紫花苜蓿藍光受體CRY2B過表達載體構建及轉化
2017-09-13 05:05:26樊文娜李潤林王占彬杜紅旗趙凌平王成章
草地學報 2017年5期
樊文娜, 李潤林, 王占彬, 杜紅旗, 趙凌平, 王成章*
(1. 河南科技大學動物科技學院, 河南 洛陽 471003;2. 河南農業大學牧醫工程學院, 河南 鄭州 450002)
苜蓿(Medicagosativa)的秋眠性(Fall Dormancy)是苜蓿在秋季因日照長度變短和氣溫下降所表現出的一種適應性生長特性[1-2],是選擇栽培苜蓿品種時考慮的首要指標。從本質上講,秋眠性是對日照長度的反應,短日照是其主要的影響因子[3]。苜蓿作為長日照植物,晚春及夏季長日照條件下有利于其生長發育,秋冬季短日照則休眠,說明苜蓿秋眠存在光周期效應[4]。
目前,研究已發現的前3種光信號接受的光受體[5-8],即紅光和遠紅光的受體——光敏色素(Phytochrome )、藍光受體和紫外光受體(UV-receptor)。隱花色素(Cryptochrome)在植物中是很重要的感受光受體,是光形態建成反應的必需元素,但在苜蓿的秋眠中發揮的作用機理目前還未知。高等植物受藍光調節的反應主要包括:向光性、抑制莖芽的生長、葉綠體的轉移、刺激氣孔張開和關閉,從而調節相關基因的表達。藍光光受體——隱花色素可以調控植物開花時間以及晝夜節律循環[9],CRY2B(Cryptochrome2B)是主要接受和傳遞光信息的藍光受體,并且紫花苜蓿轉錄組高通量測序結果差異基因的統計也證明,調控苜蓿秋眠的藍光受體CRY2B的Log2(Fold Change)[10-11]是最大的,這證明CRY2B作為重要的光受體在調控苜蓿的秋眠中發揮著至關重要的作用。
本研究擬構建紫花苜蓿藍光受體CRY2B過表達載體,過表達載體的構建本質上是基因重組的過程。構建紫花苜蓿藍光受體CRY2B過表達載體是獲得紫花苜蓿CRY2B轉基因植株的關鍵步驟和基礎。
1 材料與方法
1.1 試驗材料
1.1.1植物材料 供試的紫花苜蓿(Medicago
sativaL.)品種為一級品種‘馴鹿’,由河南農業大學牧醫工程學院草業科學實驗室培養提供,采集葉片后迅速放入液氮速凍后-80℃保存。
1.1.2菌株 大腸桿菌(Escherichia.coli)DH5α,農桿菌菌株GV3301,均由河南農業大學草業科學實驗室提供。
1.2 載體
克隆載體pMDl8-T(Takara);
