家養雙峰駝SRY基因的序列分析與拷貝數研究
2017-11-29 03:34:34陳慧玲任戰軍葉翔楊趙紀萍張成東
西北農業學報 2017年11期
陳慧玲,任戰軍,葉翔楊,趙紀萍,張成東
(西北農林科技大學 動物科技學院,陜西楊陵 712100)
家養雙峰駝SRY基因的序列分析與拷貝數研究
陳慧玲,任戰軍,葉翔楊,趙紀萍,張成東
(西北農林科技大學 動物科技學院,陜西楊陵 712100)
為探究中國家養雙峰駝SRY基因的序列以及群體間拷貝數變異,對7個中國家養雙峰駝群體共94個個體的SRY基因部分序列進行測序,分析SRY基因序列與其他物種的相似性;并且以 CYP2A基因為內參基因,運用實時熒光定量PCR技術檢測SRY基因的拷貝數。結果表明,中國家養雙峰駝與人、馬、豬、羊及普通牛的SRY基因相似度分別為67.7%~68.1%、68.5%~69.4%、73.8%~74.4%、73.8%~74.8%、74.1%~74.8%。在家養雙峰駝SRY基因的多個拷貝中,除1個拷貝外,其余拷貝的HMG序列與羊駝、人、馬、豬、綿羊及普通牛的HMG序列相比,相似度分別為97.3%、84.2%、85.6%、86.3%、87.0%、85.6%。SRY基因在中國家養雙峰駝Y染色體上以多拷貝形式存在,變幅為 1~9 個拷貝。表明中國家養雙峰駝SRY基因的HMG序列是高度保守的,而且該基因屬于多拷貝基因。
家養雙峰駝;SRY基因;拷貝數
性別決定基因(Sex-determining-region on Y chromosome,SRY)存在于哺乳動物Y染色體的雄性特異區,具有性別決定功能,參與睪丸組織的發育過程。該基因編碼的蛋白包含C端、N端和高泳動類非組蛋白(High-mobility group, HMG)區域,C端與N端在物種間保守性較弱,而HMG區域在物種間具有高度保守性[1-2]。早期對SRY基因的研究發現,SRY保守區的突變會破壞其雄性性別決定功能,引起性別反轉[3-4],SRY基因也能夠誘導基因型為XX的雌性小鼠向雄性發育[5]。SRY蛋白能夠識別并結合 SOX9(Sry-related HMG box-9)基因上游的增強子[6],使睪丸支持細胞的前體細胞中 SOX9基因的轉錄水平顯著上調,進而促進下游基因的轉錄,誘導睪丸支持細胞的分化。隨后,分化后的睪丸支持細胞組裝成睪丸索,并且刺激生殖細胞、睪丸間質細胞、睪丸血管細胞以及其他間質細胞的分化,最終形成睪丸組織。
在多數哺乳動物中,SRY基因是以單拷貝的形式存在[7],但是少數物種中該基因存在多拷貝現象。貓、小鼠、家兔和水牛中SRY為多拷貝基因[8-12]。接觸輻射或性染色體異常的人類SRY基因會出現拷貝數變異現象[13]。迄今為止,未見中國家養雙峰駝SRY基因研究的報道。本試驗以中國4個地區7個家養雙峰駝群體為研究對象,對其SRY基因部分序列進行測序,并且運用實時熒光定量PCR技術檢測其拷貝數,以期對家養雙峰駝SRY基因有初步了解。
1 材料與方法
1.1 試驗材料
按照隨機整群抽樣的方法,采集內蒙古(阿拉善、蘇尼特)、甘肅(河西)、青海(海西州)、新疆(南疆、北疆和東疆)4個地區94峰雄性家養雙峰駝的血樣(表1)。另外,采集5峰雌性家養雙峰駝血樣作為陰性對照。頸靜脈采集血液于已滅菌的離心管中,加入ACD抗凝劑后在-80 ℃保存,備用。
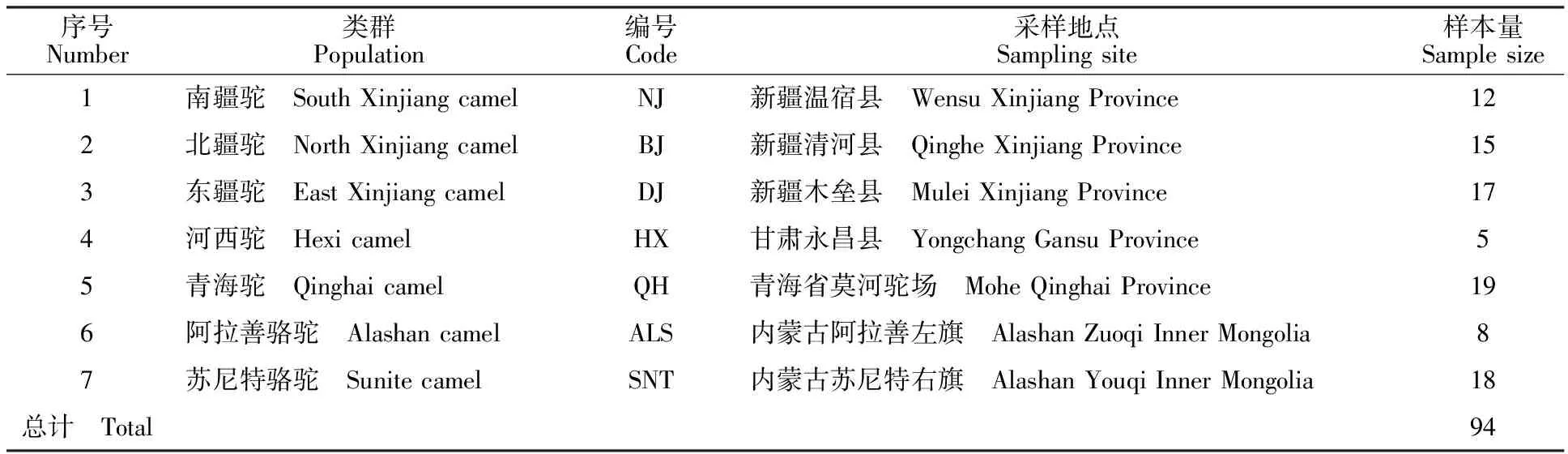
表1 家養雙峰駝樣本來源與樣本量Table 1 Source and number of domestic bactrian camel
1.2 SRY引物設計與合成
