異槲皮苷對HepG2細胞中Raf/MEK/ERK信號通路的干預作用
2017-09-23 03:25:06蔣國君劉天旭黃桂紅陶麗群朱釗銘
中國藥理學通報 2017年10期
蔣國君,劉天旭,黃桂紅,巫 亭,陶麗群,朱釗銘
(桂林醫學院1.臨桂臨床醫學院,2.第二附屬醫院藥學部,廣西 桂林 541199)
異槲皮苷對HepG2細胞中Raf/MEK/ERK信號通路的干預作用
蔣國君1,劉天旭1,黃桂紅2,巫 亭1,陶麗群2,朱釗銘2
(桂林醫學院1.臨桂臨床醫學院,2.第二附屬醫院藥學部,廣西 桂林 541199)
目的研究異槲皮苷對HepG2細胞中Raf/MEK/ERK信號通路的干預作用。方法采用異槲皮苷(0、40、80、160、320 μmol·L-1)作用于HepG2細胞,MTT法檢測異槲皮苷對HepG2細胞增殖的影響;倒置顯微鏡下觀察24、48 h后,細胞形態及生長情況;流式細胞術檢測異槲皮苷(0、40、80、160 μmol·L-1)作用HepG2細胞48 h后細胞周期情況;熒光定量PCR及Western blot檢測異槲皮苷作用HepG2細胞后Ras、Raf、MEK、ERK mRNA及相關蛋白的表達。結果MTT檢測發現異槲皮苷對HepG2細胞生長有明顯抑制作用,且與異槲皮苷的濃度及作用時間呈正相關;不同濃度的異槲皮苷作用HepG2細胞24、48 h后,倒置顯微鏡下觀察發現隨著濃度的增高及作用時間的延長,細胞生存數量逐漸減少,且細胞形態發生明顯變化;流式細胞術檢測發現隨著異槲皮苷濃度的增高,細胞被阻滯在G1期的數量逐漸增加;熒光定量PCR及Western blot結果均表明,與空白組相比,異槲皮苷(80 μmol·L-1)作用HepG2細胞后Ras、Raf、MEK、ERK mRNA及其相關蛋白的表達均明顯降低(P<0.05),差異有統計學意義。結論異槲皮苷有誘導HepG2細胞凋亡的作用,其作用機制可能與對Raf/MEK/ERK信號通路中相關因子的干預有關。
異槲皮苷;誘導;HepG2細胞;Raf/MEK/ERK;信號通路;凋亡
肝癌是一種常見的世界性惡性腫瘤,發病率與病死率極高,危害性極大。目前,臨床上針對肝癌的治療手段主要包括外科手術、化療、放療以及肝移植等,這些治療手段都有明顯缺陷,如預后差、副作用大[1]。因此,肝癌病人迫切需要尋找具有有效抗癌作用的新型治療藥物。異槲皮苷存在于大量的中草藥、植物及食物當中,具有抗氧化、降血壓、抗炎、抗腫瘤及抗惡性細胞增殖的作用[2-6]。課題組前期研究發現異槲皮苷具有誘導癌細胞凋亡的功能,但其具體的分子機制尚未完全明確。本實驗擬從細胞水平探討異槲皮苷對肝癌細胞抑制作用的分子機制,為異槲皮苷治療肝癌的研究提供新的理論依據。
1 材料與方法
1.1材料人肝癌HepG2細胞購自中科院上海細胞研究所;異槲皮苷購自Spring&autumn公司(純度≥98%,批號:482-35-9);Ras、Raf、p-Raf、MEK、p-MEK、ERK、p-ERK、β-actin抗體購自Abcam公司;熒光定量PCR試劑(批號:RR420A)、TRIzol(批號:9108)購自日本TaKaRa公司;血清(胎牛)購自ExCell Bio公司(批號:11E074);DMEM培養基購于Gibco公司(批號:8114351);Ras、Raf、MEK、ERK、β-actin上下游引物和探針由InvitrogenTM公司設計并合成。
1.2方法
1.2.1MTT檢測異槲皮苷對HepG2細胞增殖的影響 取對數生長期細胞,接種于96孔板中(每孔2×103個細胞),置37℃、5% CO2條件下培養24 h后,吸棄上清,用培養基將異槲皮苷按下列濃度稀釋,制成0、40、80、160、320 μmol·L-1,每組設6個復孔,分別培養24、48、72 h,在檢測前4 h,每孔加入制備好的MTT溶液20 μL,置37℃、含5% CO2的飽和濕度培養箱避光繼續孵育,4 h后棄去各孔上清液,加入100 μL DMSO,振蕩10 min后,將96孔板置于酶標儀450 nm波長下檢測,測定各孔OD值。實驗重復3次,取平均值,按公式分析實驗結果:細胞生長抑制率IR/%=(OD藥物組- OD0組)/(OD空白組-OD0組)×100%,求出半數致死率IC50。
1.2.2細胞形態學觀察 取對數生長期細胞,以每孔1×106個細胞接種于6孔板中,置37℃、5% CO2條件下培養24 h后,吸棄上清,用培養基將異槲皮苷按下列濃度稀釋,制成0、40、80、160、320 μmol·L-1,分別培養24、48 h后,置于倒置顯微鏡下觀察細胞形態及生長情況。
