馬鈴薯Y病毒對不同馬鈴薯品種的致病力
2017-11-29 03:28:40白艷菊韓樹鑫高艷玲范國權邱彩玲聶先舟文景芝
西北農業學報 2017年11期
關鍵詞:癥狀
白艷菊,韓樹鑫,高艷玲,張 威,范國權,邱彩玲,申 宇,張 抒,尚 慧,聶先舟,文景芝
(1.東北農業大學 農學院,哈爾濱 150030;2.黑龍江省農業科學院 植物脫毒苗木研究所,哈爾濱 150086;3.加拿大農業部弗里得里克頓研究中心,新不倫瑞克 弗雷德里克頓 E3B 4Z7)
馬鈴薯Y病毒對不同馬鈴薯品種的致病力
白艷菊1,2,韓樹鑫1,高艷玲1,2,張 威1,2,范國權2,邱彩玲2,申 宇2,張 抒2,尚 慧1,聶先舟3,文景芝1
(1.東北農業大學 農學院,哈爾濱 150030;2.黑龍江省農業科學院 植物脫毒苗木研究所,哈爾濱 150086;3.加拿大農業部弗里得里克頓研究中心,新不倫瑞克 弗雷德里克頓 E3B 4Z7)
為明確不同馬鈴薯Y病毒(PVY)株系對馬鈴薯的影響及危害,采用PVYN:O、PVYN-Wi、PVYNTN-NW(SYRⅠ)和PVYNTN-NW(SYRⅡ)株系分離物人工侵染馬鈴薯,并通過透射電子顯微鏡觀察經PVY侵染后馬鈴薯植株細胞的超微結構。結果表明,敏感品種更適用于PVY病毒致病力鑒定,敏感品種‘克新13’和‘克新18’對PVY不同分離物的反應強烈,差異顯著,而‘興加2號’對4個PVY分離株均表現耐病,雖然細胞內均出現葉綠體變形、單層膜小囊泡和典型的風輪狀內含體,但癥狀只表現為不同程度的花葉。同時,發現PVYN-Wi致病力較弱,侵染后,4個馬鈴薯品種均出現花葉癥狀,細胞內部產生多膜結構、風輪狀內含體,引起敏感品種‘克新13’和‘克新18’細胞變形、葉綠體變形和髓鞘樣結構;PVYNTN-NW致病力最強,感病的‘克新13’和‘克新18’植株受害癥狀明顯加重,且植株早衰、細胞破壞嚴重,有些死亡較快的植株甚至未產生特征性內含體結構;PVYN:O致病力中等,植株受害癥狀和超微結構變化也介于PVYN-Wi和 PVYNTN-NW之間。不同馬鈴薯品種對PVY病毒的感病程度有較大差異,‘興加2號’為耐病品種。
馬鈴薯Y病毒(PVY);株系;馬鈴薯品種;超顯微結構;癥狀
馬鈴薯Y病毒(Potato virus Y,PVY)是馬鈴薯生產中最常見的一種馬鈴薯病害,染病馬鈴薯植株常出現品種退化、產量減少、品質變差等癥狀。PVY與其他病毒復合侵染后,可致馬鈴薯最高減產80%,經濟損失嚴重。PVY屬于單鏈正義RNA病毒,其基因組長約9.7 kb。侵染馬鈴薯的PVY病毒主要可分為PVYO、PVYC、PVYN、PVYZ和PVYE等5個株系[1]。研究發現,PVYO、PVYN以及兩者重組衍生的新的株系侵染較為普遍[2],而有關其他株系侵染的報道極少。目前,已報道的PVY重組株系有PVYN:O、PVYN-Wi、PVYNTN-NW,有學者對PVY有規律的重組變化細化歸類為A、B和Ⅰ、Ⅱ、Ⅲ幾種類型,本研究只選擇其中有代表性的PVYN:O、PVYN-Wi、PVYNTN-NW(SYRⅠ)和PVYNTN-NW(SYRⅡ)等株系的分離物進行研究。
自然界中,對PVY免疫的馬鈴薯品種較少,抗PVY病毒的品種選育和應用則較為普遍。受PVY侵染的馬鈴薯植株常表現出多種不同癥狀,如花葉、皺縮、壞死等,侵染嚴重時可導致植株矮化或早衰,癥狀的差異是由于病毒對植物細胞功能的破壞產生的。目前,多采用透射電子顯微鏡技術進行病原鑒定和病原致病力研究。自Kausche和Melcher首次通過電鏡觀察到煙草花葉病毒(TMV)以來,電鏡技術已成為植物病毒研究必不可少的常規手段之一[3],Maramorosch和Franchi在其專著中通過照片的方式為廣大研究者提供病毒診斷的參照,從而使越來越多的植物病毒在不同植物中被發現,可以很清楚地觀測到病毒的形態和結構。另外,病毒在寄主細胞中的狀態、寄主細胞的變化也同樣可以被直觀發現和記錄[4-5]。1996年,胡向武等[6]利用透射電子顯微鏡對受煙草花葉病毒侵染的煙草細胞進行研究,發現其細胞中葉綠體發生腫脹、空泡化、片層結構疏松、散失甚至外膜破碎等現象。2004年,彭晏輝等[7]利用透射電鏡技術對受PVY侵染的馬鈴薯葉綠體進行研究,并以此分析侵染后對其植物光合作用的影響。
由于PVY為多株系,且分子遺傳多樣性豐富,PVY在基因水平上的差異與其致病能力有一定相關性,聶碧華[8]鑒定PVYO株系的2個分離株,在核酸和蛋白同源性分別為97.1%和98.5%的情況下,致病力表現出較大差異。本研究通過使用不同遺傳背景的PVY對4個品種的馬鈴薯進行接種,并通過對侵染后的馬鈴薯細胞進行超薄切片,然后用透射電鏡進行分析,擬通過表觀癥狀與超微結構下的表現相對照,找到不同PVY株系癥狀與超微結構相對應的關系,同時通過病毒對馬鈴薯細胞的破壞規律找到不同株系PVY致病力差異,為PVY的研究及防治提供理論依據。
