福建省登革熱監測點白紋伊蚊mtDNA-COI基因遺傳多態性研究
2017-11-21 03:19:24游麗斌何文祥翁育偉王金章闞乃鵬張擁軍
中國人獸共患病學報 2017年10期
游麗斌,朱 穎,何文祥,翁育偉,2,王金章,闞乃鵬,張擁軍,2
福建省登革熱監測點白紋伊蚊mtDNA-COI基因遺傳多態性研究
游麗斌1,朱 穎1,何文祥1,翁育偉1,2,王金章1,闞乃鵬1,張擁軍1,2
目的探討福建省登革熱監測點白紋伊蚊線粒體細胞色素C氧化酶亞基I(mtDNA-COI)基因序列特征和遺傳多態性。方法選擇福建省登革熱監測點2015—2016年送檢部分批次白紋伊蚊雌性成蚊,提取基因組DNA,擴增全長mtDNA-COI基因片段并測序,分析基因特征并構建系統發育樹。結果PCR擴增mtDNA-CO I基因共12個片段,測序得到的片段長度在1 476 bp—1 494 bp之間。與白紋伊蚊參比序列相比,同源性為99%,包含9個位點的堿基差異。根據不同單倍體型序列構建種系發育樹,發現6個登革熱監測點的白紋伊蚊分屬3個單倍體型,即H02、H03、H08。結論確定福建省登革熱監測點白紋伊蚊mtDNA-COI基因存在9個單核苷酸差異位點,包括H02、H03、H08共3種單倍體型,為福建省蟲媒病毒病監測及其防控提供技術支撐。
白紋伊蚊;線粒體細胞色素C氧化酶亞基I;遺傳多態性;種系發生分析
白紋伊蚊(Aedesalbopictus)屬雙翅目,蚊科,伊蚊屬(Aedes),是登革熱、黃熱病、西尼羅熱、基孔肯雅熱等病毒性疾病傳播的主要媒介[1]。白紋伊蚊原廣泛分布于亞洲的熱帶、亞熱帶地區和部分溫帶地區,過去數十年間其孳生地不斷擴大,北美、南美、歐洲和非洲大陸的一些國家均有分布[2]。在中國,白紋伊蚊孳生地遍及遼寧、陜西、江蘇、福建、云南等20個省份[3]。福建省自1873年廈門首次報道登革熱的暴發流行[4],至今已發生過多起登革熱的局部暴發流行,引起嚴重的公共衛生問題[5]。蚊媒控制不僅是處置這類暴發疫情的關鍵,也是預防其它輸入性蟲媒病毒病導致本地病例的核心。近年來國內外部分研究表明,生態環境變化、區域間交通運輸和人員交流,甚至蚊蟲防治措施的實施都會影響白紋伊蚊的種群遺傳結構[6-7]。不同地理種群白紋伊蚊在生態習性、媒介效能方面的差異影響其流行病學特征[8]。因此,研究白紋伊蚊不同環境下遺傳結構的差異和種群動態特征對于這些蟲媒病毒病的防控工作具有重要參考價值。
研究白紋伊蚊種群結構和系統發育分析的常用遺傳標記包括:同工酶、線粒體DNA(mtDNA)、核糖體DNA(rDNA)、微衛星等[9]。其中,最常見的靶基因是線粒體基因組細胞色素C氧化酶亞基I(mitochondrial cytochrome C oxidase subunit I,mtDNA-COI),該基因具有序列保守、進化速率恰當、無重組等特點,廣泛應用于白紋伊蚊蚊種鑒定和進化遺傳研究[10]。本研究首次嘗試擴增白紋伊蚊mtDNA-COI基因全長,分析2015—2016年福建省白紋伊蚊mtDNA-COI基因序列的多態性,為福建省蚊媒監測提供分子依據。
1 材料與方法
1.1樣本來源 選取福建省各登革熱監測點2015-2016年送檢的白紋伊蚊雌性成蚊,共計12批,分別來自6個監測點:福州臺江區(1)、連江縣(5)、廈門集美區(2)、寧德霞浦縣(1)、莆田涵江區(1)、建甌市(2),其中連江縣樣品采集時間為2015年6月、7月、8月、10月及2016年(表1)。
