氮水平 (對角線上方) 和低氮水平 (對角線下方) 的 Pearson 相關系數Table 3 Pearson’s correlation coefficients between the traits under high N (above diagonal) and low N conditions (below diagonal) in the SSSL population
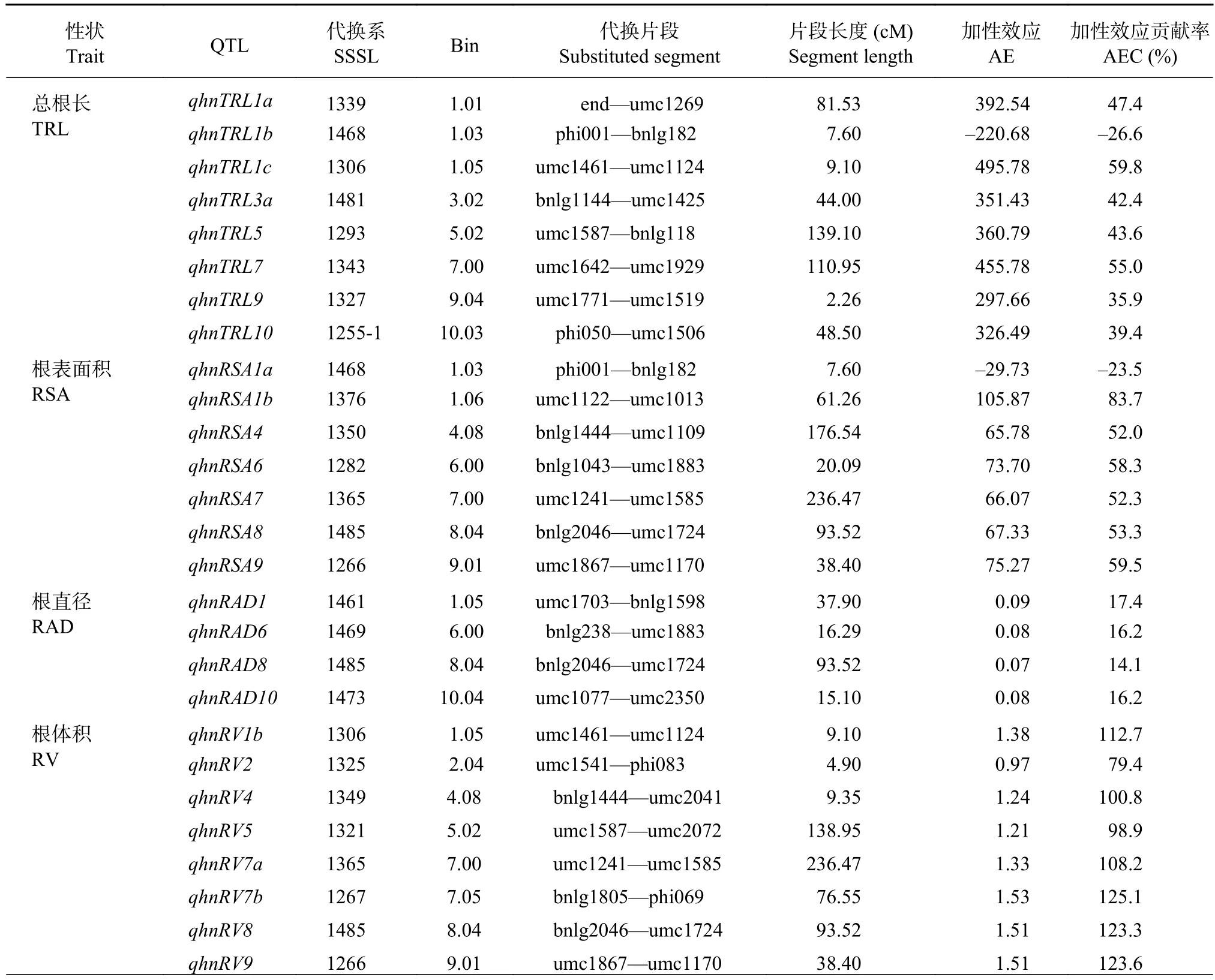
表4 高氮下玉米 SSSL 群體的根系形態 QTL 及其效應值Table 4 QTL location of the maize root morphology in the SSSL population under the high nitrogen condition

續表4Table4continued
2.4 不同氮條件下生物量 QTL 分析
高氮條件下,通過QTL代換作圖在P≤0.001時,SSSL群體共檢測到20個地上部干重QTL和14個根干重QTL(表6)。20個地上部干重QTL分布在除第8染色體的其余9條染色體上。其中qhnSDW1b表現為負向加性效應,其余QTL均表現為正向加性效應,單個QTL的加性效應貢獻率為–40.1%~88.3%。14個根干重QTL分布在除第2、8染色體外的其他8條染色體上,其中有4個表現負向加性效應,其他10個QTL為正向加性效應。單個QTL的加性效應貢獻率為–10.2%~42.8%。
低氮條件下,通過QTL代換作圖,在P≤0.001條件下共檢測到22個生物量QTL,地上部干重QTL和根干重QTL均為11個(表7)。其中地上部干重QTL有6個分布在第1染色體,其余分布在第3、4、5、7和10號染色體上,加性效應貢獻率為–24.5%~59.9%,qlnSDW5的加性效應貢獻率最高。與地上部干重QTL分布情況類似,檢測到的根干重QTL也有5個分布于第1染色體,其余分布在第2、3、4、5和7號染色體上,單個QTL的加性效應貢獻率為–19.2%~48.4%。
2.5 不同氮條件下植株氮累積量 QTL 分析
高氮條件下,通過QTL代換作圖,在P≤0.001條件下共檢測到13個地上部氮累積量QTL和15個根部氮累積量QTL(表8)。這些QTL中只有qhnSNU1b和qhnRNU1a表現為負向加性效應。除第3和第8染色體外,其余染色體均檢測到地上部氮累積量QTL,單個QTL的加性效應貢獻率為–42.7%~60.9%。根部氮累積量的QTL則在10條染色體上均有分布,其中第1和9染色體分別檢測到3個QTL,這些QTL的加性效應貢獻率為–35.5%~71.7%。
低氮條件下,共檢測到6個地上部氮累積量QTL和10個根部氮累積量QTL(P≤0.001)(表9)。6個地上部氮累積量QTL均為正向加性效應,并且3個位于第1染色體上,單個QTL的貢獻率為33.2%~55.2%。檢測到的10個根部氮累積量QTL中有6個表現負向加性效應,4個為正向加性效應,單個QTL的貢獻率為–27.3%~33.8%。