1 材料與方法
1.1 試驗材料
‘克新13’‘克新18’‘費烏瑞它’和‘興加2號’4個馬鈴薯品種原原種均由黑龍江省農業科學植物脫毒苗木研究所提供,種子大小均勻,質量約10 g。接種采用的PVY毒源為遺傳背景不同的4個PVY不同株系的分離物:HLJ-BDH-2(PVYN-Wi)、IMN-2013-W-369-12[PVYNTN-NW(SYRⅡ)]、HLJ-30-2(PVYN:O)和HLJ-9-4[PVYNTN-NW(SYRⅠ)],4個分離株的重組位點見圖1。
以上材料均由黑龍江省農業科學院植物脫毒苗木研究所保存并提供。

圖1 不同株系PVY基因結構差異Fig.1 Genetic structure difference of different strains of PVY
1.2 試驗方法
1.2.1 PVY病毒接種 每個品種的馬鈴薯均種植50株,共計200株,溫室保持20~26 ℃,每日光照14 h。當馬鈴薯植株生長至4~6葉期時,采用摩擦接種的方法將4個PVY分離物分別接種至馬鈴薯葉片,具體方法:將帶有病毒的植物葉片放于磷酸緩沖液(PB)研磨[m(攜病毒葉片)∶V(緩沖液)=1∶10],研磨后2 000 r/min離心,上清液即為研究所用的病毒混合液。每株植物均使用約300 μL的病毒混合液,每株分別接種2片葉片,4個馬鈴薯品種均分別接種不同的PVY株系分離物,每處理3次重復。于接種后第1天即開始,每天觀察癥狀并記錄。接種后的第3天、第6天、第9天和第12天分別取植株新生葉片檢測,以確定接種是否成功,檢測采用DAS-ELSIA的方法。
1.2.2 超薄切片樣品的采集及前固定 馬鈴薯接種24 d后,采集植株自上往下第4片完全展開的葉片,并按電子顯微鏡樣品固定的要求裁剪樣品[9],迅速放入含φ=3%戊二醛的固定液進行固定。
1.2.3 超薄切片樣品的制備 將經過前固定處理的樣品,用1 g/mL四氧化鋨固定液在室溫下固定3 h,再經濃度梯度酒精及環氧丙烷等處理后,用820樹脂進行包埋,并于37 ℃孵育4 h,70 ℃ 過夜聚合。聚合完成后,對包埋塊進行切片,從而獲得可使用透射電子顯微鏡觀察的超薄切片(厚70 nm),最后采用醋酸雙氧鈾飽和溶液及檸檬酸鉛2種溶液的雙染法對超薄切片進行染色及觀察。
2 結果與分析
2.1 不同品種馬鈴薯植株受PVY侵染后癥狀差異
DAS-ELISA檢測結果顯示,各PVY株系分離物均能成功侵染4個馬鈴薯品種,并使其獲毒,且不同PVY分離物在馬鈴薯品種上表現的生理癥狀差異較大。其中,經PVYN-Wi株系侵染的4個馬鈴薯品種表現的生理癥狀均較輕微,僅表現出花葉癥狀。SYRⅠ和SYRⅡ可致‘費烏瑞它’‘克新13’和‘克新18’3個馬鈴薯品種產生花葉、主莖壞死、莖壞死和葉片黑色點斑等癥狀,且‘克新13’和‘克新18’植株受害癥狀更明顯,出現主莖壞死、葉片大面積黑色點斑、垂葉枯萎以及更快的發病速度等,甚至植株過早死亡。其中,SYRⅠ可導致‘克新13’產生花葉、主莖壞死、莖壞死和葉片黑色點斑等癥狀,卻不能致其過早死亡,但能使‘克新18’過早死亡;SYRⅡ可導致‘克新13’快速死亡(表1)。PVYN:O可導致‘克新13’‘克新18’及‘費烏瑞它’植株主莖壞死、葉脈壞死,并使部分葉片出現黑色點斑,而‘克新13’于接種PVYN:O1個月后死亡,致病能力比PVYN-Wi強,但弱于PVYNTN-NW(表1,圖2)。

表1 接種不同PVY株系后馬鈴薯植株癥狀Table 1 Symptoms of different potato varieties inoculated with different strains of PVY
注:接種時間為2015-09-04。M.花葉;SSN.主莖壞死;VN.葉脈壞死;BSL.葉片黑色點斑;DPL.垂葉干枯;DP.植株死亡;PTNRD .薯塊環斑壞死。a.植株死亡時間為接種后30 d;b.植株死亡時間為接種后15 d;c 植株死亡時間為接種后25 d。