植物表達載體pCAMBIA3301含有CaM 35 S啟動子和Nos polyA終止子(圖1)。
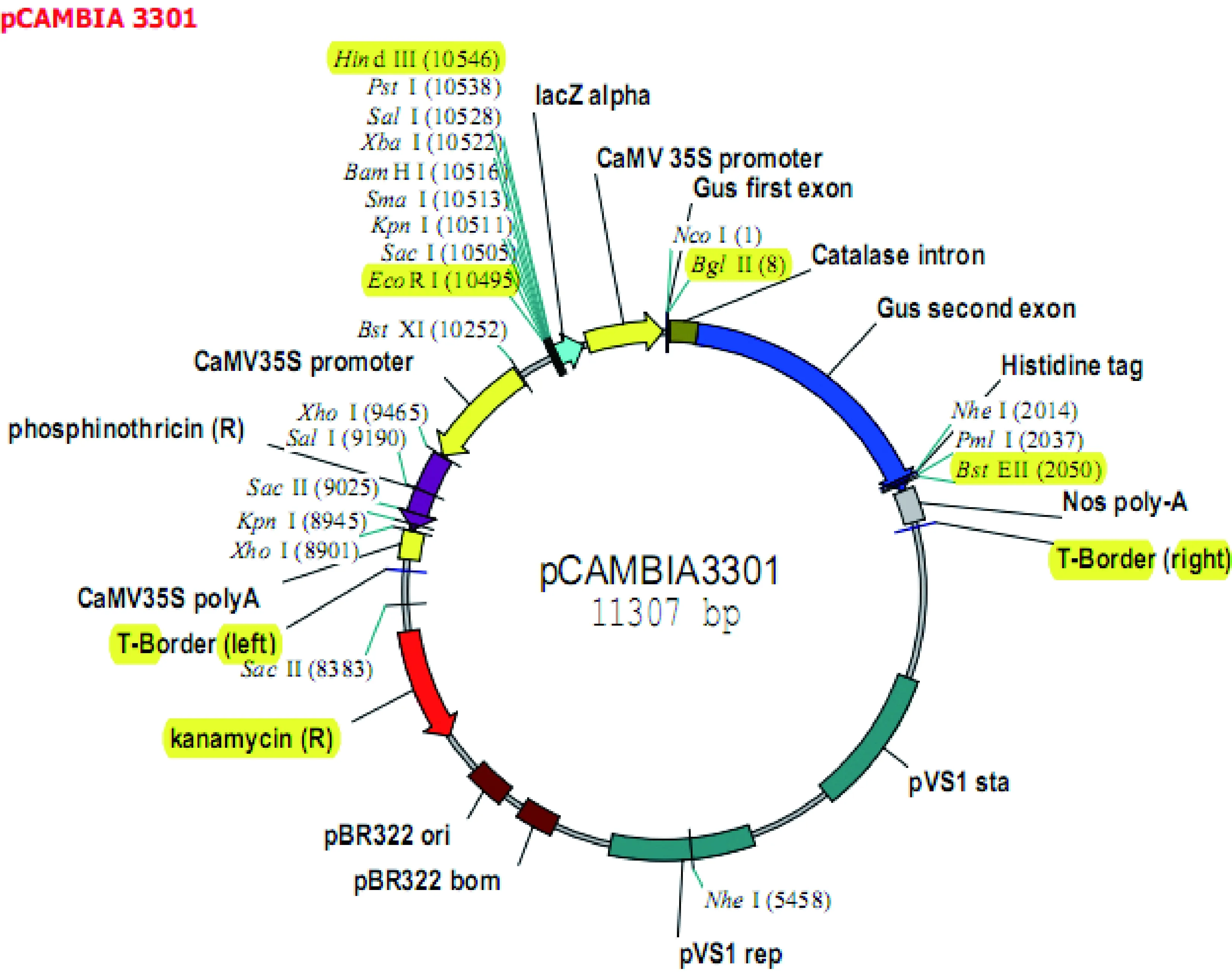
圖1 pCAMBIA3301結構圖Fig.1 Diagram of Pcambia3301
1.3 試驗方法
1.3.1CRY2B基因引物設計 引物設計前分析序列(紫花苜蓿基因組測序尚未完成,結合紫花苜蓿轉錄組測序信息,以模式植物蒺藜苜蓿CRY2B基因為模板 ,NCBI登錄ID:XM-003589988.1),基因序列編碼區696-2498(即CRY2B完整的開放閱讀框),設計引物擴增出來的目的片段要包含此閱讀框,如下:AATCTGAAAATAGGTATGAATAGGACCATAGTT-TGGTTTAGGAGGGACCTAA-GAATTGAGGACAA………TACCATAACAATGTTCCTCT-CCTCTTCTACTAATATAAAGTT-GTCATGGTAAGAACATAG(加粗部分劃線為CRY2B起始密碼子和終止密碼子,目的基因過表達要求引物設計的長度從起始密碼子和終止密碼子,含完整的開放閱讀框)。
利用Premier 5設計引物,選擇過表達載體上NcolI、PmlI為酶切位點(同時對植物表達載體pCAMBIA3301和目的基因CRY2B進行雙酶切),在設計好的CRY2B引物(696-2553)上加入酶切位點(劃線部分為限制性酶切位點),為保證酶切質量,酶切位點前面為保護堿基(表1) ,并送生工生物(上海)股份有限公司。

表1 基因引物序列Table 1 The primers of CRY2B gene
1.3.2CRY2B基因克隆與測序 PCR產物經1.5%瓊脂糖凝膠電泳檢測后回收目的片段,進行PMD18-T克隆轉化,菌液PCR檢測為陽性后送上海生工測序。測序結果在http://blast.ncbi.nlm.nih.gov/Blast.cgi進行比對,CRY2B克隆成功。用質粒提取試劑盒提取質粒,保存并命名為PMD18-T-CRY2B。
1.3.3純化回收的產物與PMD-18T載體的連接 采用北京百泰克生物技術有限公司瓊脂糖凝膠DNA回收試劑盒回收,嚴格按照回收試劑盒的操作步驟進行,回收到的產物置于-20℃冰箱中保存備用。