依據GenBank數據庫中雄性野生雙峰駝的相應序列(GenBank登錄號:NW_006220067),運用Primer 5.0軟件設計預期擴增片段長度為708 bp 的引物用于SRY基因的測序。其中上游引物:5′-ATGCTTCTGCTATGTTTGCG-3′,下游引物:5′-ACCAAAAGTAACGGTGAGAATG-3′。引物由上海生工生物工程股份有限公司合成。
1.3 PCR擴增與測序
利用酚—氯仿法提取基因組DNA[14],采用10 g/L瓊脂糖凝膠電泳和Nanodrop 2000檢測其純度和質量濃度。將提取效果良好的基因組DNA樣品統一稀釋至20 ng/μL,置于-20 ℃保存,備用。
挑選6~10個雄性個體的基因組DNA混合成DNA池,共構建13個DNA池,-4 ℃保存。DNA池中每個樣品的質量濃度盡可能保持一致。
首先用SRY引物對雄性DNA池進行PCR擴增(以雌性雙峰駝DNA為陰性對照),采用10 g/L瓊脂糖凝膠電泳檢測引物的雄性特異性,并且確定其最適退火溫度為65.3 ℃。反應體系為10 μL:EsTaqMaster Mix 5 μL,上下游引物各0.2 μL,DNA模板0.5 μL,ddH2O 4.1 μL(北京康為世紀有限公司)。 PCR反應程序:95 ℃變性5 min;94 ℃ 30 s,65 ℃ 40 s, 72 ℃ 50 s,34個循環;72 ℃ 10 min。雄性特異的擴增產物由上海生工生物工程股份有限公司進行測序。
如果測序結果出現雙峰,則對該DNA池中的每個個體分別進行PCR擴增測序,以再次確定SNP位點。如果單個個體的序列峰圖仍在相同位點處出現雙峰,則需要進行TA克隆測序予以驗證。在每個群體中分別挑選1個個體進行PCR擴增,用瓊脂糖凝膠DNA回收試劑盒(Takara公司)純化回收PCR產物,產物經過純化與回收之后,與pGEM-T載體連接,然后轉化至大腸桿菌感受態細胞,在37 ℃平板培養。每個個體隨機挑選10個陽性菌落在具有Amp+的LB培養基中震蕩培養過夜,吸取部分菌液送至上海生工生物工程股份有限公司進行克隆測序。
1.4 SRY序列相似性分析
用DNASTAR軟件對所有個體的序列進行比對,分析個體間序列的差異。運用MegAlign軟件分析人(GenBank登錄號:JQ811934.1)、馬(GenBank登錄號:NM_001081810.1)、豬(GenBank登錄號:U49860.3)、綿羊(GenBank登錄號:Z30265.1)及普通牛(GenBank登錄號:Z30327.1)的SRY基因序列與測序所得的SRY基因部分序列的相似度以及物種間HMG區DNA序列的相似度。
1.5 熒光定量引物的設計與合成
TA克隆測序完成后,運用MegAlign軟件比對所有序列,選擇在SRY基因的保守區設計SRYa引物用于實時熒光定量PCR擴增。選擇常染色體上的單拷貝基因細胞色素P4502A( CYP2A)為內參基因[14],計算SRY基因的拷貝數。引物信息見表2。
1.6 熒光定量PCR擴增
采用實時熒光定量PCR技術檢測基因的拷貝數。首先挑選質量較好的DNA模板用于標準曲線的制作,質量濃度梯度依次為40、20、10、5和2.5 ng/μL,每個質量濃度設置3個重復。熒光定量PCR程序完成之后,獲得各質量濃度相對應的Ct值,分別制作CYP2A和SRYa的標準曲線。將待測樣本均稀釋至5 ng/μL,每個樣本在同一板中同時進行2對引物的擴增,而且每對引物對應的每個DNA樣本均設置3個重復,每板都設有相同的校正樣本以減小板間的系統誤差。

表2 SRYa和CYP2A引物信息Table 2 Information of SRYa and CYP2A
反應體系:DNA模板0.8 μL,SYBR Premix EXTaqⅡ(2×)(Takara) 5 μL,上下游引物(10 μmol/L)各0.4 μL,RNase-Free Water 3.4 μL。熒光定量反應程序為:95 ℃ 預變性30 s;95 ℃變性5 s,退火30 s,35個循環;95 ℃ 10 s,65 ℃ 5 s,65 ℃~95 ℃(0.5 ℃/s)。