Note:Infect time is 2015-09-04.M.Mosaic; SSN.Stem streak necrosis; VN. Necrosis on leaves veins; BSL.Black spots on leaves; DPL.Dry and pendulous leaves; DP.Dead plants; PTNRD.Potato tuber necrotic ringspot disease.a.Time of plant death is 30 days after inoculated with virus; b.Time of plant dead is 15 days after inoculated with virus; c.Time of plant dead is 25 days after inoculated with virus.

A.接種PVYN:O后,‘克新13’葉片出現花葉 Mosaic after ‘Kexin 13’ inoculated with PVYN:O;B.接種PVYN:O后,‘克新13’主莖壞死 Stem streak necrosis after ‘Kexin 13’ inoculated with PVYN:O;C.接種PVYNTN-NW(SYRⅠ)后,‘費烏瑞它’葉片葉脈壞死 Veins necrosis after ‘Favorita’ inoculated with PVYNTN-NW(SYRⅠ);D.接種PVYNTN-NW(SYRⅡ)后,‘克新13’葉片出現黑色點斑 Black spots after ‘Kexin 13’ inoculated with PVYNTN-NW(SYRⅡ) on leaves;E.接種PVYNTN-NW(SYRⅠ)后,‘克新18’植株葉片垂葉干枯 Dry and pendulous leaves after ‘Kexin 18’ inoculated with PVYNTN-NW(SYRⅠ);F.接種PVYNTN-NW(SYRⅡ)后,‘克新13’植株死亡 Plants was dead after ‘Kexin 13’ inoculated with PVYNTN-NW(SYRⅡ);G.接種PVYNTN-NW(SYRⅠ)后,‘克新18’薯塊產生環斑壞死 Potato tuber necrotic ringspot disease (PTNRD) in the tuber after ‘Kexin 18’ inoculated with PVYNTN-NW(SYRⅠ);H.未接種病毒的對照植株 Negative control that not inoculated with PVY
圖2接種PVY后馬鈴薯植株癥狀
Fig.2SymptomsofdifferentpotatovarietiesinoculatedwithdifferentstrainsofPVY
4個品種,‘中克新13’‘克新18’ 和‘費烏瑞它’均較敏感,感染不同株系后發病癥狀差異明顯,致病力弱的癥狀溫和,致病力強的壞死嚴重,甚至植株死亡;而‘興加2號’耐病性較好,感染后僅出現花葉癥狀,不同PVY類型造成的癥狀差異也僅是花葉輕重程度的差異。
2.2透射電子顯微鏡觀察受侵染植株細胞超微結構差異
PVY侵染后,馬鈴薯細胞的超顯微結構發生改變,不同PVY株系病毒對不同馬鈴薯品種的細胞傷害有較大差異,這些差異體現在多個方面,不僅對植物細胞整體結構有影響,對各細胞器也有不同程度的破壞。其中,‘克新13’的細胞超顯微結構中,風輪狀內含體的數量較多,且細胞中表現較為明顯,這種現象在PVYN-Wi、PVYN:O、PVYNTN-NW(SYRⅠ)株系侵染后的馬鈴薯細胞中均可發現。其他2個品種則與‘克新13’相反,相同研究條件下,受PVY侵染后,‘費烏瑞它’植株細胞中幾乎未觀察到有產生風輪狀內含體的獨特標志性結構。另外,在其他2個品種植株細胞中觀察到風輪狀內含體的數量則與侵染的PVY分離物有關, PVYNTN-NW(SYRⅡ)和PVYN-Wi會導致‘費烏瑞它’和‘興加2號’植株細胞中出現風輪狀內含體,并較明顯,而其他品種則未在細胞中發現有這種特異性結構(表2和圖3)。

表2 接種不同PVY株系后馬鈴薯細胞超微結構Table 2 Different symptoms of cellular ultrastructure after PVY infected with different strains of PVY
注:PW .風輪狀內含體;LIC.線狀內含體;CD.細胞變形;ChD.葉綠體變形;ChDe.葉綠體裂解;MB.髓鞘樣結構;M.多泡體結構;PM.多膜結構。
Note: PW.Pinwheels; LIC.Laminate inclusion components; CD.Cell deformation; ChD.Chloroplast deformation; ChDe.Chloroplast degradation; MB.Myeloid body; M.Multivesicular; PM.Proliferated membranes.