2.6 QTL 在染色體上的分布
兩個氮水平下檢測到的QTL在染色體上的分布如圖2所示。結果顯示,所檢測到的根系性狀和氮吸收相關QTL在玉米的10條染色體上均有分布,其中第1染色體上的QTL數目最多,第2和第8染
色體上的QTL數目最少。對低氮和高氮下分別檢測到的QTL進行分析可以得到不同氮條件下的特異性QTL。有些染色體區間僅能在低氮水平下檢測到QTL,例如第1染色體bnlg182—bnlg2295(代換系1419的代換區間)和umc1013—umc2047(代換系1314的代換區間)區間。包含這些染色體片段的代換系很有可能只控制低氮條件下的根系形態,有利于我們發掘低氮特異的QTL。同樣,圖2結果也顯示在有些染色體區段僅檢測到高氮條件下QTL,例如,第1染色體phi001—bnlg182(bin1.03,代換系1428的代換區間),umc1122—umc2396(bin1.06,代換系1376的代換區間),第6染色體bnlg1043—umc1883(代換系1282的代換區間),第9染色體的umc2078—umc1636(代換系1238的代換區間),以及第10染色體的umc1077—umc2350(代換系1473的代換區間)。這些代換系在低氮條件下未檢測到QTL,我們將其定義為高氮特異的QTL。這兩類QTL為氮環境特異性QTL,說明在不同氮條件下玉
米有不同的代謝機制。其余的QTL區間均同時檢測到高氮和低氮條件下的QTL。此外,從QTL在染色體上的分布可以看出,玉米氮吸收相關的QTL區間均能檢測到根系形態相關的QTL,說明玉米根系形態與其氮吸收能力密切相關(圖2)。
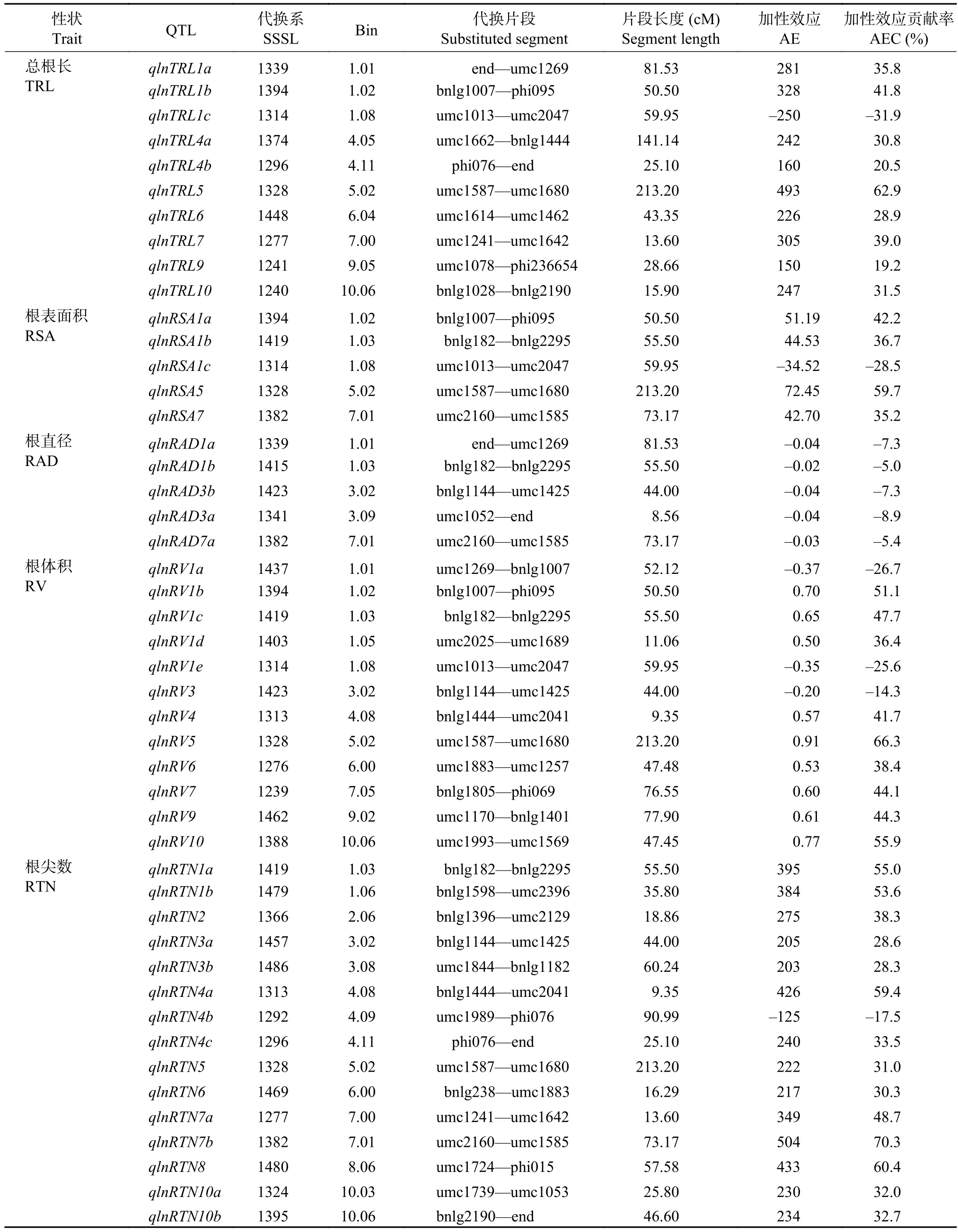
表5 低氮下玉米 SSSL 群體的根系形態 QTL 及其效應值Table 5 QTL location of the root morphology in the SSSL population under the low nitrogen condition

表6 高氮條件下玉米苗期生物量 QTL 及其效應Table 6 QTL location of plant biomass in the SSSL population under the high nitrogen condition
3 討論