1.2基因組DNA的提取 采用高鹽抽提法提取基因組DNA,簡述如下:取數只白紋伊蚊腿于1.5 mL離心管中,加400 μL消化液(10 mmol/L NaCl,10 mmol/L Tris(pH8.0),10 mmol/L EDTA(pH8.0),0.5% SDS);加蛋白酶K(20 mg/mL)40 μL,55 ℃過夜;12 000 r/min離心2 min,轉移上清液到新的1.5 mL離心管;加400 μL醋酸鈉溶液(3 mol/L,pH5.2),-20 ℃放置15 min;12 000 r/min離心20 min,取上清液到新的1.5 mL離心管;加800 μL無水乙醇,-20 ℃放置15 min;12 000 r/min離心20 min,棄上清;加500 μL無水乙醇洗滌1次,12 000 r/min離心5 min,棄上清;加70%無水乙醇500 μL洗滌2次,12 000 r/min離心5 min,棄上清,自然干燥后加50 μL雙蒸滅菌水重懸,-20 ℃保存。
1.3PCR擴增和測序 根據白紋伊蚊線粒體基因組參比序列(GenBank:NC_006817)中mtDNA-COI基因編碼區(nt1436-2972)設計擴增全長編碼區的引物。上游引物(COX1FL-1424F):5′-CCAGCCATTTAATCGCGACA-3′和下游引物(COX1FL-2999R):5′-TTCATTGCACTAACTCGCCA-3′由生工生物工程(上海)股份有限公司合成。以提取的白紋伊蚊基因組DNA為模版,進行目的基因擴增。擴增試劑盒Premix TaqTM(Ex TaqTMVersion 2.0 plus dye)為大連TAKARA公司,反應體系為50 μL:Premix Taq 25 μL,上下游引物(10 μmol/L)各2 μL,模版2 μL,ddH2O 19 μL。PCR反應條件為:94 ℃預變性4 min;94 ℃變性30 s;50 ℃退火90 s;72 ℃延伸30 s;共30個循環;72 ℃后延伸10 min。擴增產物用1%瓊脂糖凝膠電泳鑒定,4 ℃保存,陽性產物送至鉑尚生物技術(上海)有限公司測序。
1.4mtDNA-CO I基因序列分析 應用Bioedit軟件查看雙向測序峰圖,核對序列,與參比序列進行多序列比對。從GenBank下載白紋伊蚊mtDNA-COI基因的72種單倍體型序列(GenBank:KC690896-KC690961,AB907796-AB907801)[11-12],埃及伊蚊mtDNA-COI基因序列(GenBank:AY056597)作為外群,應用Mega 6.0[13]構建系統發育樹。采用基于Kimura雙參數校正模型的最大似然法(maximum likelihood,ML),并用Bootstraping法對系統發育樹進行評估,Bootstrap步數設置為1 000。
2 結 果
2.1白紋伊蚊mtDNA-COI基因片段的PCR擴增 提取福建省6個登革熱監測點2015—2016年送檢的12批白紋伊蚊雌性成蚊腿的基因組DNA,擴增全長mtDNA-COI基因。PCR產物經1%瓊脂糖凝膠電泳,約在1 500 bp處可見單一條帶,與預期片段大小吻合(圖1)。
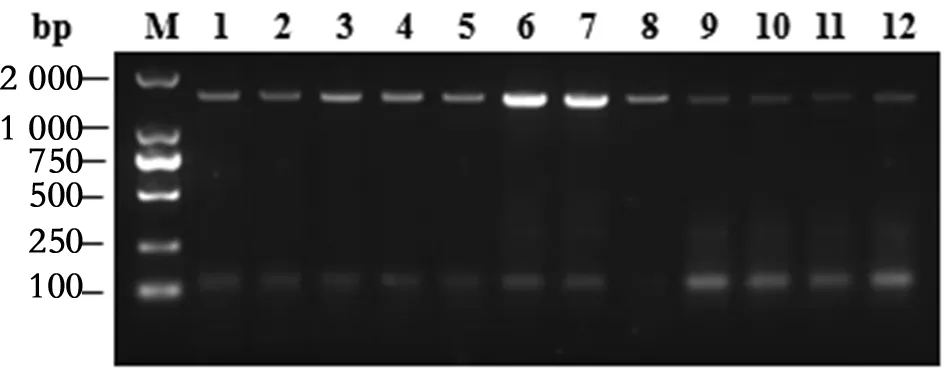