PVY病毒對馬鈴薯細胞的破壞及細胞內細胞器的形態結構的變化,會導致植株產生相應的癥狀,用來分析PVY對馬鈴薯植株的危害。研究發現,所有受PVY侵染的馬鈴薯植株均會在細胞中產生或多或少、或大或小的改變。如在‘克新13’和‘克新18’中,無論受到何種PVY分離物侵染后,該品種的細胞均會表現出細胞變形(CD)、葉綠體變形(ChD)或多膜結構(PM)。而當‘克新13’受SYRⅡ侵染后,該品種的細胞變化更加明顯,葉綠體發生大面積的變形及裂解(ChDe),同時,細胞中其他細胞器的形態也發生改變(如線粒體聚集、多泡體結構、多膜結構等),說明細胞結構已經遭到破壞。與‘克新13’相似的是,在‘費烏瑞它’和‘興加2號’植株細胞中,均可發現由PVY侵染引起的各種現象,如葉綠體變形、結構被破壞、出現單層膜結構小泡等現象,這些現象或多或少的出現在各品種的馬鈴薯細胞中(表2)。
2.3 生理癥狀與超顯微結構的相關性
觀察馬鈴薯植株癥狀及其細胞的超顯微結構發現,植株表現出較嚴重的癥狀時,其細胞器的受害表現更為嚴重,有些細胞的細胞器甚至出現破裂、消融、不完整,且細胞形態不正常,并且在部分馬鈴薯細胞中發現較多的風輪狀內含體、線狀內含體等,這是僅有在PVY病毒侵染后才能發現的特異性結構;當植株受害癥狀表現較輕微時,細胞的細胞器結構均較為完整,同時,很少在這些馬鈴薯細胞中發現風輪狀內含體、線狀內含體等標志性結構。‘克新13’‘克新18’和‘費烏瑞它’對PVYN:O、PVYNTN-NW(SYR I)、PVYNTN-NW(SYRⅡ)3個株系的侵染表現較敏感,受害后植株發病嚴重。觀察它們對應的細胞超顯微結構,發現僅在PVYN:O和PVYNTN-NW(SYRⅠ)侵染‘克新13’,PVYNTN-NW(SYRⅡ)侵染‘克新18’和‘費烏瑞它’ 的植株細胞中發現風輪狀內含體(圖4),而不是所有樣品中均能觀察到風輪狀內含體等較為明顯的特征結構(表2)。經4個不同PVY株系侵染后,‘興加2號’植株均未表現出明顯的生理癥狀,而僅出現花葉。觀察‘興加2號’細胞的超顯微結構,也未發現其細胞器受到較嚴重的破壞,同時僅在受PVYN-Wi侵染的植株細胞中觀察到風輪狀內含體結構,說明植株癥狀表現與細胞內觀察結果密切相關。
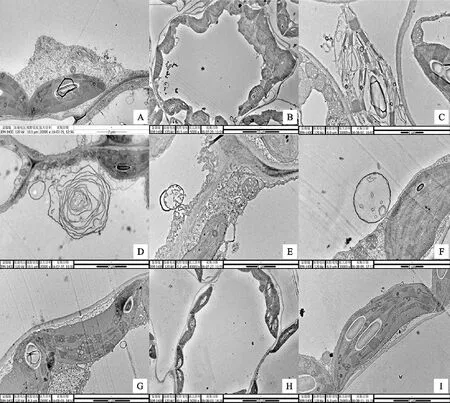
A.PVYN-Wi侵染后,‘克新13’產生風輪狀內含體,×20 000 After being infected by PVYN-Wi,‘Kexin 13’ was found to be with pinwheel inclusion in leaf cells,×20 000;B.PVYNTN-NW(SYRⅠ)侵染后,‘克新13’細胞形態發生改變,×5 000 Cellular morphology of ‘Kexin 13’ after being infected by PVYNTN-NW,×5 000;C.PVYNTN-NW(SYRⅠ)侵染后,‘克新13’葉綠體變形并裂解,×30 000 Cellular chloroplast morphology changes and lysis of ‘Kexin 13’ after being infected by PVYNTN-NW(SYRⅠ),×30 000;D.PVYNTN-NW(SYRⅠ)侵染后,‘興加2號’細胞內的髓鞘樣結構,×20 000 After being infected by PVYNTN-NW,‘Xingjia 2’ was found to be with myeloid body inclusion in leaf cells,×20 000;E.PVYN-Wi侵染后,‘克新13’及‘費烏瑞它’細胞內出現多膜結構,×40 000 After being infected by PVYN-Wi,cells of ‘Kexin 13’ and ‘Favorita’ have multi-film structure,×40 000,;F. PVYN-Wi侵染后,‘克新13’及‘費烏瑞它’細胞內出現多膜結構,×30 000 After being infected by PVYN-Wi,cells of Kexin 13 and Favorita have multi-film structure,×30 000;G,I.未接種PVY的正常馬鈴薯植株細胞結構,×25 000 The negative control that did not inoculated with PVY,×25 000;H.未接種PVY的正常馬鈴薯植株細胞結構,×5 000 The negative control that did not inoculated with PVY,×5 000