純化回收產物CRY2BPMD-18T載體連接前,使用BspHI/PmlI內切酶,分別雙酶切CRY2B目的片段和植物表達載體pCAMBIA3301,為提高CRY2B和pCAMBIA3301載體兩個目的片段的酶切效率和準確性,先將目的片段與PMD-18T載體連接,增加CRY2B目的片段的拷貝數。
DH5α感受態細胞轉化,用于擴繁、培養,進行PCR鑒定。
1.3.4提取陽性克隆DNA質粒 利用百泰克公司小量質粒提取試劑盒提取,嚴格按照提取質粒試劑盒的操作步驟進行。
1.3.5含有T載體的CRY2B片段質粒與pCAMBIA3301質粒雙酶切 在冰上進行操作,反應液混勻后37℃酶切反應1 h。
1%的瓊脂糖凝膠電泳觀測酶切效果,對酶切正確的目的片段進行切膠純化回收,以備連接反應。
1.3.6連接反應 將純化回收酶切后的CRY2B目的片段連接到酶切后的pCAMBIA3301上,連接反應目的片段與載體的濃度摩爾比例為1:6~10。
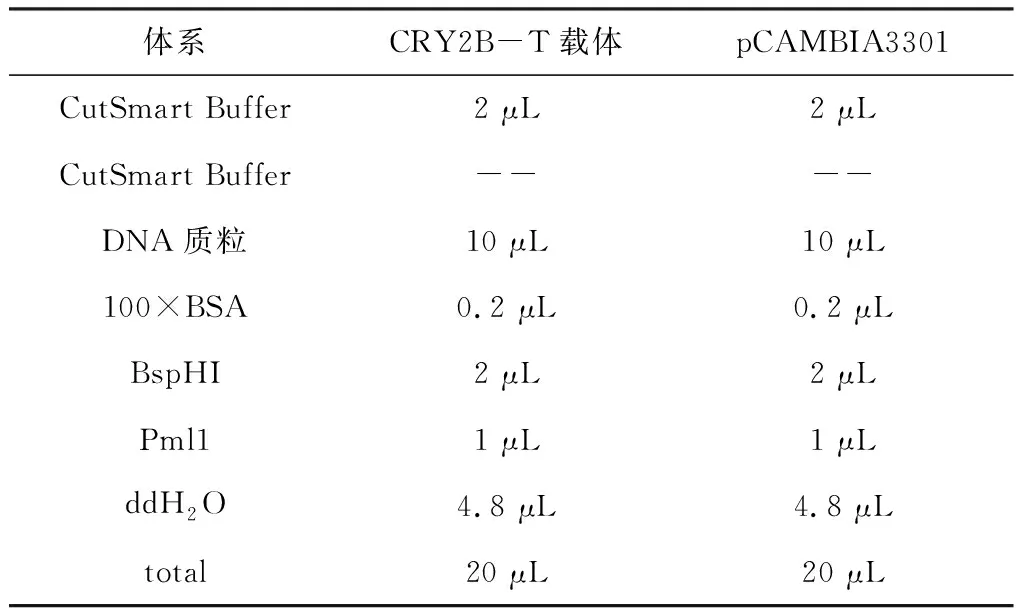
表2 雙酶切反應體系Table 2 The reaction system of restriction endonuclease
1.3.7連接產物轉化及鑒定 pCAMBIA3301載體與CRY2B目的片段連接產物轉化操作同上述DH5α感受態細胞轉化步驟,由于載體pCAMBIA3301含有Kan、Amp的特性,所以配制含有Kan、Amp特性的LB培養基,轉化后對含有目的片段菌液進行PCR檢測。PCR檢測為陽性克隆的菌液進行測序并提取質粒,長期保存,然后進行酶切檢測。
1.3.8酶切檢測 由于CRY2B基因上游添加的酶切位點是NcolI的同尾酶BspHI,因此重新整合的重組質粒不能用原來的酶切位點NcolI或PmlI去進行酶切鑒定,利用generunner分析可知,CRY2B基因片段上含有和pCAMBIA3301載體上相同的酶切位點EcoR1,此酶切位點不在pCAMBIA3301載體的表達區。利用EcorI進行酶切,若瓊脂糖凝膠電泳得到10 000 bp和2 000 bp大小的兩個片段(可根據pCAMBIA3301載體序列酶切位點的分析和CRY2B基因擴增長度判斷),則說明目的片段pCAMBIA3301-CRY2B重組質粒構建成功,如果沒有得到兩個片段,則質粒重組不成功。