1.2.3流式細胞數檢測細胞周期 取對數生長期細胞,以每孔1×106個細胞接種于6孔板中,置37℃、5% CO2條件下培養24 h后,分別加入異槲皮苷,使其終濃度為0、40、80、160 μmol·L-1,培養48 h后,用0.25%胰酶收集細胞,1 000 r·min-1離心5 min,棄上清,加入預冷的0.1 mol·L-1PBS洗滌1次,以1 000 r·min-1離心5 min,棄上清后,使用1 mL PBS重懸,加入2 mL預冷的無水乙醇固定30 min。上機前收集所有固定標本,離心棄上清,用PBS清洗1次,加入PI染液,室溫下避光染色30 min,采用流式細胞儀進行細胞周期檢測。
1.2.4熒光定量PCR檢測Ras、Raf、MEK、ERK相關基因mRNA表達 取對數生長期細胞,以每孔1×106個細胞接種于6孔板中,置37℃、5% CO2條件下培養24 h后,分別加入異槲皮苷及空白培養基,繼續培養48 h后收集細胞。TRIzol一步抽提法提取總RNA,取1 μg總RNA逆轉錄成cDNA,采用15 μL反應體系進行PCR擴增。引物序列如下:Ras上游引物: 5′-GGCAAGAGTGCCTTGACGAT-3′, 下游引物: 5′-TTGACCTGCTGTGTCGAGAA-3′; Raf上游引物:5′-CTCCTTGAATCGGGCTGGTT-3′, 下游引物: 5′-TGGACAGGAAACGCACCATA-3′; MEK上游引物: 5′-TGTGAAGGCGCTGTACTACC-3′, 下游引物: 5′-CAGGATGTTGGAGGGCTTGA-3′; ERK: 上游引物5′-GGCTGTTCCCAAATGCTGAC-3′, 下游引物:5′-TTGAATGGTGCTTCGGCGAT-3′; 內參β-actin作為對照, 上游引物: 5′-CCACTCCTCCACCTTTGAC-3′, 下游引物: 5′-ACCCTGTTGCTGTAGCCA-3′。反應體系包括SYBR 7.5 μL,H2O 6.9 μL,cDNA 0.2 μL,引物0.4 μL。反應條件為:95℃,30 s;95℃,5 s→60℃,30 s,40個循環;65℃→95℃,5 s,0.5℃/循環。
1.2.5Western blot檢測Ras、Raf、MEK、ERK相關蛋白的表達 細胞培養及干預作用同上,作用48 h后收集細胞,提取2組細胞的總蛋白,并用BCA法測定蛋白濃度,加熱變性后-20℃保存。取各組細胞的總蛋白質70 μg進行SDS-PAGE凝膠電泳,將分離后的蛋白質電轉移到PVDF膜,封閉液封閉濾膜4 h,加入適量一抗(Ras:1 ∶50 000,Raf:1 ∶5 000,p-Raf:1 ∶5 000,MEK:1 ∶20 000,p-MEK:1 ∶20 000,ERK:1 ∶10 000,p-ERK:1 ∶10 000,β-actin:1 ∶1 000)4℃過夜,TBST洗膜3次,每次10 min,二抗(1 ∶5 000)室溫孵育2 h,TBST洗膜3次,每次10 min,按1 ∶1加入AB發光液,在Bio-Rad的化學發光成像儀上成像,用圖像分析軟件對條帶的灰度值進行分析,分析出Ras、Raf、MEK、ERK相關蛋白的表達。
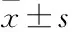
2 結果


Fig 1 The growth effect of HepG2 cells treated with isoquercitrin
2.2異槲皮苷對HepG2細胞形態的影響如Fig 2所示,倒置顯微鏡下觀察發現,當異槲皮苷作用細胞時間一定時,隨著藥物濃度的升高,細胞形態發生變化,與空白組相比,隨著濃度的升高,可見形態發生變化的細胞數量逐漸增多,且存活的細胞數量逐漸減少,即貼壁的長梭形細胞減少,而懸浮的圓形細胞逐漸增多;當藥物濃度一定時,隨著異槲皮苷作用時間的延長,可見的形態發生變化的細胞數量也逐漸增多,且存活的細胞數量逐漸減少。
2.3異槲皮苷對HepG2細胞周期的影響Fig 3流式細胞儀檢測細胞周期結果顯示,與空白對照組相比,隨著藥物濃度的增加,細胞周期被阻滯在G1期的細胞百分比值逐漸增加(P<0.05),差異有統計學意義。
2.4異槲皮苷對HepG2細胞中Ras、Raf、MEK、ERKmRNA表達的影響Fig 4熒光定量PCR結果顯示,與空白組相比,異槲皮苷組Ras、Raf、MEK、ERK mRNA表達明顯降低(P<0.05,P<0.01),差異有統計學意義。

Fig2IsoquercitrininhibitsHepG2cellproliferationobservedbyinvertedmicroscopy(n=3)