M:DL2000 marker;1-12號泳道樣品編號依次為:JO-2015, TJ-2015, JM-2015, HJ-2015, JO-2016, XP-2016, JM-2016, LJ-2016, LJ-2015-Jun, LJ-2015-Jul, LJ-2015-Aug, LJ-2015-OctM: DL2000 marker; Lane 1-12: A. albopictus samples from JO-2015, TJ-2015, JM-2015, HJ-2015, JO-2016, XP-2016, JM-2016, LJ-2016, LJ-2015-Jun, LJ-2015-Jul, LJ-2015-Aug, LJ-2015-Oct圖1 白紋伊蚊全長mtDNA-COI基因擴增Fig.1 PCR amplification of full-length mtDNA-COI genes from A. albopictus
2.2白紋伊蚊mtDNA-COI基因變異分析 分別采用擴增引物對PCR產物進行測序,經過拼接,片段長度在1 476 bp—1 494 bp之間(GenBank)。與白紋伊蚊參比序列(GenBank:NC_006817)進行多序列比對,結果顯示序列相似性均達99%,共確認9個變異位點(1505C/T、1580C/G、1678C/A、1706G/A、1804G/A、1966C/T、2104G/A、2296T/C、2805T/C)(表1)。來自福建省6個登革熱監測點的12個白紋伊蚊樣本之間,mtDNA-COI基因遺傳距離在0.00-0.002,觀察到僅在4個位點(1804G、2104G、2296T、2805T)存在一個或多個變異,其余5處位點均一致。
2.3系統發育分析 從GenBank下載已知白紋伊蚊mtDNA-COI基因的72種單倍體型序列(這些單倍體型分別來自中國、日本、新加坡、意大利、美國、哥斯達尼加、巴拿馬等),并以埃及伊蚊相應序列(GenBank:AY056597)作為外群,以共有片段1 388 bp的序列構建出系統發育樹(圖2)。從圖2中看出,來自福建省登革熱監測點的這12條序列分屬3個單倍體型。HJ-2015、JO-2015、JO-2016屬于H03單倍體型,LJ-2015-Aug屬于H02單倍體型,鄰近的單倍體型有H25、H62。其余8條序列JM-2015、JM-2016、LJ-2016、TJ-2015、XP-2016、LJ-2015-Jun、LJ-2015-Jul、LJ-2015-Oct屬于H08單倍體型,在進化樹的上下游為H12、H13等單倍體型。值得注意的是,連江縣的5個批次樣品,分屬H02和H08單倍體型,除了2015年8月樣品LJ-2015-Aug為H02單倍體型,其余樣品均為H08單倍體型。
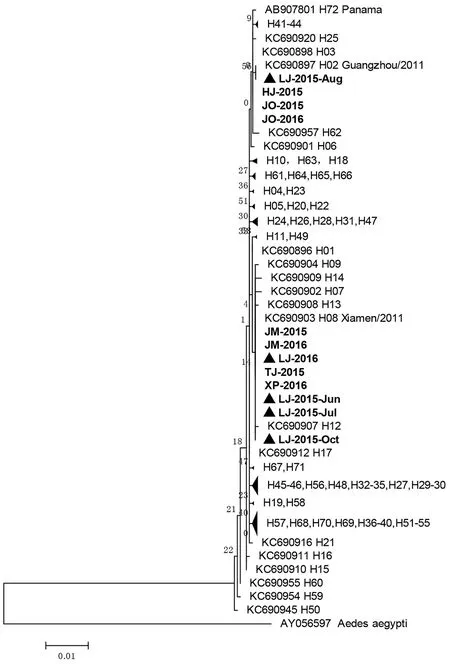
圖2 福建省登革熱監測點白紋伊蚊COI基因的系統發育樹Fig.2 Phylogenetic analysis on COI genes of A. albopictus from dengue surveillance sites in Fujian
3 討 論