1.3.9農桿菌介導pCAMBIA3301-CRY2B轉化苜蓿植株 質粒為上述試驗構建的MBIA3301-CRY2B過表達載體,首先制備農桿菌GV3101感受態細胞,然后選擇粒大飽滿的‘馴鹿’紫花苜蓿種子,使用清水沖洗30 min,然后75% 的酒精浸泡3 min進行消毒,棄溶液,然后用0.1%HgCl2(氯化汞)溶液滅菌15 min,再用無菌水清洗3~5次,將種子置于多層無菌濾紙上直至吸去多余水分,用已滅菌的鑷子將種子接種到MS培養基上(30 g·L-1蔗糖和2.5 g·L-1Phytagel)。將接種的培養基放入光照培養箱里,在白晝光周期為16 h/8 h,白晝溫度為22/20℃的條件下培養,采用直接分化芽再生途徑的方法進行農桿菌介導pCAMBIA3301-CRY2B重組載體轉化紫花苜蓿[12-16]。
1.3.9.1 農桿菌的侵染
(1)在含重組質粒pCAMBIA3301-CRY2B農桿菌的YEB(50 mg·L-1kan+100 mg·L-1rif)固體培養基上劃線培養,放入28℃培養箱培養48 h左右。
(2) 2 d后,用牙簽挑取單菌落,接種于含有50 mg·L-1kan和100 mg·L-1rif的20 mL YEB液體培養基中,28℃ 200~220 rpm震蕩培養16~24 h, OD600=0.5~0.7,用于外植體的侵染。
(3)將切割好的子葉放入準備好的農桿菌菌液中侵染60 min(隔10 min輕微晃動1次),使農桿菌能夠充分接觸子葉節的傷口。接著將侵染過的子葉轉移到干燥的無菌濾紙上,吸去表面多余的菌液,然后接種到共培養培養基上(光照培養箱24℃黑暗培養,3~4 d)。
1.3.9.2 芽誘導和篩選
共培養后的子葉轉入芽誘導培養基中,注意把生長點插入培養基以保證子葉的生長方向,不定芽直立向上生長,并且子葉節處的傷口能夠接觸到培養基[17-20]。培養條件為光周期:16 h/8 h,溫度:白晝25℃/22℃。轉化成功的植株會在子葉節的傷口處直接分化出不定芽,且不定芽出現后會迅速生長。未轉化成功的外植體會逐漸變黃、發褐甚至死亡。
1.3.9.3 誘導生根
選擇芽誘導長勢較好且已長有小根須的樣本,在無菌條件下轉入含有生根培養基(1/2MS+Yeastextract +carb)的三角瓶(瓶口較大,太小容易污染)中,用封口膜將錐形瓶口封緊,放入光照培養箱,設置培養條件,光照時間和溫度:16 h(25℃)/黑暗時間和溫度8 h(22℃)。誘導根生長30天左右,幼苗主根茁壯,接近于正常的植株。
1.3.9.4 煉苗移栽