1.7 基因拷貝數的計算
參照HAMILTON等[15]的方法計算SRY基因的拷貝數。計算公式如下:
依據獲取的各質量濃度梯度所對應的Ct值,繪制標準曲線,可以獲得標準曲線的斜率k,進而計算出引物的擴增效率:E=10-1/k。
依據所有96孔板中校正樣本的Ct值,計算校正樣本Ct值的平均值。再根據平均值計算校正樣本的拷貝數:CN校正樣本=(Er)Ct r/(Et)Ct t。
每個96孔板中校正樣本都設置3個重復,分別計算不同板中校正樣本3個Ct值的平均值,再將不同板的待測樣本校正到同一水平,待測樣本與校正樣本之間校正系數的計算:
R=(Et)△Ct t/(Er)△Ct r。
待測樣本拷貝數計算:CN=(CN校正樣本) ×R。
k代表標準曲線的斜率,t代表目的基因SRY,r代表內參基因 CYP2A,E代表擴增效應,Et代表SRYa引物的擴增效率,Er代表CYP2A 引物的擴增效率,Ctr代表CYP2A引物的Ct值,Ctt代表SRYa引物的Ct值,△Ctt=校正樣本SRYa的Ct值-待測樣本SRYa 的Ct值,△Ctr=校正樣本CYP2A 的Ct值-待測樣本CYP2A 的Ct值。
運用SPSS 18.0軟件Kolmogorov-Smirnov標準對所研究個體的基因拷貝數進行正態分布檢驗。
2 結果與分析
2.1 SRY基因的PCR擴增
Touchdown PCR擴增后,擴增產物經10 g/L的瓊脂糖凝膠電泳檢測,結果如圖1所示,陰性對照與空白對照無擴增產物,而所有雄性個體的條帶明亮清晰,片段長度與預期長度一致,并且無非特異性擴增,表明SRY引物具有雄性特異性。

圖1 SRY引物的雄性特異性檢測Fig.1 Detection of SRY specificity in male camel
2.2 SRY基因序列分析
運用DNASTAR軟件對SRY引物擴增所得序列進行比對,結果如圖2所示,13個DNA池中SRY基因擴增片段的測序峰圖均出現雙峰,本圖只展示其中3個DNA池的比對結果,其余DNA池測序結果與該圖一致,因此初步推斷SRY基因存在SNP位點以及堿基的插入。
以DNA池中各單樣為模板分別對SRY引物進行PCR擴增并對擴增產物測序,經過序列比對,對應DNA池中SNP位點處尋找各個單樣之間堿基的差異。結果如圖2所示,每個個體的測序峰圖在與池DNA序列的相同位點處存在雙峰,而且也具有堿基插入位點,其他個體測序結果也與該結果一致,由此推測在家養雙峰駝中SRY基因可能是多拷貝基因。
TA克隆測序結果如圖3所示,同一個體10個克隆之間的序列存在變異位點和1個AT二堿基插入,本圖只展示3個克隆序列的比對結果,其他群體的測序結果與青海駝一致。由于Y染色體MSY區不與X染色體重組,因而證明在家養雙峰駝中單個個體出現SNP位點的SRY基因是多拷貝基因。

圖2 3個DNA池的SRY基因測序Fig.2 Sequencing result of the SRY gene of three gene pools

圖3 1峰青海駝的SRY基因克隆測序Fig.3 SRY gene of a Qinghai sample by PCR cloning and sequencing
2.3 不同物種間序列相似性分析
對中國家養雙峰駝的SRY基因部分序列進行克隆測序,共獲得708 bp的序列。運用MegAlign軟件對家養雙峰駝SRY基因不同拷貝的序列與人、馬、豬及普通牛的SRY基因序列進行比對分析,結果發現,中國家養雙峰駝SRY基因與人、馬、豬、綿羊及普通牛的SRY基因相似度約為67.7%~68.1%、68.5%~69.4%、73.8%~74.4%、73.8%~74.8%、74.1%~74.8%。