玉米已成為我國種植面積最大的農作物,但是目前其高產和超高產依賴大量的氮肥施用。如何提高玉米的氮肥利用率、減少肥料投入、保護環境已成為玉米高產高效的研究熱點和亟待解決的問題。植物對土壤中氮素的獲取很大程度上取決于其根系形態[22]。研究者通過對不同基因型玉米進行研究提出了玉米氮高效吸收的理想根系形態,即通過增加根體積,包括根干重、根長和根密度,提高玉米氮吸收能力和籽粒產量[8,23]。研究表明玉米的根系形態受到土壤中有效氮含量影響,在缺氮條件下玉米節根數目降低,但是總根長增加[7]。玉米通過這種根系形態的改變從土壤中獲取更多的氮素,用于植株的生長發育。本研究結果也顯示,在低氮條件下,代換系玉米幼苗莖葉的生物量與總根長、根表面積、根體積、根尖數呈極顯著相關性,而與根直徑之間的相關性未達到顯著水平。并且,低氮下植株氮吸收量與總根長、根表面積、根體積和根尖數均呈極顯著正相關(表3)。這些結果說明,通過對玉米根性狀相關QTL的研究有利于深入認識玉米氮吸收的遺傳機制。
氮吸收效率和根系形態都是復雜的性狀,受基因型和環境的共同影響。目前我們對玉米氮高效和根系形態的遺傳基礎仍知之甚少,對于兩者之間的關系認識不足,這也限制了基于根系形態來篩選氮高效材料或品種的效率。近來QTL作圖已經成為確定復雜性狀的遺傳位點甚至候選基因的有效工具。很多氮高效相關的農藝性狀已經被定位,包括生物量[25]、籽粒氮吸收量[26]、吐絲后氮吸收效率以及氮再利用效率等[5]。同時,很多調控根系形態的QTL也在不同的玉米連鎖群中被定位[10–11,27–28]。例如,Burton等[27]利用三個玉米重組自交系對21個根系性狀進行QTL定位,共定位到15個QTL,分布在8條染色體上(第6和10染色體上沒有檢測到QTL),表型變異為0.44%~13.5%。Burton等[27]對Zhu等[13]、Lebreton等[12]研究中所定位到的QTL進行元分析,發現在bin 1.03、1.04和7.03位置分別有控制初生胚根上側根長度、節根數目和次生胚根數目的QTL。此外,Song等[11]利用營養液培養方法,對含有204個材料的重組自交系根系性狀進行QTL定位,在bin1.06檢測到平均側根長度(SLL)和根冠比的QTL。本研究利用氮效率差異顯著的兩個玉米自交系構建的單片段代換系群體對玉米苗期的根系形態和氮吸收量進行了QTL定位分析。在高氮條件下,共檢測到40個根系性狀QTL,34個生物量QTL,以及28個氮吸收量QTL。對HN下的QTL進行分析發現,有些染色體區段僅特異檢測到HN下的QTL,并且不同性狀的QLT成簇排列(圖2),這些區段檢測到HN下QTL不少于5個,表明這些區段可能包含有高氮條件下控制玉米根系形態的主效QTL。本研究也在bin1.03(第1染色體phi001—bnlg182區間)和bin1.06(umc1122—umc2396區間)檢測到正常氮條件下根形態QTL,與Burton等[27]和Song等[11]定位到的根形態相關QTL區間有重疊。這些結果表明在這兩個QTL區間可能存在控制氮充足條件下玉米根系形態的主效QTL。目前就我國農業生產現狀而言,氮肥的施用量短期內很難顯著降低[29],因此,培育在正常或高肥力條件下高效吸收的品種也是提高氮效率的途徑之一[30]。