A: The morphological changes of HepG2 cells treated with 0, 40, 80, 160 or 320 μmol·L-1of isoquercitrin after 24 h or 48 h were observed by inverted microscopy; B: Percentage of cell proliferation after isoquercitrin(0, 40, 80, 160 or 320 μmol·L-1) treated HepG2 cells for 24 and 48 h.*P<0.05,**P<0.01vscontrol

Fig 3 Effect of isoquercitrin on cell cycle distribution in HepG2 cells(n=3)
A: Control; B: Isoquercitrin 40 μmol·L-1; C: Isoquercitrin 80 μmol·L-1; D: Isoquercitrin 160 μmol·L-1; E: Isoquercitrin(0, 40, 80, 160 μmol·L-1) blocks HepG2 cell cycle in the G1phase.*P<0.05vscontrol

Fig 4 Isoquercitrin inhibits Ras, Raf, MEK,ERK mRNA expression levels in HepG2 cells(n=3)
*P<0.05,**P<0.01vscontrol
2.5異槲皮苷對HepG2細胞中Ras、Raf、MEK、ERK及相關蛋白表達的影響Fig 5 Western blot結果顯示,與空白組相比,異槲皮苷組Ras、p-Raf/Raf、p-MEK/MEK、p-ERK1/ERK1蛋白表達的相對比值明顯降低,表明異槲皮苷對HepG2細胞中Ras、Raf、MEK、ERK相關蛋白的表達具有下調作用(P<0.05,P<0.01),差異有統計學意義。
3 討論
中草藥因其資源豐富、毒副作用小、能夠調節機體代謝等優點已成為國內外關注的焦點,中草藥抗腫瘤治療也越來越受到人們的關注[7]。異槲皮苷可以從多種中草藥中提取,如鬼針草、荷葉、紅景天以及桑葉等,也可化學合成制得[8-10]。因其廣泛的生物活性,如抗氧化、改善心肌功能、抗腫瘤、調節血壓、降血脂等作用而備受關注,本課題前期研究也發現異槲皮苷具有誘導癌細胞凋亡的功能。

Fig 5 Isoquercitrin inhibits protein expression levels of Ras, Raf, MEK, ERK in HepG2 cells(±s,n=3)
*P<0.05,**P<0.01vscontrol
Ras是小分子量G蛋白家族成員之一,參與細胞的生長、增殖及分化,還影響細胞的生長形態發生及凋亡,活化的受體酪氨酸激酶能夠激活Ras蛋白,發生蛋白激酶的級聯反應,從而傳送到細胞核,這條通路可以由生長因子、細胞因子、射線、滲透壓以及體液流過細胞表面時產生的切應力等多種因素激活,被稱為絲裂原活化蛋白激酶(mitogenic activated protein kinase,MAPK)通路[11]。MAPK系統還包括ERK、MEK及Raf 3種因子,Raf是MAPK級聯反應的第1個分子,它的活化可引發蛋白質磷酸化級聯反應,而MEK可以使位于其下游的蘇氨酸和酪氨酸殘基磷酸化而激活,即Raf作用于MEK,使之磷酸化而活化,活化的MEK再作用于屬于MAPK家族的ERK,使其磷酸化而活化,從而介導炎癥及惡性細胞增殖[12]。且已有研究報道顯示,肝癌患者Raf 及其下游基因 MEK、ERK 的表達均上調,Raf存在過度表達現象,肝癌患者Raf水平明顯高于正常人的表達水平[13]。
本實驗發現, 異槲皮苷對HepG2細胞的具有抑制作用,且隨著藥物濃度的增加及作用時間的延長,抑制作用逐漸增強。異槲皮苷作用HepG2細胞48 h后, 倒置顯微鏡下觀察到HepG2的典型凋亡細胞,且隨著藥物濃度的增加,被阻滯在G1期的細胞百分比值逐漸增大。RT-PCR 和 Western blot 實驗顯示,與空白對照組相比,異槲皮苷能夠明顯降低ERK、MEK、Raf、Ras mRNA表達,并使ERK1、p-ERK1、p-MEK、Raf、p-Raf 及Ras蛋白表達降低,差異均具有統計學意義。提示異槲皮苷治療肝癌的機制與誘導肝癌細胞凋亡有關,其中,通過MAPK信號轉導下調Ras、Raf、MEK、ERK mRNA及相關蛋白的表達可能是異槲皮苷誘導肝癌細胞凋亡的重要機制之一。本實驗顯示異槲皮苷具有較好的誘導肝癌細胞凋亡的作用,在治療肝癌方面具有較好的應用前景。