不同物種的SRY蛋白都含有保守的HMG結構域,該結構域是SRY蛋白重要的功能區。因此本試驗比對羊駝HMG序列和家養雙峰駝708 bp 的序列,結果顯示在家養雙峰駝SRY基因的多個拷貝中,除1個拷貝外,其余拷貝中有146 bp的序列與羊駝HMG序列相似度高達97.3%,與人、馬、豬、綿羊及普通牛的HMG序列相比,相似度分別為84.2%、85.6%、86.3%、87.0%、85.6%。因此,可以初步確定中國家養雙峰駝SRY基因的HMG區域。由于拷貝之間的HMG序列有5個突變位點,因而認為家養雙峰駝的HMG序列為:
TCATTGTATGGGCTCGTGATCAAAGGCGAAAGGTGGCTCTASAGAATMCCAAA- ATGCAGAACKCAGAGATCAGCAAGCGGC-TGGGATACCAGTGGAAATTACTTACAGA-AGSTGAAAAGCGGCCGTTCTTCGAGGAG-GCRCAGAGA。
結合野生雙峰駝SRY基因的預測序列、氨基酸序列(GenBank登錄號:XP_006195462)以及羊駝HMG的氨基酸序列(GenBank登錄號:ABI31754.1),推測家養雙峰駝HMG的氨基酸序列為:
IVWARDQRRKVAL(E/Q)N(T/P)KMQN(A/S)EISKRLGYQWKLLTE(A/G)EKRPFFEE AQR。其與野生雙峰駝和羊駝序列相似度約為95.8%。
2.4 SRYa和CYP2A引物的PCR擴增
Touchdown PCR擴增后,用10 g/L的瓊脂糖凝膠電泳檢測(圖4),結果顯示SRYa引物只在雄性個體中有目的條帶,雌性個體無條帶。CYP2A引物的擴增產物為單一條帶,且無引物二聚體。

圖4 SRYa(a)和CYP2A(b)引物的特異性檢測Fig.4 Detection of SRYa(a) and CYP2A(b) specificity
2.5 溶解曲線
引物的熒光定量溶解曲線如圖5所示,2對引物的擴增產物單一,無引物二聚體及非特異性條帶擴增。由此可見,該引物均可用于后續的熒光定量PCR擴增。
2.6 標準曲線
CYP2A和SRYa引物的標準曲線如圖6所示,CYP2A和SRYa引物的標準曲線斜率分別為-3.207 1和-3.223 3。由此計算出CYP2A和SRYa引物的擴增效率分別為2.05和2.04,R2均大于0.99,表明2對引物的擴增效率較好,可用于后續的試驗。
2.7 拷貝數計算
根據公式計算出校正樣本SRY基因的拷貝數是2,隨后計算94個樣本的拷貝數。結果顯示,在94個個體中,SRY基因拷貝數中值數為4,變幅為1~9。各群體拷貝數見表3。

圖5 CYP2A和SRYa的 qPCR 溶解曲線Fig.5 Dissociation curves of CYP2A and SRYa
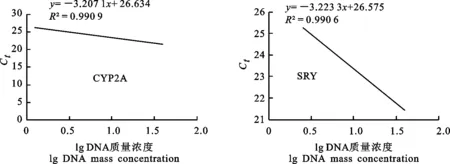
圖6 CYP2A 和SRY擴增引物的標準曲線Fig.6 Standard curves of CYP2A and SRY primers

表3 7個家養雙峰駝群體SRY基因的拷貝數中值Table 3 Copy median number (CN) of SRY in seven populations
94個個體基因拷貝數的正態分布分析結果顯示所有個體的拷貝數均不服從正態分布(圖7-a)。箱線圖結果發現少數雙峰駝個體的SRY基因拷貝數偏高或偏低,大約占群體的1.06%。其中,蘇尼特駱駝有1個個體與其他個體離散,拷貝數為9(圖7-b)。
3 討 論

平均值=4.11,標準差 = 1.44 Mean = 4.11,standard deviation = 1.44