圖3PVY侵染后馬鈴薯植株細胞的超顯微結構
Fig.3PerformanceofmicrostructurewithdifferentpotatovarietiesinoculatedwithdifferentstrainsofPVY
3 討 論
PVY作為最常見的馬鈴薯病毒病,受到高度關注。大多數研究者通過觀察寄主植物受病原侵染后產生的癥狀及相應的血清學檢測、分子生物學檢測結果,從而判斷馬鈴薯植株受病原侵害的情況。如Ali等[10]利用薯塊癥狀及對PVY全序列測序,確定新的PVY株系,Nie[11]利用多種方式測試PVY的侵染途徑,并通過癥狀判斷及血清學檢測確定PVY的侵染方式為機械摩擦傳播。目前,采用電子顯微鏡技術研究PVY病毒的文獻較少[11]。徐玲等[12]觀察經煙草假單胞桿菌侵染的煙草細胞的超微結構,發現侵染后植株細胞葉綠體破壞嚴重,接種2 d后甚至出現葉綠體完全裂解的現象。另外,在接種后期,細胞中大量的細胞器也發生解體,甚至細胞僅留有空腔。劉國順等[13]采用電子顯微鏡等技術進行觀察,發現經花葉病毒侵染后的煙草植株及葉片細胞的保護酶活性及細胞中細胞器發生變化,電鏡技術為病理研究提供一條直觀的路徑。

A.PVYN:O侵染后,‘克新13’葉片出現花葉 Mosaic after ‘Kexin 13’ inoculated with PVYN:O;B.PVYNTN-NW(SYRⅠ)侵染后,‘克新18’葉片出現花葉 Mosaic after ‘Kexin 18’ inoculated with PVYNTN-NW(SYRⅠ);C.PVYNTN-NW(SYRⅡ)侵染后,‘費烏瑞它’葉片出現花葉 Mosaic after Favorita inoculated with PVYNTN-NW(SYRⅡ);D.PVYN-Wi侵染后,‘興加2號’葉片出現花葉 Mosaic after ‘Xingjia 2’ inoculated with PVYN-Wi;E.PVYN:O侵染后,‘克新13’主莖壞死 Stem streak necrosis after‘ Kexin 13’ inoculated with PVYN:O;F.PVYNTN-NW(SYRⅠ)侵染后,‘克新18’主莖壞死、垂葉干枯 Stem streak necrosis and dry and pendulous leaves after ‘Kexin 18’ inoculated with PVYNTN-NW(SYRⅠ);G.PVYNTN-NW(SYRⅠ)侵染后,‘費烏瑞它’葉片葉脈壞死 Veins necrosis after ‘Favorita’ inoculated with PVYNTN-NW(SYRⅠ);H.PVYN:O侵染后,‘興加2號’葉片出現花葉 Mosaic after ‘Xingjia 2’ inoculated with PVYN:O;I.PVYN:O侵染后,‘克新13’葉片細胞出現風輪狀內含體,×30 000 After infected by PVYN:O,‘Kexin 13’ was found to be with pinwheel inclusion in leaf cells,×30 000;J.PVYNTN-NW(SYRⅡ)侵染后,‘克新18’葉片細胞出現風輪狀內含體,×30 000 After infected by PVYNTN-NW(SYRⅡ),‘Kexin 18’ was found to be with pinwheel inclusion in leaf cells,×30 000;K.PVYNTN-NW(SYRⅡ)侵染后,‘費烏瑞它’葉片細胞出現風輪狀內含體,×30 000 After infected by PVYNTN-NW(SYRⅡ),‘Favorita’ was found to be with pinwheel inclusion in leaf cells,×30 000;L.PVYN-Wi侵染后,‘興加2號’細胞出現風輪狀內含體,×25 000 After infected by PVYN-Wi,‘Xingjia 2’ was found to be with pinwheel inclusion in leaf cells,×25 000;M.PVYN:O侵染后,‘克新13’細胞形態,×3 000 Cellular morphology of ‘Kexin 13’ after infected by PVYN:O,×3 000;N.PVYNTN-NW(SYRⅠ)侵染后,‘克新18’細胞出現多膜結構,×15 000 After infected by PVYNTN-NW(SYRⅠ),cells of ‘Kexin 18’ have multi-film structure,×15 000;O.PVYNTN-NW(SYRⅠ)侵染后,‘費烏瑞它’細胞葉綠體形態發生改變,×25 000 Change of cellular chloroplast morphology in ‘Favorita’ infected by PVYNTN-NW(SYRⅠ),×25 000;P.PVYN:O侵染后,‘興加2號’細內葉綠體形態改變,×25 000 Cellular chloroplast morphology changes in ‘Xingjia 2’ infected by PVYN:O,×25 000
圖4不同品種馬鈴薯接種后癥狀及超微結構差異
Fig.4DifferencesamongdifferentpotatovarietiesinoculatedwithdifferentstrainsofPVY
本研究通過觀察不同株系PVY侵染后馬鈴薯植株癥狀及超顯微結構的差異,從而掌握不同株系PVY對馬鈴薯品種的危害。結果顯示,經SYRⅡ株系侵染后,‘克新13’植株受害癥狀表現最為明顯,整株植株提前死亡。通過透射電鏡觀察‘克新13’細胞的超顯微結構,發現該植株細胞的葉綠體變形,且裂解程度明顯較其他品種及株系侵染后要嚴重,這可能是導致該品種提前死亡的原因之一。彭晏輝等[7]研究認為,經PVY侵染后,馬鈴薯細胞的葉綠體會出現過早成熟和早衰,進而導致植株光合作用能力下降;郭興啟等[14]利用PVY侵染煙草以研究PVY與其他病毒的協生作用,發現PVYN株系單獨侵染后會導致植物細胞中出現風輪狀內含體,而受損的細胞還會出現葉綠體嚴重損傷、髓鞘樣結構、多膜結構等明顯特征。本研究中,在經不同PVY株系侵染的馬鈴薯植株中也觀察到與上述研究相同的現象;但不同的是,并不是在所有的植株細胞中均能發現這種受PVY侵染后的標志性結構——風輪狀內含體,這可能是由于不同品種的抗PVY能力有差異;同時,不同遺傳背景的PVY分離物的致病能力也有差異,不同PVY病毒分離物與不同馬鈴薯品種的發病的速度、癥狀與細胞內部結構變化建立某種聯系。
本研究中,未出現風輪狀內含體結構的樣品主要有‘克新13’,其僅在接種SYRⅡ時,發病迅速,出現細胞器裂解、細胞膜破裂等現象,植株快速死亡,這是因為接種的時期為苗期,植株抗性較弱,反應比較強烈。在其他品種中,植株表現出不同的發病程度,細胞中出現風輪狀內含體、多泡(多膜)結構和髓鞘狀結構。尤其是‘興加2號’,植株長勢正常,癥狀輕微,細胞破壞也比其他品種輕微。內含體和多膜、多泡狀結構的形成很可能是對PVY病毒抗性的一種反映,這一系列變化過程,可能將病毒毒力部分化解、轉移,使病毒的致病力弱化,也是細胞內病毒與細胞防御體系斗爭的結果。接種SYRⅡ的‘克新13’則是由于病毒對細胞的危害巨大,導致其細胞迅速衰亡,細胞內尚未產生有效的防御反應;植株只表現受害癥狀而未死亡的,是因為細胞正常代謝雖受到影響,但尚能維持病毒破壞與光合作用的平衡。
本研究通過對不同馬鈴薯品種及不同PVY株系的病毒癥狀和超微結構變化進行研究,很好地揭示品種抗性的原因,發現在不同PVY株系作用下,馬鈴薯植株的表觀癥狀與細胞超微結構間有密切聯系。且發現所研究的4個品種有明顯的抗性異, ‘克新13’在苗期高感PVY,在其他資料里‘克新13’均表現為PVY田間過敏抗性[15],‘克新18’和‘費烏瑞它’也表現為感病品種,而‘興加2號’表現出顯著的耐病性。該結果為進一步研究PVY致病機理提供有關病毒與寄主互作的細胞病理學資料,并為PVY的防治及鑒定提供一定的生物學及影像學參考。
Reference:
[1] SINGH R P,VALKONEN J P,GRAY S M,etal.Discussion paper:the naming of potato virus Y strains infecting potato[J].ArchivesofVirology,2008,153(1):1-13.
[2] NIE B,SINGH M,MURPHY A,etal.Response of potato cultivars to five isolates belonging to four strains of potato virus Y[J].PlantDisease,2012,96(10):1422-1429.
