青海海北3種典型高寒草甸植物群落特征研究
2017-09-14 09:10:34井向前王瑞紅任德智白瑪玉珍周堯治
草地學報 2017年1期
關(guān)鍵詞:物種
井向前,王瑞紅,任德智,白瑪玉珍,潘 剛,周堯治*
(1.高原生態(tài)研究所,西藏 林芝860000 2.西藏大學農(nóng)牧學院,西藏 林芝860000)
高寒草甸是適應(yīng)高原高寒環(huán)境條件而形成的植被類型,其在青藏高原具有很強的典型性[1],具有雙重地帶性的典型植被,即高原地帶性和山地垂直地帶性[2],也是畜牧業(yè)的主要物質(zhì)基礎(chǔ)。近年來由于畜牧業(yè)的發(fā)展,高寒草甸超載放牧,導(dǎo)致草地生態(tài)系統(tǒng)發(fā)生不同程度的惡化,表現(xiàn)為草地退化面積增大,物種多樣性減少,草甸初級生產(chǎn)力下降,草地生態(tài)環(huán)境日益惡化[3]。目前有關(guān)高寒草地多樣性的研究多集中在草地荒漠化和不同草地植物群落特征[4]及不同放牧強度下高寒草地的退化過程等方面。隨著放牧強度的增加,物種多樣性和豐富度指數(shù)逐漸降低,草地初級生產(chǎn)力也隨之降低。植被群落特征是反映草地生產(chǎn)狀況的重要指標,尤其是植物多樣性和初級生產(chǎn)力是權(quán)衡草地生產(chǎn)狀況及其趨勢的重要因子。本文通過對高寒草地不同植物群落特征及多樣性進行比較,旨在了解高寒草地不同植物群落的植被現(xiàn)狀,為研究區(qū)域內(nèi)不同草地植物群落保護和進一步利用提供依據(jù)。
1 材料與方法
1.1 研究區(qū)概況
門源縣位于中國青海省東北部,屬海北藏族自治州管轄區(qū)域,在海北藏族自治州東部,面積為6896km2。青海省門源縣門源馬場,N 37.5°~38°,E 102°~102.5°。區(qū)內(nèi)以高原亞寒帶氣候為主,干濕季分明,冬季寒冷干燥,夏季涼爽濕潤。全年日照時數(shù)2264.8~2739.8h,氣溫日較差11.6~17.5℃,年平均氣溫0.8℃,年平均降水520mm。
1.2 試驗設(shè)計及調(diào)查地點
在門源馬場干柴灘、風匣口南灘和口門子分別選擇1個草地,共3個,于2010年植物生長季后期進行調(diào)查,每個草地各設(shè)5個調(diào)查樣地,共15個。根據(jù)草地情況,在每個樣地設(shè)置一條100m樣線,沿樣線設(shè)置10個1m×1m的小樣方,共計150個小樣方。植被調(diào)查以樣方為單位,估測整個群落的蓋度,同時用鋼卷尺測量群落內(nèi)植物的高度,并統(tǒng)計植物的多度;剪取地上生物量,裝入自封袋,測定鮮重。將取回的地上生物量在烘箱內(nèi)以65℃烘干至恒重,稱取干重。地下生物量的測定采用土柱法取樣。地下生物量樣方設(shè)在地上的生物量樣方內(nèi),共3次重復(fù)。每個樣方分4層取樣,分別為0~10cm,10~20cm,20~30cm,30~40cm。樣品按層標注裝入土壤袋中,帶回實驗室沖洗、分離,然后在65℃烘箱內(nèi)烘干至恒重,及時稱其干重。
1.3 多樣性指數(shù)計算
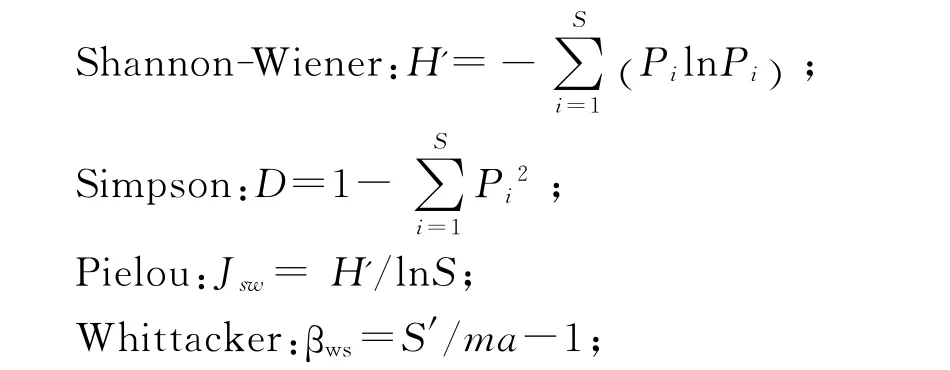
式中Pi為群落內(nèi)第i種物種的重要值,S為樣地中的物種數(shù)。ma為各樣方或樣本的平均物種數(shù)。
草本植物重要值計算公式:Pi=(相對蓋度+相對高度+相對多度)/3。
1.4 數(shù)據(jù)分析
數(shù)據(jù)來源于中國科學院海北高寒草甸生態(tài)系統(tǒng)定位研究站2010年調(diào)查數(shù)據(jù),采用origin9.0軟件進行地上、地下生物量分布分析。
2 結(jié)果與分析
2.1 不同高寒草甸植物群落的組成與結(jié)構(gòu)