國內外不同團隊先后開展了關于白紋伊蚊mtDNA-COI基因特征的研究,但分析的片段大都在400—700 bp之間[14-17],由于該基因本身相對保守,且蚊媒樣品來源受到地域限制,能夠提供的信息有限。近幾年,Zhong等[11]將研究對象的地理范圍擴大,首次對來自中國、日本、新加坡、意大利、美國等地白紋伊蚊樣品擴增了1 433 bp的片段,確定66種單倍體型(H01—H66),其中包括來自福建廈門的樣品。隨后日本學者Futami等[12]對哥斯達尼加、巴拿馬采集的白紋伊蚊進行了類似研究,擴增了1 390 bp的片段,確定了另外6種單倍體型(H67—H72)。因此,鑒于福建省開展登革熱蚊媒監測10余年,尚未明確我省存在哪些單倍體型的白紋伊蚊,本研究的首要目標,即是確定各登革熱監測點明確存在的白紋伊蚊單倍體型別。
首先,我們設計引物并擴增了白紋伊蚊的全長mtDNA-COI基因,選擇2015—2016年各登革熱監測點送檢的部分批次成蚊,擴增得到12條片段,測序后片段長度在1 476—1 494 bp之間。初步同源性分析顯示,所得12條mtDNA-COI片段與白紋伊蚊參考序列(GenBank:NC_006817)的同源性達99%。在發現的9個堿基差異位點中,核苷酸序列1505C、1580C、1678C、1706A、1966C位點上,6個登革熱監測點的12個白紋伊蚊樣本序列表現出一致性突變,而核苷酸序列1804G、2104G、2296T、2805T位點上,12個白紋伊蚊樣本序列表現出多態性(表1)。福建省6個監測點白紋伊蚊樣本間多序列比對結果顯示,白紋伊蚊樣本mtDNA-COI基因遺傳距離在0.00-0.002之間,遺傳距離極小,說明監測期間各監測點白紋伊蚊同源性較高。與Zhong等在廈門29只樣品中發現11個可變位點的結果接近,但低于深圳40只樣品發現24個可變位點的結果[11,18]。
表1 福建省登革熱監測點白紋伊蚊樣品信息及COI基因變異位點
Tab.1 List of A. albopictus samples collected from dengue surveillance sites in Fujian Province and variation sites of mtDNA-COI gene
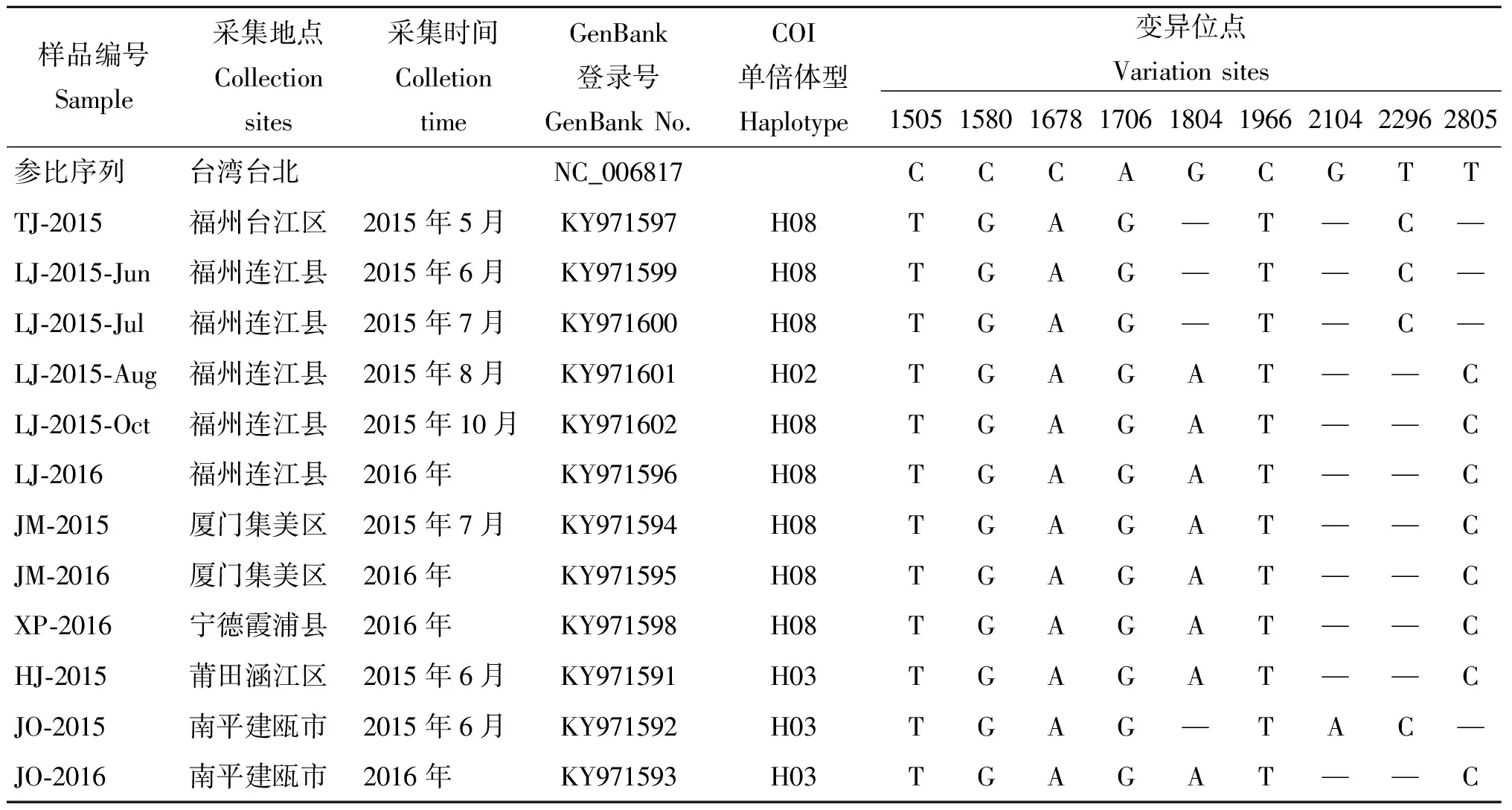
樣品編號Sample采集地點Collectionsites采集時間ColletiontimeGenBank登錄號GenBankNo.COI單倍體型Haplotype變異位點Variationsites150515801678170618041966210422962805參比序列臺灣臺北NC_006817CCCAGCGTTTJ?2015福州臺江區2015年5月KY971597H08TGAG—T—C—LJ?2015?Jun福州連江縣2015年6月KY971599H08TGAG—T—C—LJ?2015?Jul福州連江縣2015年7月KY971600H08TGAG—T—C—LJ?2015?Aug福州連江縣2015年8月KY971601H02TGAGAT——CLJ?2015?Oct福州連江縣2015年10月KY971602H08TGAGAT——CLJ?2016福州連江縣2016年KY971596H08TGAGAT——CJM?2015廈門集美區2015年7月KY971594H08TGAGAT——CJM?2016廈門集美區2016年KY971595H08TGAGAT——CXP?2016寧德霞浦縣2016年KY971598H08TGAGAT——CHJ?2015莆田涵江區2015年6月KY971591H03TGAGAT——CJO?2015南平建甌市2015年6月KY971592H03TGAG—TAC—JO?2016南平建甌市2016年KY971593H03TGAGAT——C