(致謝:本實驗在桂林醫學院附屬醫院肝臟疾病研究室完成。)
[1] Van Meer S, de Man R A, Siersema P D, et al. Surveillance for hepatocellular carcinoma in chronic liver disease: evidence and controversies[J].WorldJGastroenterol, 2013,19(40):6744-56.
[2] Wu P, Li F, Zhang J, et al. Phytochemical compositions of extract from peel of hawthorn fruit, and its antioxidant capacity, cell growth inhibition, and acetylcholinesterase inhibitory activity[J].BMCComplementAlternMed, 2017,17(1):151.
[3] Li H, Liu Y, Yi Y, et al. Purification of quercetin-3-O-sophoroside and isoquercitrin from Poacynum hendersonii, leaves using macroporous resins followed by Sephadex LH-20 column chromatography[J].JChromatogrBAnalytTechnolBiomedLifeSci, 2017,1048:56-63.
[4] Ge L, Chen A, Pei J, et al. Enhancing the thermostability of α-L-rhamnosidase from Aspergillus terreus and the enzymatic conversion of rutin to isoquercitrin by adding sorbitol[J].BMCBiotechnol, 2017,17(1):21.
[5] Dawbaa S, Aybaster ?, Demir C. Ultrasensitive determination of DNA oxidation products by gas chromatography-tandem mass spectrometry and the role of antioxidants in the prevention of oxidative damage[J].JChromatogrBAnalytTechnolBiomedLifeSci, 2017,1051:84-91.
[6] Chen Q, Li P, Li P, et al. Isoquercitrin inhibits the progression of pancreatic cancerinvivoandinvitroby regulating opioid receptors and the mitogen-activated protein kinase signalling pathway[J].OncolRep, 2015,33(2):840-8.
[7] 劉慧穎,姜長濤,馮 娟,等.丹參酮類化合物研究進展[J].中國藥理學通報,2016,32(12):1643-7.
[7] Liu H Y, Jiang C T, Feng J, et al. Research progress of tanshinones[J].ChinPharmacolBull,2016,32(12):1643-7.
[8] Baby B, Antony P, Al Halabi W, et al. Structural insights into the polypharmacological activity of quercetin on serine/threonine kinases[J].DrugDesDevelTher, 2016,10:3109-23.
[9] 馬天翔, 史 寧, 陳 乾,等. 紅景天中8種成分體外抗氧化作用的比較[J]. 中國藥理學通報, 2012,28(9):1224-8.
[9] Ma T X, Shi N, Chen Q, et al. Comparison on the antioxidant activity of eight components from Rhodiolainvitro[J].ChinPharmacolBull, 2012,28(9):1224-8.