當苜蓿轉基因植株根部產生多條粗而壯的根須時,可以進行煉苗,即對三角瓶內的轉基因幼嫩植株進行馴化,以適應外界生長條件[21]。打開三角瓶瓶口,加入一定量的滅菌去離子水(避免出現干燥應激)。在光照培養箱中生長4~6 d,將幼苗從三角瓶中取出,沖洗根部的培養基(一定要沖洗干凈,避免病菌滋生使根部壞死)。之后將轉基因苜蓿植株移栽于混有營養土/蛭石(1:1)的小花盆中。當苜蓿
植株長出健壯莖葉時,即可進行轉基因陽性苗的鑒定。
2 結果與分析
2.1 CRY2B克隆PCR擴增效果
根據克隆的基因以及所需的酶切位點設計CRY2B引物,進行PCR擴增,用1%的瓊脂糖凝膠電泳檢測。結果如圖2所示,目的片段條帶清晰明亮,可進行目的片段純化回收。
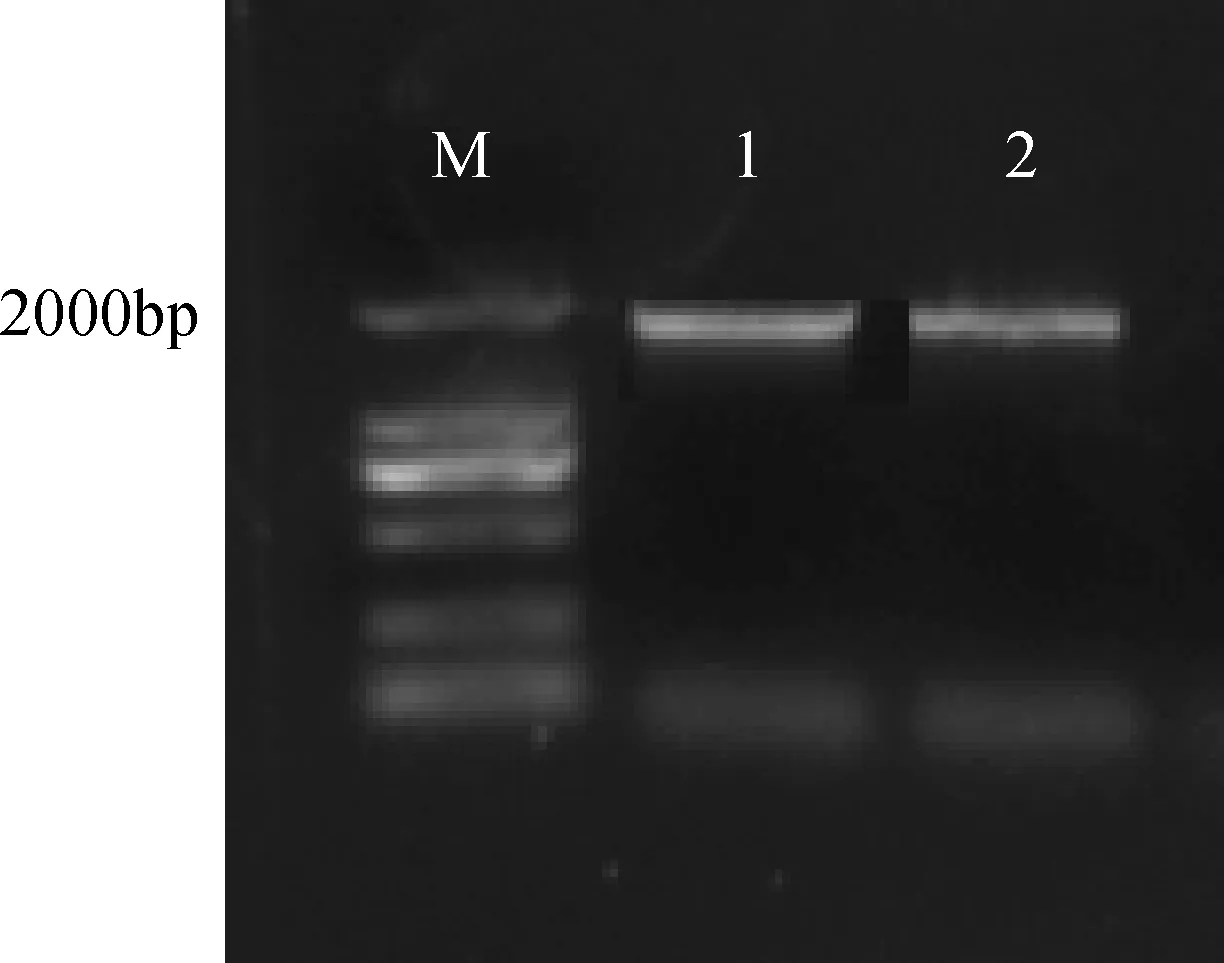
圖2 CRY2B PCR 擴增Fig.2 Electrophoresis of the product of CRY2B PCR amplificationM: DNA分子量標記;1-2: CRY2BM: DNA ladder; 1-2: CRY2B
2.2 pCAMBIA3301載體與CRY2B目的片段連接的菌液PCR檢測
將純化回收酶切后的CRY2B目的片段連接到酶切后的pCAMBIA3301連接產物,轉化到DH5α大腸桿菌過夜培養。根據克隆的CRY2B基因以及所需的酶切位點所設計的引物,以菌液為模板,進行PCR擴增檢測。擴增出如圖3清晰明亮的目的片段,說明目的片段已經成功連接到pCAMBIA3301載體上,可以進行測序鑒定與酶切反應。
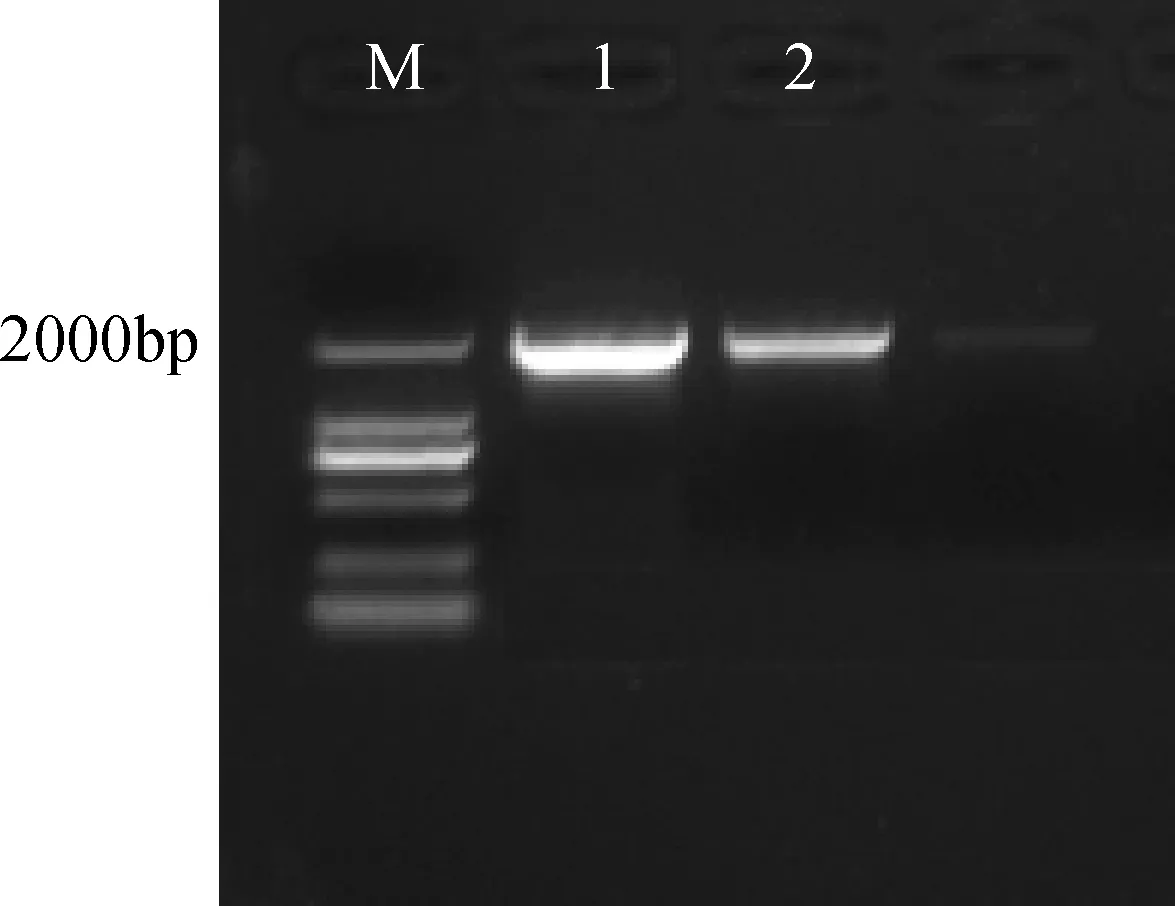
圖3 CRY2B 菌液PCR 擴增Fig.3 CRY2B bacteria PCR amplificationM: DNA分子量標記;1-2: CRY2BM: DNA ladder; 1-2: CRY2B
2.3 pCAMBIA3301載體與CRY2B目的片段連接產物的酶切效果與分析
pCAMBIA3301載體與CRY2B目的片段連接產物轉化操作同上述DH5α感受態細胞轉化,由于載體pCAMBIA3301含有Kan、Amp的特性,所以配制含有Kan、Amp特性的LB培養基,轉化后對含有目的片段的菌液進行PCR檢測,檢測為陽性克隆的菌液,然后進行EcoRI酶切鑒定檢測。如圖4酶切出現一個10 000 bp大小和一個2 000 bp大小的片段條帶。