在高產栽培條件下,隨著產量水平的提高,作物需氮量增加,施氮總量也會提高。但是,我國目前玉米農田生產中多數農民采用一次性基肥,不僅增加了硝酸鹽淋失的可能性,而且過高的硝酸鹽濃度也限制了根系的生長發育,進而降低氮肥的吸收效率。因此,米國華等[31]提出玉米氮高效吸收的理想根構型具有在高產氮肥投入條件下仍能維持正常的側根生長、總根長密度高,提高整體剖面氮素有效性的特點。本研究中所定位到的高氮特異的QTL簇對研究玉米高氮條件下氮高效吸收的遺傳機制和基因定位具有一定意義。
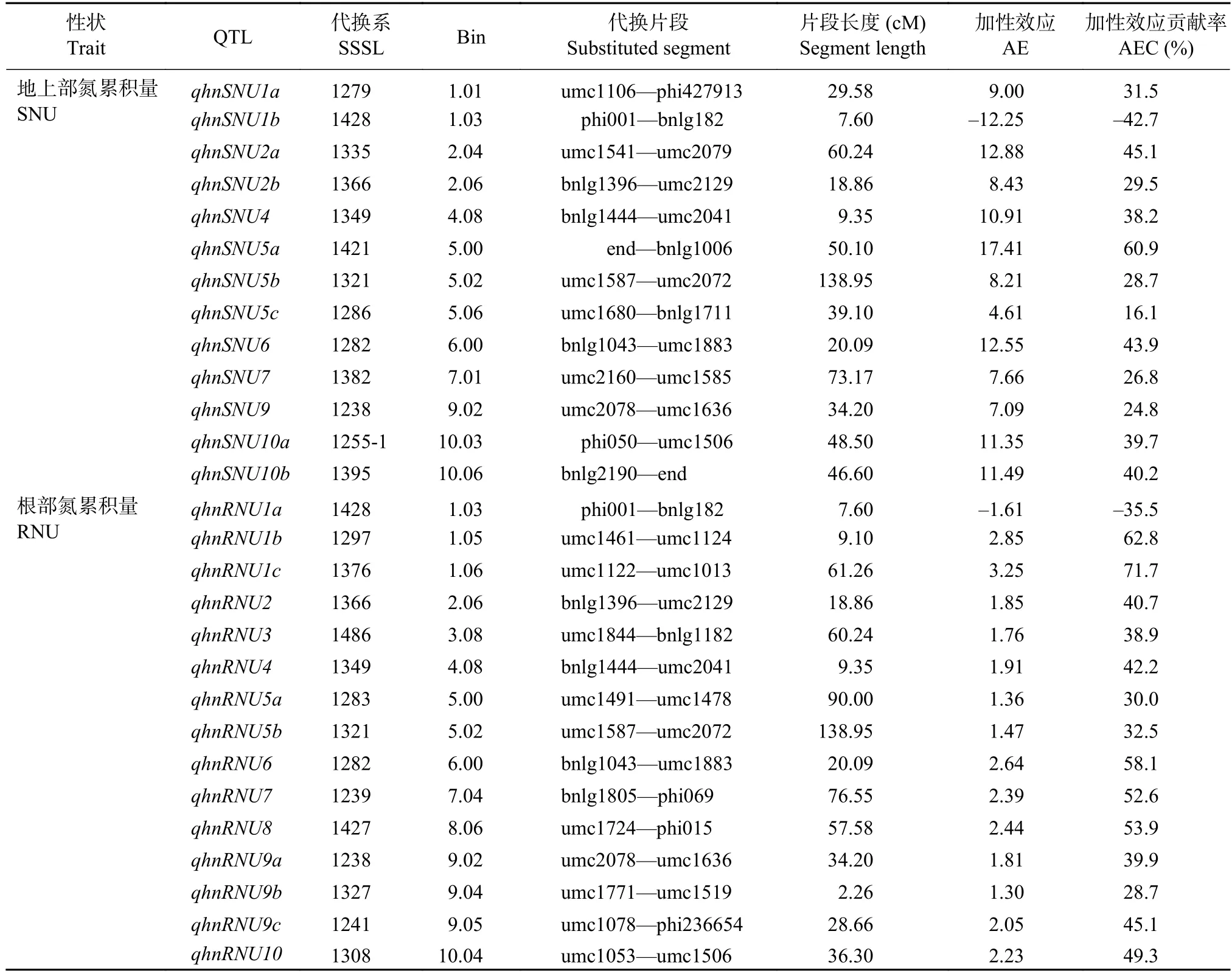
表8 高氮條件下苗期玉米植株氮累積量 QTL 分析Table 8 QTL location of the nitrogen accumulation in the SSSL population under the high nitrogen condition
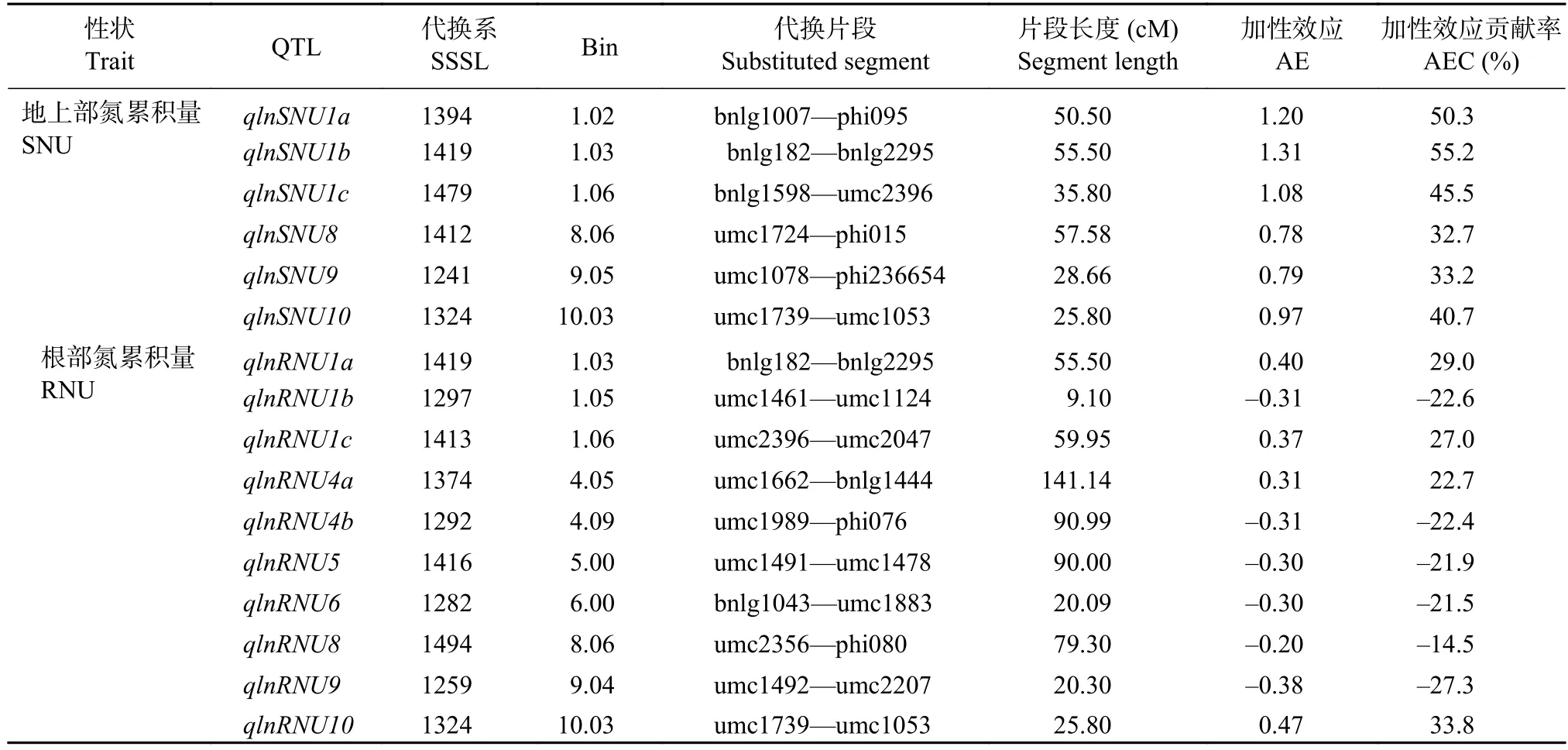
表9 低氮條件下苗期玉米植株氮累積量 QTL 分析Table 9 QTL location of the nitrogen contents in the SSSL population under the low nitrogen condition
本研究在低氮條件下共檢測到85個根性狀相關QTL,在第1染色體bnlg182—bnlg2295(bin1.04)和umc1013—umc2047(bin1.08)區間不僅檢測到低氮下根系形態相關QTL,而且同時檢測到低氮下生物量和氮吸收量的QTL,說明低氮脅迫下植株氮吸收量與根系形態緊密聯系。其中,第1染色體bnlg182—bnlg2295(bin1.04)區間在Burton等[27]、Song等[11]的研究中均檢測到根系形態相關QTL。Li等[14]利用重組自交系,設置高氮和低氮水平,通過大田試驗和水培實驗對氮利用效率(NUE)和根系構型(RSA)相關性狀的QTL進行定位,共檢測到331個QTL。該研究發現氮吸收效率與根系構型之間存在顯著的表型相關性。研究表明大約70%的NUE相關QTL簇與根系構型QTL簇重疊,這表明根系構型與氮利用率相關性狀之間有顯著的遺傳學關系,可以通過分子標記篩選優異的根系構型來提高玉米的氮利用效率[14]。本研究結果也顯示,除了極個別的代換區間僅單獨檢測到氮累積量的QTL,其他絕大部分植株氮吸收量QTL都與根形態相關QTL成簇存在(圖2)。此外,Li等[14]的研究發現有5個重要的QTL簇對玉米獲得大的根系和更高的氮吸收效率有利,其中一個QTL簇位于染色體區間bin1.04。這一結果與我們發現的QTL簇區間bnlg182—bnlg2295(bin1.04)有重疊,并且在這個QTL區間同時檢測到低氮下植株氮吸收量的QTL,表明該區間很有可能包含有控制玉米氮高效的主效QTL位點,進一步對包含該區段的代換系1419進行精細定位可能獲得氮高效候選基因。