[10] de Freitas M M, Fontes P R, Souza P M, et al. Extracts of Morus nigra L. leaves standardized in chlorogenic acid, rutin and isoquercitrin: tyrosinase inhibition and cytotoxicity[J].PLoSOne, 2016,11(9):e0163130.
[11] Xie W, Wang M, Chen C, et al. Hepatoprotective effect of isoquercitrin against acetaminophen-induced liver injury[J].LifeSci, 2016,152:180-9.
[12] Wang Y, Nie H, Zhao X, et al. Bicyclol induces cell cycle arrest and autophagy in HepG2 human hepatocellular carcinoma cells through the PI3K/AKT and Ras/Raf/MEK/ERK pathways[J].BMCCancer, 2016,16(1):742.
[13] Deming D, Geiger P, Chen H, et al. ZM336372, a Raf-1 activator, causes suppression of proliferation in a human hepatocellular carcinoma cell line[J].JGastrointestSurg, 2008,12(5):852-7.
TheinhibitoryeffectofisoquercitrinonRaf/MEK/ERKsignalingpathwayinHepG2cells
JIANG Guo-jun1, LIU Tian-xu1, HUANG Gui-hong2, WU Ting1, TAO Li-qun2, ZHU Zhao-ming2
(1.LinguiClinicalCollege,GuilinMedicalUniversity,GuilinGuangxi541199,China;2.DeptofPharmacy,theSecondAffiliatedHospitalofGuilinMedicalUniversity,GuilinGuangxi541199,China)
AimTo study the inhibitory effect of isoquercitrin on Raf/MEK/ERK signaling pathway in HepG2 cells.MethodsMTT was used to detect the proliferation of human liver cancer HepG2 cells after the treatment of isoquercitrin. The morphology and growth of cells were observed under inverted microscope after the different concentrations of isoquercitrin(0, 40, 80, 160, 320 μmol·L-1) to treat HepG2 cells for 24 and 48 h. Cell cycle was assessed by flow cytometry. Ras, Raf, MEK, ERK expression was assayed by Western blot, and mRNA expression was detected by quantitative fluorescence PCR.ResultsIsoquercitrin could inhibit the growth of HepG2 cells in a concentration-and time-dependent manner. Typical morphological changes of apoptosis were observed by inverted microscopy after HepG2 cells were treated with different concentrations of of isoquercitrin for 24 h or 48 h. The cell cycle assay showed that with the increasing concentration of isoquerditrin, the number of cells that was arrested in G1phase gradually increased. Compared with the blank group, the expressions of Ras, Raf, MEK, ERK mRNA were down-regulated, and related proteins expression were also down-regulated(P<0.05), and these results had statistical significance.ConclusionIsoquercitrin can induce the apoptosis of HepG2 cells, which may be related to the Raf/MEK/ERK signaling pathway.
isoquercitrin; induce; HepG2 cells; Raf/MEK/ERK; signaling pathway; apoptosis
10.3969/j.issn.1001-1978.2017.10.011
A
:1001-1978(2017)10-1382-06
R284.1;R329.24;R329.25;R735.702.2
時間:2017-9-5 9:26 網絡出版地址:http://kns.cnki.net/kcms/detail/34.1086.R.20170905.0925.022.html
2017-05-29,
2017-07-03
國家自然科學基金資助項目(No 81260661);廣西自然科學基金資助項目(No 2012GXNSFAA053079);桂林市科學研究與技術開發計劃項目面上項目(No 20140120-1-9;No 20170109-47);廣西肝損傷與修復分子醫學重點實驗室子課題資助項目(No 15-140-46-14)
蔣國君(1990-),女,碩士生,研究方向:臨床藥學與應用,E-mail:784684807@qq.com; 黃桂紅(1972-),女,碩士,教授,碩士生導師,研究方向:糖尿病及哮喘藥物,通訊作者,E-mail:huangguihong120@126.com
猜你喜歡
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
天津醫科大學學報(2019年3期)2019-08-13 06:53:08
中成藥(2016年8期)2016-05-17 06:08:14
海峽科技與產業(2016年3期)2016-05-17 04:32:12
腫瘤預防與治療(2015年1期)2015-09-26 07:26:20
中國當代醫藥(2015年16期)2015-03-01 02:03:11