對酶切鑒定出現正確片段的菌液送往生工生物工程(上海)股份有限公司測序,測序結果和目的片段序列一致(圖5),表明pCAMBIA3301-CRY2B重組質粒構建成功,保存pCAMBIA3301-CRY2B陽性克隆的菌液或提取陽性克隆菌液的質粒保存。
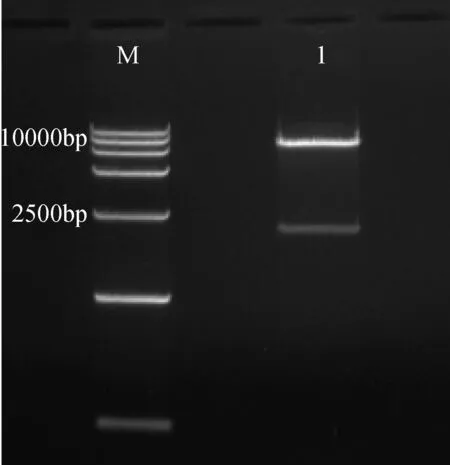
圖4 pCAMBIA3301-CRY2B質粒酶切鑒定Fig.4 pCAMBIA3301-CRY2B plasmid restrictionenzyme digestion analysis1: pCAMBIA3301-CRY2B 酶切;M: DNA 分子量標記;1 pCAMBIA3301-CRY2B restriction enzymeanalysis; M: DNA ladder
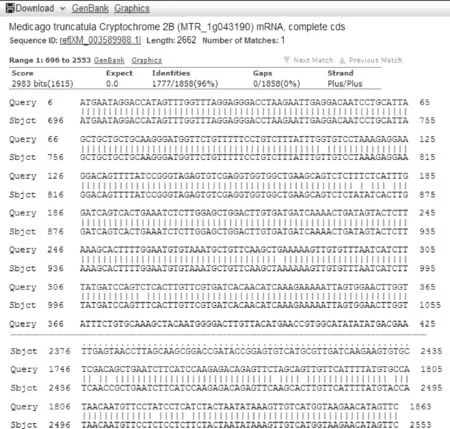
圖5 克隆片段與CRY2B基因片段序列比對Fig.5 Sequence alignment between cloning sequence and CRY2B fragment
2.4 重組質粒PCR檢測
將重組質粒pCAMBIA3301-CRY2B導入農桿菌GV3101感受態細胞,在含有50 mg·L-1Kan + 50 mg·L-1rif的YEB培養基上涂板,28℃培養48小時。以菌液為模板,進行PCR擴增,陽性菌落的PCR瓊脂糖凝膠電泳圖如圖6所示。挑選的單菌落都為陽性菌落,表明植物表達載體pCAMBIA3301-CRY2B已轉化進農桿菌GV3101中,可用于后期紫花苜蓿的遺傳轉化。
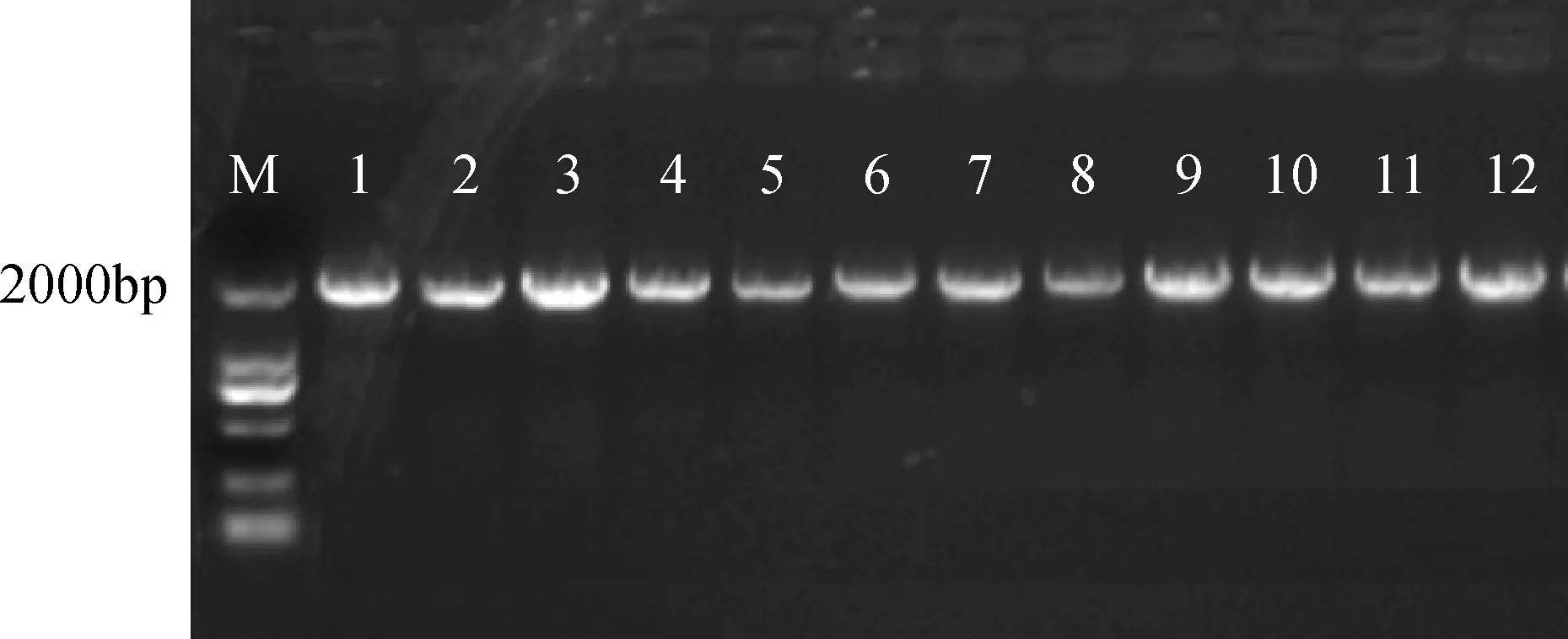
圖6 pCAMBIA3301-CRY2B質粒PCR擴增Fig.6 pCAMBIA3301-CRY2B plasmid PCR amplification1-12 CAMBIA3301-CRY2B PCR擴增;M: DNA 分子量標記;1-12 CAMBIA3301-CRY2B PCR amplification; M: DNA ladder
2.5 酶切鑒定重組質粒
利用上述菌液提取質粒進行酶切鑒定,選用pCAMBIA3301載體和CRY2B共有的酶切位點EcoRI,酶切鑒定結果如圖7所示,酶切出兩條正確的基因片段。
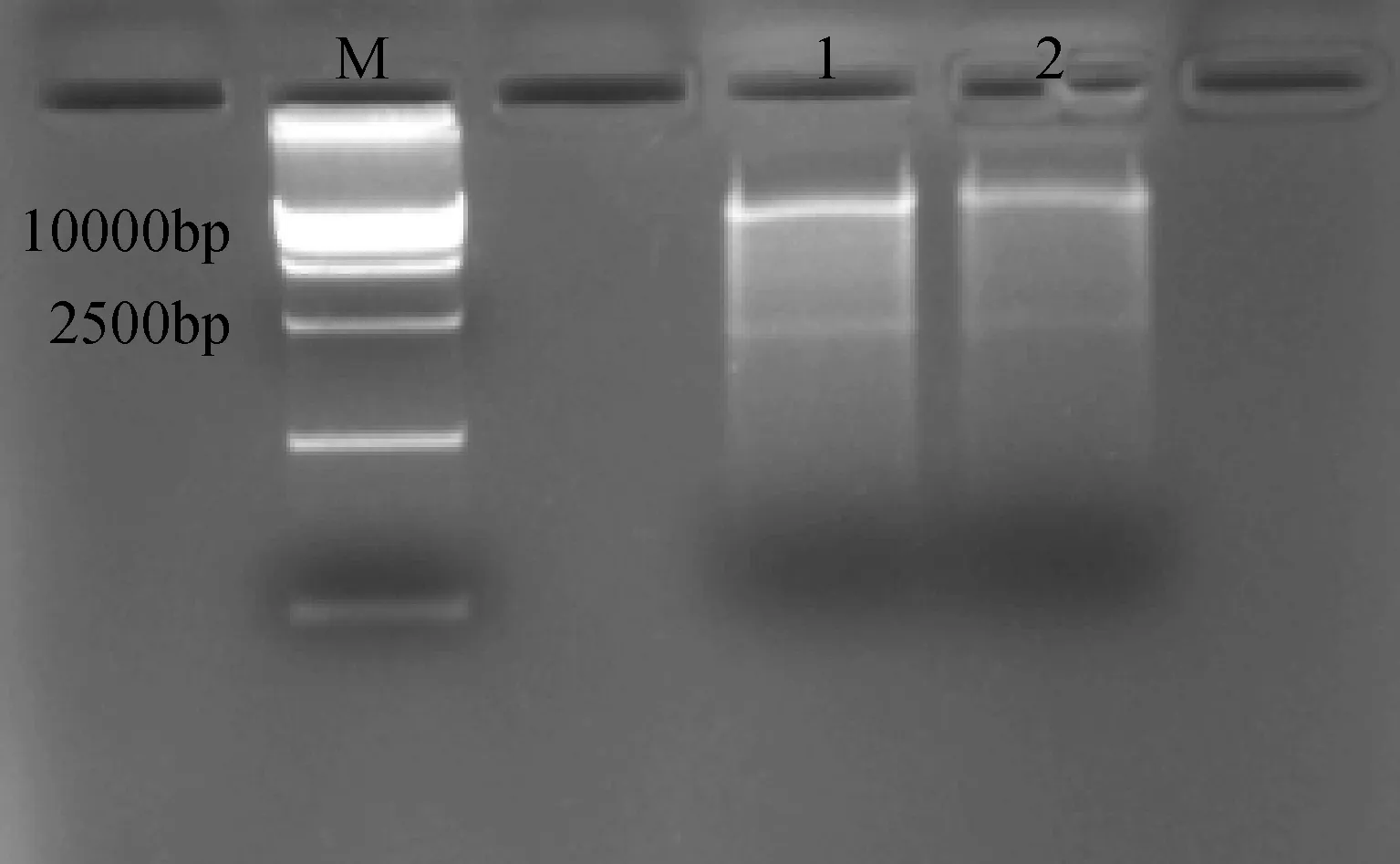