Gallais等[9]通過大田和水培相結合的方法對玉米花后氮素吸收、氮再利用、籽粒產量、葉片硝酸還原酶、谷氨酰胺合成酶活性等性狀進行了QTL定位,發現三個染色體位置上檢測到產量QTL、氮再利用QTL、GS活性QTL和一個編碼胞質GS的基因具有一致性,并且位于第5染色體的GS位點能夠部分解釋玉米氮利用效率的表型變異。同時,該研究定位的QTL簇有些與本研究重要QTL簇位置接近,例如,Gallais等[9]在第1染色體的頂端區間檢測到莖稈氮再利用QTL、GDH和GS酶活性相關QTL,而我們的結果顯示在同一區段檢測到總根長、植株生物量等QTL;在位于第5染色體umc1587—umc1680(代換系1328的代換區間)的QTL簇接近的位置,Gallais等[9]定位到葉片硝酸鹽含量、GS和NR活性QTL;在bin1.08位置(umc1013—umc2047區間,代換系1314)Gallais等檢測到控制整株氮再利用的QTL。此外,齊歡歡等[32]研究發現第1染色體的umc2217—umc1770區段與穗長、穗位高和穗行數有關,這一區間與我們的QTL簇bnlg182—bnlg2295(代換系1419)部分重疊。這說明本研究所定位到的低氮特異QTL簇可能在玉米低氮條件下氮素高效吸收中起重要作用,與氮高效吸收密切相關。此外,在一些QTL區間同時檢測到多個高氮和低氮QTL,且成簇排列,這些QTL不受氮水平特異誘導。例如第3染色體bnlg144—umc1425區間,第7染色體umc1241—umc1642區間,第8染色體umc1724—phi080區間(圖2)。這些不同環境下檢測到的QTL簇表明玉米在不同的氮環境下具有不同的代謝機制。高、低氮特異的QTL簇對我們研究玉米適應氮脅迫具有重要意義,有利于挖掘不同氮水平下的氮高效基因,更有針對性地開展玉米氮高效育種。

圖2 不同氮水平下玉米苗期根系形態及氮累積量 QTL 在染色體上的分布Fig. 2 Position of the QTLs for maize root morphology and N uptake at the seedling stage under low and high N levels
4 結論
不同氮水平下,玉米的根系形態存在顯著差異,對高、低氮條件下檢測到的特異QTL進行分析有助于解析不同氮水平下玉米高效吸收氮素的遺傳機制。本研究在染色體區段bnlg182—bnlg2295 (bin1.04,代換系1419)、umc1013—umc2047 (bin1.08,代換系1314)區間檢測到多個低氮特異的QTL,在phi001—bnlg182(bin1.03,代換系1428)和umc1122—umc2396(bin1.06,代換系1376)等區間檢測到高氮條件下特異的QTL。這些QTL區間同時包含根系形態QTL和氮吸收量QTL,并且在前人的研究中均有玉米氮效率或根系形態相關QTL檢測到,說明這些區間在玉米氮高效吸收中可能起重要作用,后期會對包含該區段的代換系進行精細定位,獲得候選基因。