注:所有位點均參照GenBank登錄號NC_006817
Note: The variation sites are listed according to the sequence of NC_006817 in GenBank.
然后,通過與目前已知的72種白紋伊蚊mtDNA-COI單倍體型參比序列進行種系發生分析,本研究首次確定了2015-2016年福建省登革熱監測點的白紋伊蚊mtDNA-COI基因單倍體型(圖2)。共發現3種單倍體型,即H02、H03、H08。其多樣性低于文獻報道中廣州6種、廈門11種單倍體型,而與江蘇2種單倍體型接近[11]。對照Zhong等觀察到的結果,其中H08單倍體型為廈門獨有(29只廈門樣品中有9只屬于H08型),本研究中12批次樣品也有8份為H08型。因此,H08單倍體型是否為福建省特有,需要今后更多序列數據證明。同時,本研究中有3份樣品屬于H03單倍體型。該型別在Zhong等觀察到的結果屬于優勢型,總共346份樣品中有113份屬于H03,并且在廣州、廈門、江蘇樣品中占優勢(20/32、9/29、23/30)。從以上數據看來,H03單倍體型極有可能是國內的優勢基因型。另外,本研究還發現唯一一份樣品LJ-2015-Aug屬于H02型,而該單倍體型根據文獻僅出現于來自廣州的樣品,如果能夠得到其它證據的支持,表明福建省可能發生過不同遺傳背景的白紋伊蚊入侵事件。從這個角度來看,鑒于目前白紋伊蚊孳生地在全球不斷擴張的事實,國內外研究人員按照統一的標準和單倍體型分型方案,將有助于精確定位白紋伊蚊的來源、遷徙路線及種群進化,科學評估新的種群入侵可能帶來的危害。
綜上所述,本研究首次擴增比較來自福建省各登革熱監測點白紋伊蚊mtDNA-COI全長片段序列,觀察到各監測點之間樣品同源性較高,并且參照國內外現有的72種mtDNA-COI單倍體型確定了各監測點的基因型,為今后在分子水平持續進行蚊媒監測、推動蟲媒病毒病防控奠定了堅實的基礎。
[1] Haddad N, Mousson L, Vazeille M, et al.Aedesalbopictusin Lebanon, a potential risk of arboviruses outbreak[J]. BMC Infect Dis, 2012, 12:300. DOI: 10.1186/1471-2334-12-300
[2] Qian QQ, Fang YL, Zhang JQ, et al. Research on the different geographic strains ofAedesalbopictus[J]. Chin Frontier Health Quarant, 2016, 39(1): 66-68. DOI: 10.16408/j.1004-9770.2016.01.018 (in Chinese)
錢茜茜, 方義亮, 張建慶,等. 不同地理株白紋伊蚊研究現狀[J]. 中國國境衛生檢疫雜志, 2016, 39(1):66-68.