[3] MARTELLI G P,RUSSO M.Use of thin sectioning for visualization and identification of plant viruses[J].MethodsinVirology,1984,8:143-224.
[4] MARAMOROSCH K .The Atlas of Insect and Plant Viruses,Including Mycoplasmaviruses and Viroids[M].New York:Academic Press,1977:1-30
[5] FRANCKI R I B,MILNE R G,HATTA T.Atlas of Plant Viruses[M].Boca Raton:CRC Press,1985:39-49.
[6] 胡向武,唐劍云,張林普.煙草花葉病毒的侵染部位及細胞病理變化的電鏡觀察[J].安徽師范大學學報(自然科學版),1996,19(2):140-143.
HU X W,TANG J Y,ZHANG L P.Observation on the infected position of TMV particles and pathologic changes of cells under the electron microscope[J].JournalofAnhuiNormalUniversity(NaturalScienceEdition),1996,19(2):140-143 (in Chinese with English abstract).
[7] 彭晏輝,雷娟利,黃黎鋒,等.馬鈴薯Y病毒侵染對葉綠體超微結構、光合和熒光參數的影響[J].植物病理學報,2004,34(1):32-36.
PENG Y H,LEI J L,HUANG L F,etal.Effects of potato virus Y infection on chloroplast ultrastructure,photosynthesis and chlorophyll fluorescence quenching in potato leaves[J].ActaPhytopathologicaSinica,2004,34(1):32-36 (in Chinese with English abstract).
[8] 聶碧華.馬鈴薯Y病毒新變異株系的克隆鑒定及馬鈴薯-病毒互作機制研究[D].武漢:華中農業大學,2010:100-101.
NIE B H.Cloning and reeognition of novel variant of potato virus Y and preliminary exploring of mechanism in potato-virus interaetions[D].Wuhan:Huazhong Agriculturail University,2010:100-101 (in Chinese with English abstract).
[9] 郭 欣,申家恒,王艷杰,等.水稻雙受精過程的超微結構觀察[J].植物學報,2013,48(4):429-437.
GUO X,SHEN J H,WANG Y J,etal.Ultrastructure observation of double fertilization inOryzasativa[J].ChineseBulletinofBotany,2013,48(4):429-437 (in Chinese with English abstract).
[10] ALI M C,MAOKA T,NATSUAKI T,etal.PVYNTN-NW,a novel recombinant strain of potato virus y predominating in potato fields in Syria[J].PlantPathology,2010,59(1):31-41.
[11] NIE X.Detection of molecular markers linked toRygenes in potato germplasm for marker-assisted selection for extreme resistance to PVY in AAFC’s potato breeding program[J].CanadianJournalofPlantScience,2016,96(5):737-742.