[1]Mi G,Chen F,Zhang F.Physiological and genetic mechanisms for nitrogen use efficiency in maize[J].Journal of Crop Science Biotechnology,2007,10(2):57–63.
[2]Xu G,Fan X,Miller AJ.Plant nitrogen assimilation and use efficiency[J].Annual Review of Plant Biology,2012,63:153–182.
[3]DeBruin J,Messina CD,Munaro E,et al.N distribution in maize plant as amarker for grain yield and limits on its remobilization after flowering[J].Plant Breeding,2013,132(5):500–505.
[4]Mackay AD,Barber SA.Effect of nitrogen on root growth of two corn genotypes in the field[J].Agronomy Journal,1986,78(4): 699–708.
[5]Coque M,Martin A,Veyrieras JB,et al.Genetic variation for N-remobilization and post-silking N-uptake in aset of maize recombinant inbred lines.3.QTL detection and coincidences[J]. Theoretical and Applied Genetics,2008,117(5):729–747.
[6]Mu X,Chen F,Wu Q,et al.Genetic improvement of root growth increases maize yield via enhanced post-silking nitrogen uptake[J]. European Journal of Agronomy,2015,63:55–61.
[7]Saengwilai P,Tian X,Lynch JP.Low crown root number enhances nitrogen acquisition from low-nitrogen soils in maize[J].Plant Physiology,2014,166(2):581–589.