SRY基因是哺乳動物性別決定的開關基因,它的表達能夠啟動雄性發育的級聯信號通路,誘導性別特異的性腺發育,因而在物種的性別分化過程中具有至關重要的作用。本試驗通過對中國家養雙峰駝的SRY基因部分序列進行測序,而后將所得序列與其他物種SRY基因序列進行比對,結果顯示其與牛和豬的相似度很高。Jirimutu等[14]通過對動物基因組中的 2 345 個單拷貝直系同源構建系統進化樹,結果發現駱駝與牛和豬的關系最近。雖然本試驗所研究的SRY基因是多拷貝基因,但是物種間的序列相似度也能在一定程度上反映駱駝與牛、豬的進化關系,進化關系越近,序列變異程度越小。因此,該結果與Jirimutu等[14]的研究結果一致。
本試驗運用實時熒光定量PCR技術檢測中國家養雙峰駝SRY基因的拷貝數,結果發現拷貝數變幅為1~9。和其他物種相比,拷貝數也有差異。目前研究發現,SRY基因在人類、馬、牛、黑猩猩、豬中均為單拷貝基因[7,15-17],而在家貓和水牛中則為多拷貝基因[10,12]。大鼠的SRY基因有11個拷貝,拷貝序列之間有一定的差異[11],11個拷貝中至少有6個拷貝能夠正常表達[18]。不同物種拷貝數的差異進一步表明Y染色體在不同的物種間表現出異質性。
HMG區域在物種間相對保守,是SRY基因發揮性別決定功能的區域。因此,本試驗對中國家養雙峰駝SRY基因的HMG區域進行預測,結果顯示,HMG的堿基序列位于本試驗所擴增片段的193~338 bp,而二堿基插入在該區域的下游,因而不會引起該區域序列的移碼突變。由于不同拷貝的序列之間有變異位點存在,部分氨基酸會發生改變,氨基酸的改變是否會影響SRY基因的功能還是未知數。目前,對于大鼠SRY基因序列的分析結果顯示,11個拷貝的SRY序列之間的突變位點導致氨基酸序列改變,進而引起核定位以及轉錄活性改變[11]。前人[19-20]研究發現SRY蛋白的功能并不局限于性別決定,其在成年雄性大腦、腎臟和腎上腺中也有表達。本試驗所得708 bp的序列在拷貝序列間也存在多個突變位點及1個二堿基的插入位點,推測這些位點的改變可能會影響睪丸組織的發育。這些變異也可能與SRY基因的其他功能相關聯,然而這些問題還有待于進一步探討。
Reference:
[1] LUNDRIGAN B L,TUCKER P K.Tracing paternal ancestry in mice,using the Y-linked,sex-determining locus,Sry[J].MolecularBiologyandEvolution,1994,11(3):483-492.
[2] NACHMAN M W,AQUADRO C F.Polymorphism and divergence at the 5’ flanking region of the sex-determining locus,Sry,in mice[J].MolecularBiologyandEvolution,1994,11(3):539-547.
[3] HARLEY V R,JACKSON D T,HEXTALL P J,etal.DNA binding activity of recombinant SRY from normal males and XY females[J].Science,1992,255(5043):453-456.
[4] HAWKINS J R,TAILOR A,BERTA P,etal.Mutational analysis of the SRY:nonsense and missense mutations in XY sex reversal[J].HumanGenetics,1992,88(4):471-474.