青海海北不同高寒草甸群落組成存在明顯差異,矮嵩草草甸植物群落的優(yōu)勢種為垂穗披堿草(Elymusnutans)和矮嵩草(Kobresiahumilis);金露梅草甸植物群落的優(yōu)勢種為金露梅(Potentillafruticosa)和溚草(Koeleriasp.);門源嵩草草甸群落的優(yōu)勢種為門源嵩草(Kobresiamenyuanica)和矮嵩草(Kobresiahumilis)。矮嵩草草甸植物伴生種以禾本科的優(yōu)良牧草羊茅(Festucaovina)為主,金露梅草甸以莎草科的優(yōu)良牧草線葉嵩草(Kobresiacapillifolia)居多,門源嵩草草甸以禾本科的異針茅(Stipaaliena)為主,亦為優(yōu)良牧草(表1)。植物物種數(shù)表現(xiàn)為矮嵩草草甸>金露梅草甸>門源嵩草草甸。3種群落間植被高度差異較大,表現(xiàn)為矮嵩草草甸>金露梅草甸>門源嵩草草甸;3種草甸群落蓋度均大于90%,群落垂直結(jié)構(gòu)均為復(fù)層。
2.2 青海海北高寒草甸植物群落α和β多樣性
青海海北不同類型高寒草甸植物群落多樣性存在明顯差異(表2)。均勻度指數(shù)表現(xiàn)為矮嵩草草甸和門源嵩草草甸明顯大于金露梅草甸。3種高寒草甸植物群落多樣性指數(shù)與均勻度指數(shù)的相關(guān)系數(shù)分別為0.6502,0.9371和0.8729;多樣性指數(shù)與豐富度指數(shù)的相關(guān)系數(shù)分別是-0.7214,-0.7259和-0.7081,3種高寒草甸植物群落的β多樣性指數(shù)均無顯著性差異。

表1 青海海北高寒草甸植物群落特征Table 1 Community characteristics of alpine meadow in Haibei region of Qinghai province

表2 青海海北高寒草甸植物群落α和β多樣性Table 2 αandβdiversity of alpine meadow communities in Haibei region of Qinghai province
2.3 不同高寒草甸植物群落地上生物量分布趨勢
青海海北不同高寒草甸群落的地上生物量差異較為明顯(圖1),矮嵩草高寒草甸的地上生物量遠大于金露梅高寒草甸和門源嵩草高寒草甸(P<0.05),整體表現(xiàn)為矮嵩草高寒草甸地上生物量分別是金露梅高寒草甸和門源嵩草高寒草甸的1.62和1.69倍。
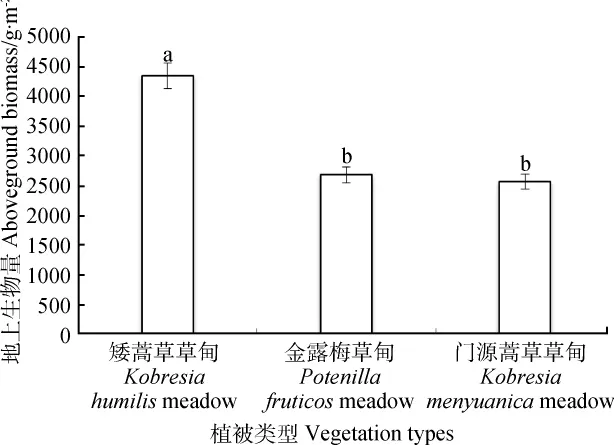
圖1 青海海北高寒草甸植物群落地上生物量Fig.1 Aboveground biomass of alpine meadow in Haibei region of Qinghai province
2.4 不同高寒草甸植物群落地下生物量的垂直分布特征
青海海北不同高寒草甸植物群落的地下生物量隨土層深度的變化而變化(表3),其地下生物量主要分布在土壤表層(0~10cm)。矮嵩草草甸的地下生物量為2842.62g·m-2,其中0~10cm土層生物量約占地下總生物量的77.44%。金露梅草甸的地下生物量為3241.79g·m-2,0~10cm土層生物量約占地下總生物量的54.02%。門源嵩草草甸地下生物量為4740.77g·m-2,其中0~10cm土層生物量約占地下總生物量的78.73%。他們的垂直空間分布結(jié)構(gòu)表現(xiàn)為總的呈T字形趨勢,鋸齒狀分布(圖2)。
2.5 不同高寒草甸植物群落地上、地下生物量之間的關(guān)系
由表4可知,矮嵩草草甸、金露梅草甸和門源嵩草草甸的地上和地下生物量呈負相關(guān)關(guān)系。3種草甸群落地下、地上生物量的比值分別為0.65,1.21和1.85。
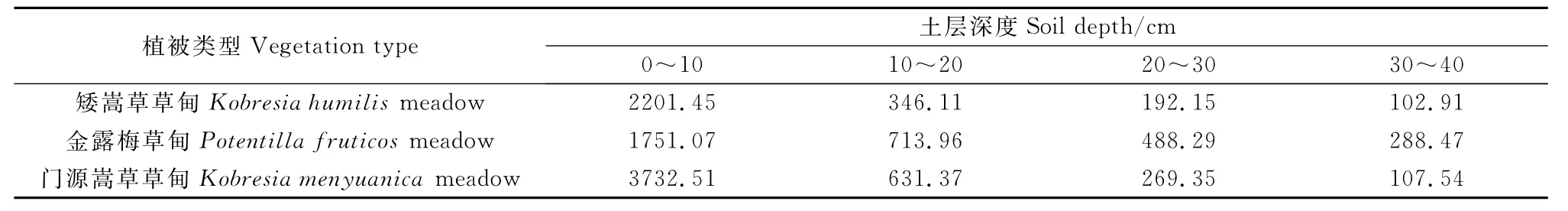
表3 高寒草甸植物群落不同土壤深度地下生物量Table 3 The belowground biomass of different soil depths of alpine meadow communities/g·m-2