[8]Lynch JP.Steep,cheap and deep:an ideotype to optimize water and N acquisition by maize root systems[J].Annals of Botany,2013, 112(2):347–357.
[9]Gallais A,Hirel B.An approach to the genetics of nitrogen use efficiency in maize[J].Journal of Experimental Botany,2004, 55(396):295–306.
[10]Hochholdinger F,Tuberosa R.Genetic and genomic dissection of maize root development and architecture[J].Current Opinion of Plant Biology,2009,12(2):172–177.
[11]Song W,Wang B,Hauck AL,et al.Genetic dissection of maize seedling root system architecture traits using an ultra-high density bin-map and arecombinant inbred line population[J].Journal of Integrate Plant Biology,2016,58(3):266–279.
[12]Lebreton C,Lazi?-Jan?i?V,Steed A,et al.Identification of QTL for drought responses in maize and their use in testing causal relationships between traits[J].Journal of Experimental Botany,1995, 46:853–865.
[13]Zhu J,Kaeppler SM,Lynch JP.Mapping of QTLs for lateral root branching and length in maize(Zea mays L.)under differential phosphorus supply[J].Theoretical and Applied Genetics,2005, 111(4):688–695.
[14]Li P,Chen F,Cai H,et al.A genetic relationship between nitrogen use efficiency and seedling root traits in maize as revealed by QTL analysis[J].Journal of Experimental Botany,2015,66(11): 3175–3188.
[15]毛克舉,李衛華,付志遠,等.玉米自交系許178背景的綜3染色體單片段代換系的構建[J].河南農業大學學報,2013,47(1):6–9,15. Mao KJ,Li WH,Fu Zh Y,et al.Development of aset of signal segment substitution lines of an elite inbred line Zong3on the genetic background Xu178in maize(Zea mays L.)[J].Journal of Henan Agriculture University,2013,47(1):6–9,15.
[16]劉宗華,王春麗,湯繼華,等.氮脅迫對不同玉米自交系若干農藝性狀和產量的影響[J].河南農業大學學報,2006,40(6):573–577. Liu ZH,Wang CL,Tang JH,et al.Effect of low nitrogen stress on several agronomy characters and grain yield of different maize inbreds[J].Journal of Henan Agriculture University,2006,40(6): 573–577.
[17]田秋英.根系形態在玉米高效獲取氮素中的作用及其生理調節機制[D].北京:中國農業大學博士學位論文,2005. Tian QY.Role of root morphology in efficient nitrogen acquisition in maize and its physiological regulation mechanism[D].Beijing: PhD Dissertation of China Agricultural University,2005.
[18]劉冠明,李文濤,曾瑞珍,等.水稻單片段代換系代換片段的QTL鑒定[J].遺傳學報,2004,12:1395–1400. Liu GM,Li WT,Zeng RZ,et al.Identification of QTLs on substituted segments in signal segment substitution lines of rice[J]. Acta Genetica Sinica,2004,12:1395–1400.
[19]Eshed Y,Zamir D.An introgression line population of Lycopersicon Pennellii in the cultivated tomato enables the identification and fine mapping of yield-associated QTL[J].Genetics,1995,141(3): 1147–1162.
[20]McCouch SR,Cho YG,Yano M.Report on QTL nomenclature[J]. Rice Genetics Newsletter,1997,14:11–13.
[21]Paterson AH,DeVerna JW,Lanini B,et al.Fine mapping of quantitative trait loci using selected overlapping recombinant chromosomes,in an interspecies cross of tomato[J].Genetics,1990, 124(3):735–742.
[22]Forde BG.Nitrogen signaling pathways shaping root system architecture:an update[J].Current Opinion of Plant Biology,2014, 21:30–36.
[23]Mi G,Chen F,Wu Q,et al.Ideotype root architecture for efficientnitrogen acquisition by maize in intensive cropping systems[J]. Science China(Life Sciences),2010,53(12):1369–1373.
[24]李卓坤,彭濤,張衛東,等.利用“永久F_2”群體進行小麥幼苗根系性狀QTL分析[J].作物學報,2010,36(3):442–448. Li ZK,Peng T,Zhang WD,et al.Analysis of QTLs for root traits at seedling stage using an"Immortalized F2"population of wheat[J]. Acta Agronomic Sinica,2010,36(3):442–448.
[25]Cai H,Chu Q,Yuan L,et al.Identification of quantitative trait loci for leaf area and chlorophyll content in maize(Zea mays)under low nitrogen and low phosphorus supply[J].Molecular Breeding,2012, 30(7):251–266.
[26]Liu Q,Qin J,Li T,et al.Fine mapping and candidate gene analysis of qSTL3,a stigma length-conditioning locus in rice(Oryza sativa L.)[J].PloS One,2015,10:e0127938.
[27]Burton AL,Johnson JM,Foerster JM,et al.QTL mapping and phenotypic variation for root architectural traits in maize(Zea mays L.)[J].Theoretical and Applied Genetics,2014,127(11):2293–2311.
[28]Salvi S,Giuliani S,Ricciolini C,et al.Two major quantitative trait loci controlling the number of seminal roots in maize co-map with the root developmental genes rtcs and rum1[J].Journal of Experimental Botany,2016,67(4):1149–1159.