圖7 pCAMBIA3301-CRY2B農桿菌質粒酶切鑒定Fig.7 pCAMBIA3301-CRY2B Agrobacteriumplasmid restriction enzyme analysis1-2: pCAMBIA3301-CRY2B 酶切;M: DNA 分子量標記1-2:pCAMBIA3301-CRY2B restriction enzyme analysis; M: DNA ladder
2.6 pCAMBIA3301-CRY2B轉基因植株誘導芽的生長
以子葉節為外植體,適宜的Kan抗性篩選濃度為100 mg·L-1,適宜的子葉節芽誘導培養基為MS+1.0 mg·L-16-BA+0.1 mg·L-1NAA+100 mg·L-1Kan+400 mg·L-1Carb,pCAMBIA3301-CRY2B轉基因無菌苗子葉經侵染之后,共培養4天,在誘導芽培養基上進行芽誘導,14~18 d左右子葉長出新芽(圖8)。

圖8 誘導芽的生長Fig.8 The growth of induced shoots
2.7 pCAMBIA3301-CRY2B轉基因植株誘導根的生長
適宜的子葉節根誘導培養基為:1/2MS+0.2 g·L-1YE+100 mg·L-1Kan+400 mg·L-1Carb。pCAMBIA3301-CRY2B長出新芽的個體移入生根培養基中生長30~40 d,無菌苗苜蓿生根情況如圖9所示。

圖9 pCAMBIA3301-CRY2B轉基因植株誘導根的生長Fig.9 The growth of induced roots with transgenic-pCAMBIA3301-CRY2B
2.8 煉苗與再生植株移栽
當再生的芽生長健壯達到約4 cm時,即可轉移到生根培養基上進行生根誘導。待根部足夠發達,出現多條根時,開始煉苗。約3~5 d后,小心取出小植株,把根部沖洗干凈,種植到小花盆中。煉苗初期采用透明薄膜罩盆保持較高的濕度,待植株生長穩定后,即可摘除,仍保持濕潤環境,保證幼嫩轉基因植株較大的成活率(圖10)。

圖10 煉苗和再生植株移栽Fig.10 Plantlets exercising and transplanting of regenerated plantlets
3 討論與結論