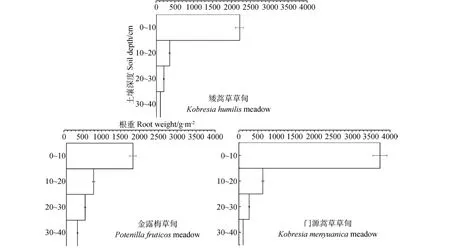
圖2 青海海北地區(qū)主要植物群落地下生物量垂直分布Fig.2 The vertical distribution of belowground biomass of dominant vegetation communities in Haibei region of Qinghai province

表4 不同草甸植物群落地上、地下生物量相關(guān)性Table 4The correlation of aboveground and belowground biomass of communities
3 討論
高寒草甸群落特征和生產(chǎn)力均隨環(huán)境的變化而變化[5]。青海海北地區(qū)門源嵩草草甸植物群落上層為禾本科牧草,下層為嵩草;而金露梅草甸植物群落上層為金露梅,下層為苔蘚,3種高寒草甸植物群落呈明顯的雙層結(jié)構(gòu),3種高寒草甸的氣候特征具有一定的相似性,但其初級生產(chǎn)力差異較大,主要原因是植物群落的優(yōu)勢種和主要伴生種存在差異。因此,應(yīng)因地制宜地制訂青海海北地區(qū)不同草地類型經(jīng)營策略。
本文采用α多樣性指數(shù)測定青海海北地區(qū)高寒草甸的植物多樣性,發(fā)現(xiàn)植物多樣性指數(shù)和均勻度指數(shù)均表現(xiàn)為矮嵩草和門源嵩草草甸植物群落大于金露梅草甸植物群落,該地區(qū)的物種既包括高原高海拔地區(qū)特有的物種,如委陵菜屬植物,又有喜濕、熱的植物,這是由于緯度,水熱條件和海拔等過渡性的生境條件為多物種的生存和發(fā)展提供了資源基礎(chǔ)。青海海北地區(qū)不同高寒草甸植物群落間共有物種較少,物種替代速率較高。
青海海北地區(qū)高寒草甸植物群落地下生物量占總生物量的比例非常大,且地下、地上生物量的比值越大,地上生物量越低。矮嵩草草甸植物群落較高的生物多樣性維持了較高的初級生產(chǎn)力,這與Isbell等[6]的研究結(jié)果具有一定的一致性,即植物物種的多樣性是草地生態(tài)系統(tǒng)維系高產(chǎn)服務(wù)的支持者。
4 結(jié)論
青海海北地區(qū)3種典型高寒草甸植物群落的垂直結(jié)構(gòu)均為復(fù)層;其多樣性指數(shù)與均勻度指數(shù)關(guān)系較其與豐富度指數(shù)的關(guān)系更為密切;不同高寒草甸植物群落內(nèi)樣方間共有物種較少,物種替代速率較高;矮嵩草高寒草甸地上生物量遠高于金露梅高寒草甸和門源嵩草高寒草甸;其地下生物量主要分布在土壤表層,垂直空間分布結(jié)構(gòu)呈T字形和鋸齒狀;其地下生物量所占總生物量的比例非常大,且地下、地上生物量呈負相關(guān)關(guān)系。
猜你喜歡
課堂內(nèi)外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內(nèi)外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
課堂內(nèi)外·初中版(科學少年)(2024年12期)2024-12-02 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
中學生博覽(2022年7期)2022-06-21 21:48:14
大科技·百科新說(2021年8期)2021-11-03 10:55:16
學苑創(chuàng)造·A版(2021年5期)2021-06-28 19:51:42
少兒美術(shù)(快樂歷史地理)(2020年9期)2020-03-19 05:10:56
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