[3] Lu BL. Fauna Sinica, Diptera: Culicidae (1, 2)[M]. Beijing: Science Press. 1997: 244. (in Chinese)
陸寶麟. 中國動物志.Ⅷ.雙翅目: 蚊科(上、下)[M].北京: 科學出版社,1997: 244
[4] Chen YD. Epidemics of dengue fever in global prevalence area[J]. Chin J Epidemiol, 1990, 11(1): 51-54. (in Chinese)
陳韻冬. 登革熱在全球主要流行區的流行近況[J]. 中華流行病學雜志, 1990, 11(1):51-54.
[5] Hu TS, Zhang HL, Liu YH, et al. Molecular characteristics of the full-length genome of dengue serotype 1 virus strains isolated from dengue fever cases in Sino-Myanmar border region in Yunnan Province, China[J]. Chin J Zoonoses, 2017, 33(6):473-480. DOI:10.3969/j.issn.1002-2694.2017.06.001(in Chinese)
胡挺松, 張海林, 劉永華等. 云南中緬邊境登革1型病毒全基因組序列特征研究[J]. 中國人獸共患病學報, 2017, 33(6):473-480.
[6] Lin Y, Zhang SY, Lin YY, et al. Genetic polymorphism analysis on rDNA ITS2 from different geographical strains ofAedesalbopictusin Fujian Province[J]. Chin J Zoonoses, 2010,(8): 735-738. (in Chinese)
林巖, 張山鷹, 林耀瑩,等. 福建省不同地理株白紋伊蚊rDNA ITS2基因多態性分析[J]. 中國人獸共患病學報, 2010,(8):735-738.
[7] Huber K, Loan LL, Hoang TH, et al. Temporal genetic variation inAedesaegyptipopulations in Ho Chi Minh City(Vietnam)[J]. Heredity(Edinb), 2002, 89(1): 7-14.
[8] Tien TK, Vazeille-Falcoz M, Mousson L, et al.Aedesaegyptiin Ho Chi Minh City (Viet Nam): susceptibility to dengue 2 virus and genetic differentiation[J]. Trans Roy Soy Trop Med Hyg, 1999, 93(6): 581-586.
[9] Goubert C, Minard G, Vieira C, et al. Population genetics of the Asian tiger mosquitoAedesalbopictus, an invasive vector of human diseases[J]. Heredity, 2016, 117(3): 125-134. DOI: 10.1038/hdy.2016.35
[10] Folmer O, Black M, Hoeh W, et al. DNA primers for amplification of mitochondrial cytochrome C oxidase subunitIfrom diverse metazoan invertebrates[J]. Mol Mar Biol Biotechnol, 1994, 3(5): 294-299.
[11] Zhong D, Lo E, Hu R, et al. Genetic analysis of invasiveAedesalbopictuspopulations in Los Angeles County, California and its potential public health impact[J]. PLoS One, 2013, 8(7): e68586. DOI:10.1371/journal.pone.0068586
[12] Futami K, Valderrama A, Baldi M, et al. New and common haplotypes shape genetic diversity in Asian tiger mosquito populations from Costa Rica and Panama[J]. J Econ Entomol, 2015, 108(2):761-768. DOI: 10.1093/jee/tou028
[13] Tamura K, Stecher G, Peterson D, et al. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0[J]. Mol Biol Evol, 2013, 30(12): 2725-2729. DOI: 10.1093/molbev/mst197
[14] Fang YL, Zhang SY, Xie HG, et al. Study on mtDNA-COI genetic characteristics ofAedesalbopictusin different geographic strains[J].Strait J Prev Med, 2011, 17(4): 1-4. (in Chinese)
方義亮, 張山鷹, 謝漢國,等. 不同地理株白紋伊蚊mtDNA-COI基因特征研究[J]. 海峽預防醫學雜志, 2011, 17(4): 1-4.