[5] KOOPMA P,GUBBAY J,VIVIAN N,etal.Male development of chromosomally female mice transgenic for SRY[J].Nature,1991,351(6322):117-121.
[6] SEKIDO R,LOVELL-BADGE R.Sex determination involves synergistic action of SRY and SF1 on a specific SOX9 enhancer[J].Nature,2008,453(7197):930-934.
[7] PARIA N,RAUDSEPP T,WILKERSON A J P,etal.A gene catalogue of the euchromatic male-specific region of the horse Y chromosome:comparison with human and other mammals[J].PLoSOne,2011,6(7):e21374.
[8] TOURE A,SZOT M,MAHADEVAIAH S K,etal.A new deletion of the mouse Y chromosome long arm associated with the loss of Ssty expression,abnormal sperm development and sterility[J].Genetics,2004,166 (2):901-912.
[9] GERALDES A,FERRAND N.A 7-bp insertion in the 3′ untranslated region suggests the duplication and concerted evolution of the rabbitSRYgene[J].GeneticsSelectionEvolution,2006,38:313-320.
[10] WILKERSON A J P,RAUDSEPP T,GRAVES T,etal.Gene discovery and comparative analysis of X-degenerate genes from the domestic cat Y chromosome[J].Genomics,2008,92(5):329-338.
[11] PROKOP J W,UNDERWOOD A C,TURNER M E,etal.Analysis of SRY duplications on the Rattus norvegicus Y-chromosome[J].BMCGenomics,2013,14(1):117-124.
[12] 徐舒遠.水牛Y-SNPs篩選及多拷貝基因鑒定[D].陜西楊陵:西北農林科技大學,2014.
XU SH Y.Y-SNPs scanning and multi-copy genes identification in water buffalo[D].Yangling Shaanxi:Northwest Aamp;F University,2014 (in Chinese with English abstract).
[13] PREMI S,SRIVASTAVA J,CHANDY S P,etal.Tandem duplication and copy number polymorphism of theSRYgene in patients with sex chromosome anomalies and males exposed to natural background radiation[J].MolecularHumanReproduction,2006,12(2):113-121.