[29]武良,張衛峰,陳新平,等.中國農田氮肥投入和生產效率[J].中國土壤與肥料,2016,(4):76–83. Wu L,Zhang WF,Chen XP,et al.Nitrogen fertilizer input and nitrogen use efficiency in Chinese farmland[J].Soils and Fertilizers Sciences in China,2016(4):76–83.
[30]米國華,陳范駿,春亮,等.玉米氮高效品種的生物學特征[J].植物營養與肥料學報,2007,13(1):155–159. Mi GH,Chen FJ,Chun L,et al.Biological characteristics of nitrogen efficient maize genotypes[J].Plant Nutrition and Fertilizer Science,2007,13(1):155–159.
[31]米國華,陳范駿,吳秋平,等.玉米高效吸收氮素的理想根構型[J].中國科學(生命科學),2010,40(12):1112–1116. Mi GH,Chen FJ,Wu QP,et al.Ideotype root architecture for efficient nitrogen acquisition by maize in intensive cropping systems[J].Science China(Life Sciences),2010,40(12):1112–1116.
[32]齊歡歡,段利超,胡偉,等.利用導入系群體對玉米產量及產量相關性狀進行定位分析[J].玉米科學,2013,21(4):24–27,32. Qi HH,Duan LC,Hu W,et al.Identification of signification loci for yield and yield-related traits in maize with introgression lines[J]. Journal of Maize Science,2013,21(4):24–27,32.
Mapping of QTLs for root morphology and nitrogen uptake of maize under different nitrogen conditions
WANG Yi1,2,3,TANG Ji-hua3,FU Yan-lei1,XU Heng3,TAN Jin-fang1,2,HAN Yan-lai1,2,3*
(1 College of Resources and Environment, Henan Agricultural University, Zhengzhou 450002, China; 2 Collaborative Innovation Center of Henan Grain Crops, Zhengzhou 450002, China; 3 State Key Laboratory of Wheat and Maize Crop Science, Zhengzhou 450002, China)
【Objectives】There is astrong relationship between maize root morphology and nitrogen uptake capacity.In this study,QTLs for maize root morphology and plant nitrogen uptake were identified using single segment substitution lines(SSSLs)to provide support for fine mapping and cloning of major QTLs controlling maize root morphology and nitrogen uptake.【Methods】150maize SSSLs derived from across between aN-efficient inbred line Xu178and aN-inefficient inbred line Zong3were used for solution culture.Ca(NO3)2was used as nitrogen source and high nitrogen level(4mmol/L NO3–)and low nitrogen level(0.05mmol/L NO3–)were supplied for each line,and each Nlevel had six seedlings.After20days culture,seedlings were harvested,and the biomass and nitrogen contents of shoots and roots were analyzed respectively.Total root length(TRL),root surface area(RSA),root volume(RV),root average diameter(RAD)and root tip numbers(RTN)were determined from the root images using WinRHIZO.According to the results of the T-test of the phenotype values of SSSL and Xu178,QTLs for each trait were mapped in the SSR genetic linkage map when Pvalue was less than 0.001.【Results】Under the high Nlevel,all root traits were significantly correlated with each other except that between RAD and either TRL or RTN,and the plant nitrogen uptake was significantly correlated with all the root morphology related traits.Under the low Nlevel,all the root morphology traits were strongly correlated with plant nitrogen uptake except for RAD,and the RSA showed the most significant correlation. Under the high Nlevel,102QTLs were detected including40QTLs for root morphology traits,34QTLs for plant biomass and28QTLs for plant nitrogen uptake.Under the low Nlevel,85QTLs were detected including 47QTLs for root morphology traits,22QTLs for plant biomass and16QTLs for plant nitrogen uptake.Most of the QTLs for Nuptake coincided in cluster with those for root morphology.Several QTLs for root morphology and nitrogen uptake were mapped in the same substituted segment region.Under the high Ncondition,five high N-specific QTLs clusters containing several root morphology traits and nitrogen uptake were detected in SSSL lines of1428,1376,1282,1266and1473.The single QTL additive effect contribution was from–43%to84%. Moreover,several QTLs for root morphology and nitrogen uptake were identified in the SSSL lines of1419and 1314under the LN condition,with the additive effect contribution from–32%to55%.【Conclusions】In the present study,several LN-specific QTLs were mapped in substituted segment of bnlg182–bnlg2295in line 1419and umc1013–umc2047in line1314,in which some QTLs related to nitrogen use efficiency of maize were detected in previous researches.It was indicated that there were some major QTLs for maize root morphology and plant nitrogen uptake located in the two regions which would play important role in maize nitrogen uptake efficiency.The present research serves as abasis for the major QTLs fine-mapping and candidate genes cloning.
maize;SSSLs;root morphology;QTL;nitrogen deficiency
2016–12–13接受日期:2017–02–11
國家自然科學基金項目(31601812);河南農業大學科技創新基金項目(KJCX2015A16)資助。
王祎(1983—),女,湖北宜城人,博士,講師,主要從事植物營養生理和遺傳研究。wangyi19830705@163.com
*通信作者E-mail:hyanlai@126.com
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48