[15] Ismail NA, Dom NC, Ismail R, et al. Mitochondrial cytochrome oxidase I gene sequence analysis ofAedesalbopictusin Malaysia[J]. J Am Mosq Control Assoc, 2015, 31: 305-312. DOI: 10.2987/moco-31-04-305-312.1
[16] Zhang RL, Yao GQ, Pan XQ, et al. Genetic diversities of different geographical populations ofAedesalbopictusbased on mitochondrial gene COI[J]. Chin J Zoonoses, 2017, 33(4):316-320.DOI:10.3969/j.issn.1002-2694.2017.04.005 (in Chinese)
張瑞玲, 姚廣琴, 潘曉倩,等. 不同地理種群白紋伊蚊線粒體基因COI的遺傳多樣性分析[J]. 中國人獸共患病學報, 2017, 33(4):316-320.
[17] Guo S, Ling F, Wang JN, et al. Genetic polymorphism analysis of cytochrome C oxidase subunit I gene inAedesalbopictusfrom Zhejiang Province, China[J]. Chin J Zoonoses, 2016, 32(2):133-136. DOI:10.3969/j.issn.1002-2694.2016.02.007(in Chinese)
郭頌, 凌鋒, 王金娜,等. 浙江省不同地理株白紋伊蚊mtDNA-COI基因多態性研究[J]. 中國人獸共患病學報, 2016, 32(2):133-136.
[18] Yang F, Zhang RL, Liu Y, et al. Genetic polymorphism analysis on mtDNA COⅠ ofAedesalbopictusin Shenzhen, China[J]. Chin J Vector Biol Control, 2015, 26(4): 337-340. DOI: 10.11853/j.issn.1003.4692.2015.04.002 (in Chinese)
陽帆, 張仁利, 劉陽,等. 深圳市白紋伊蚊細胞色素C氧化酶亞基Ⅰ基因多態性分析[J]. 中國媒介生物學及控制雜志, 2015, 26(4): 337-340.
GeneticdiversityofcytochromeCoxidasesubunitIgenesinAedesalbopictusfromdenguesurveillancesitesinFujianProvince,China
YOU Li-bin1, ZHU Ying1, HE Wen-xiang1, WENG Yu-wei1,2, WANG Jin-zhang1, KAN Nai-peng1, ZHANG Yong-jun1,2
(1.FujianCenterforDiseaseControlandPrevention,Fuzhou350001,China;2.FujianProvincialKeyLaboratoryofZoonosisDiseaseResearch,Fuzhou350001,China)
The mosquitoAedesalbopictusis the primary vector for dengue virus transmission in Fujian Province. Despite that active dengue surveillance has been launched in several sites since 2005, the genetic diversity ofA.albopictusfrom these sites remains exclusive. In this study, mosquito pools collected from dengue surveillance sites during 2015-2016 were randomly selected, the full-length mitochondrial cytochrome C oxidase subunit I (mtDNA-COI) were amplified and sequenced. Preliminary sequence alignment of 12 amplicons with the reference sequence indicated 99% homology at nucleotide level, due to variations at 9 nucleotide sites. Three haplotypes, namely H02, H03 and H08, were determined based on phylogenetic analysis with 72 reference sequences of known haplotypes. These observations facilitate surveillance and control of arboviral diseases in Fujian.
Aedesalbopitus; mitochondrial cytochrome C oxidase subunit I; genetic diversity; phylogenetic analysis
Zhang Yong-jun,Emial:zhangyj@fjcdc.com.cn
10.3969/j.issn.1002-2694.2017.10.004
福建省科技廳引導性項目(No. 2016Y0011)和福建省醫學創新課題(No. 2015-CXB-13)聯合資助
張擁軍,Email:zhangyj@fjcdc.com.cn
1.福建省疾病預防控制中心,福州 350001;
2.福建省人獸共患病重點實驗室,福州 350001
373
A
1002-2694(2017)10-0872-05
Supported by the Department of Science and Technology, Fujian Province (No. 2016Y0011) and the Innovative Project of Medical Science of Fujian Province (No. 2015-CXB-13)
2017-01-12編輯李友松