[14] JIRIMUTU,WANG Z,DING G,etal.Genome sequences of wild and domestic bactrian camels[J].NatureCommunications,2012,3:1202-1202.
[15] HAMILTONA C K,FAVETTA L A,DI MEO G P,etal.Copy number variation of testis-specific protein,Y-encoded (TSPY) in 14 different breeds of cattle (Bostaurus)[J].SexualDevelopment,2009,3(4):205-213.
[16] SKALETSKY H,KURODA-KAWAGUCHI T,MINX P J,etal.The male-specific region of the human Y chromosome is a mosaic of discrete sequence classes[J].Nature,2003,423(6942):825-837.
[17] CHANG T C,YANG Y,RETZEL E F,etal.Male-specific region of the bovine Y chromosome is gene rich with a high transcriptomic activity in testis development[J].ProceedingsoftheNationalAcademyofSciences,2013,110(30):12373-12378.
[18] TURNER M E,MARTIN C,MARTINS A S,etal.Genomic and expression analysis of multipleSRYloci from a single Rattus norvegicus Y chromosome[J].BMCGenetics,2007,8(1):1.
[19] CZECH D P,LEE J,SIM H,etal.The human testis-determining factor SRY localizes in midbrain dopamine neurons and regulates multiple components of catecholamine synthesis and metabolism[J].JournalofNeurochemistry,2012,122(2):260-271.
[20] DEWINGE P,CHIANG C W K,SINCHAK K,etal.Direct regulation of adult brain function by the male-specific factor SRY[J].CurrentBiology,2006,16(4):415-420.
CorrespondingauthorREN Zhanjun, male, associate professor. Research area:economic zoology research.E-mail:renzhanjun@nwsuaf.edu.cn
(責任編輯:顧玉蘭Responsibleeditor:GUYulan)
CopyNumberandSequenceAnalysisofSRYGeneinChineseDomesticBactrianCamel
CHEN Huiling, REN Zhanjun, YE Xiangyang, ZHAO Jiping and ZHANG Chengdong
(College of Animal Science and Technology, Northwest Aamp;F University, Yangling Shaanxi 712100,China)
The study aims to exploreSRYgene sequence of Chinese domestic bactrian camel and the copy number variation of SRY among populations. Partial sequence ofSRYgene of 94 samples from seven populations were sequenced to analyze the similarity among species. Additionally, CYP2A gene as a reference gene,SRYgene copy number was detected by real-time PCR. The similarity ofSRYgene between Chinese bactrian camel and human, horse, pig, sheep as well as bovine was 67.7%-68.1%, 68.5%-69.4%, 73.8%-74.4%, 73.8%-74.8% and 74.1%-74.8%, respectively. Except for one copy, the similarity between HMG of the rest copy of Chinese bactrian camel and HMG of alpaca, human, horse, pig, sheep as well as bovine was 97.3%, 84.2%, 85.6%, 86.3%, 87.0% and 85.6%, respectively. TheSRYgene was a multicopy gene in Chinese domestic bactrian camel and copy number variation ranged from 1 to 9. HMG sequences of Chinese domestic bactrian camel were highly conserved. TheSRYgene was a multicopy gene in Chinese domestic bactrian camel.
Domestic bactrian camel;SRYgene; Copy number
2016-09-20
2016-10-25
The National Natural Science Foundation of China (No. 31172178).
CHEN Huiling, female, master student. Research area:evaluation, protection and utilization of animal genetic resources. E-mail:chlachj@nwsuaf.edu.cn
日期:2017-11-17
網絡出版地址:http://kns.cnki.net/kcms/detail/61.1220.S.20171117.1101.004.html
2016-09-20
2016-10-25
國家自然科學基金(31172178)。
陳慧玲,女,碩士研究生,從事動物遺傳資源評價、保護和利用研究。E-mail:chlachj@nwsuaf.edu.cn
任戰軍,男,副教授,主要從事經濟動物研究。E-mail:renzhanjun@nwsuaf.edu.cn
Q951
A
1004-1389(2017)11-1569-08