對擬南芥(Arabidopsisthaliana)的研究發現,隱花色素和光敏色素具有拮抗和互補的作用[22],研究表明,光受體基因通過光周期參與了對苜蓿秋眠的調控[23-27],在對PHYA、PHYB沉默的轉基因植株研究中發現,CRY2B的表達和PHYB的表達近乎完全一致[23]。秋眠型紫花苜蓿CRY2B在夏季高溫休眠條件下基因表達量沒有發生變化,但秋眠條件下呈顯著增長[23],因此在對紫花苜蓿秋眠調控機制的研究中,CRY2B基因功能的研究至關重要。
本試驗CRY2B過表達載體構建成功后直接轉化紫花苜蓿。通過在CRY2B基因的引物兩端加入NcolI的同尾酶BspHI(使用同尾酶可產生相同的粘性末端)和PmlI位點,以提取的苜蓿RNA為模板進行PCR擴增得到CRY2B基因片段。選擇過表達載體上NcolI、PmlI作為雙酶切的酶切位點(載體上沒有BspHI酶切位點),NcolI酶切(A/CATGT),為了使氨基酸正確翻譯,CRY2B引物添加NcolI酶切位點時要使用同尾酶BspHI(T/CATGA)。然后將目的片段CRY2B和過表達載體pCAMBIA3301經過雙酶切,使其產生相同的黏性末端。將切下的目的基因CRY2B的片段插入質粒的切口處,首先堿基互補配對結合,兩個黏性末端吻合在一起,利用NEB公司T4連接酶分別將酶切后基因片段與酶切后pCAMBIA3301質粒整合到一起,構建pCAMBIA3301-CRY2B重組質粒,然后經過PCR檢測、堿基序列測序和酶切鑒定確定成功構建了pCAMBIA3301-CRY2B過表達載體,植物表達載體pCAMBIA3301-CRY2B已轉化進農桿菌GV3101中,可用于后續轉基因紫花苜蓿的遺傳轉化。
近年來,紫花苜蓿載體構建并對煙草進行轉化研究較多[28-31],但轉化方式不盡相同。本試驗采用直接分化芽再生途徑轉化紫花苜蓿,以萌發早期幼苗的子葉節為外植體,經農桿菌侵染后無需組織培養,外植體細胞不經過脫分化形成愈傷組織的階段,而是直接分化出不定芽,進而生長發育為完整植株。該途徑的優點是:植株再生周期短,遺傳變異較小,轉入的外源基因遺傳穩定性高。鄭寶仁等[32]以子葉節為外植體,在MS培養基上添加不同的激素組合直接進行不定芽的誘導,最終得到離體再生植株。Kumar等[33]研究發現對莖尖外植體可以直接進行不定芽的誘導(最多時一個外植體可以誘導出35個不定芽),然后再加入適當比例的NAA和活性炭即可誘導生根進而得到完整植株,并且該流程能夠適用于5個印度苜蓿品種。本試驗以紫花苜蓿子葉節為外植體,利用農桿菌介導pCAMBIA3301-CRY2B成功轉化苜蓿植株,為進一步研究CRY2B調控苜蓿秋眠性分子機制奠定基礎。