[12] 徐 玲,張世珖.煙草野火病毒素對煙草葉片組織超微結構的影響[J].云南農業大學學報,2006,21(1):57-60.
XU L,ZHANG SH G.The effect of tabtoxin to the ultra-structure of tobaccos’ leaves[J].JournalofYunnanAgriculturalUniversity,2006,21(1):57-60 (in Chinese with English abstract).
[13] 劉國順,張彩霞,王嬋娟,等.受烤煙花葉病病毒侵染后葉中保護酶活性和超微結構的變化[J].植物生理學報,2009,45(5):459-463.
LIU G SH,ZHANG C X,WANG CH J,etal.Changes in protective enzyme activities and ultrastructures of flue-cured tobacco infected with tobacco mosaic virus[J].PlantPhysiologyCommunications,2009,45(5):459-463(in Chinese with English abstract).
[14] 郭興啟,馮 炘,李向東,等.PVY/PVX協生作用對病毒濃度及寄主細胞超微結構的影響[J].中國農業科學,2003,36(3):281-286.
GUO X Q,FENG X,LI X D,etal.Effect of synergism between PVY and PVX on the viruses titer and the ultrastructure of infected cells of tobacco plant[J].ScientiaAgriculturaSinica,2003,36(3):281-286 (in Chinese with English abstract).
[15] 洪乃武,張 生,林長春,等.馬鈴薯新品種‘克新13號’育成報告[J].中國馬鈴薯,1999(3):188.
HONG N W,ZHANG SH,LI CH CH,etal.Report on the breeding of new potato variety ‘Kexin 13’[J].ChinesePotatoJournal,1999(3):188(in Chinese).
CorrespondingauthorWEN Jingzhi,female,professor,doctoral supervisor.Research area:plant pathology.E-mail: jzhwen2000@163.com
(責任編輯:郭柏壽Responsibleeditor:GUOBaishou)
PathogenicityofPotatoVirusYStrainsinDifferentPotatoVarieties
BAI Yanju1,2,HAN Shuxin1,GAO Yanling1,2,ZHANG Wei1,2, FAN Guoquan2,QIU Cailing2,SHEN Yu2,ZHANG Shu2, SHANG Hui1,NIE Xianzhou3and WEN Jingzhi1
(1.College of Agriculture,Northeast Agricultural University,Harbin 150030,China; 2.Virus-free Seeding Research Institute of Heilongjiang Academy of Agricultural Sciences,Harbin 150086,China; 3.Fredericton Research and Development Centre,Agriculture and Agri-Food Canada,Fredericton,New Brunswick E3B 4Z7)
Potato virus Y (PVY) is one of the most common and worldwide destructive viruses that affects potato production.To unveil the effects and the pathogenicity of different strains of PVY in different potato varieties,four natural recombinant PVY strains including PVYN:O,PVYN-Wi,PVYNTN-NW(SYR-Ⅰ) and PVYNTN-NW(SYR-Ⅱ) were mechanically inoculated onto four potato varieties.In addition to the visual foliar and tuber symptom observations,the ultrastructure and cellular morphology changes were also monitored by electron microscopy.Generally,PVYN-Wicaused mild symptoms (mainly mosaic) on all varieties,whereas the remaining strains caused severe foliar and/or tuber symptoms.The variety ‘Xingjia 2’ exhibited mild foliar symptoms compared with other varieties especially ‘Kexin 13’ and ‘Kexin 18’ upon infection with different strains of PVY.Interestingly,in spite of the fact that PVYN-Wionly caused mild symptoms in all varieties,it induced cellular morphological in varying-degrees and changed ranging from pinwheel formation in ‘Xingjia 2’ to pinwheel formation,chloroplast degradation,membrane proliferation and cell deformation in ‘Kexin 18’.The pathogenicity of PVYNTN-NW(SYR-Ⅰ and SYR-Ⅱ) was the strongest in the tested strains,which not only caused serve foliar symptoms in ‘Kexin 13’,‘Kexin 18’ and ‘Favorita’,but also incited necrotic ringspots in tubers of at least two varieties (i.e.,‘Kexin 13’ and ‘Xingjia 2’).The pathogenicity of PVYN:Ofalled between PVYN-Wiand PVYNTN-NW,and it caused intermediate level of symptoms in plants and intermediate degree of changes in ultrastructure.This study illuminated the interactions between different PVY strains and different potato varieties in both whole-plant and ultrastructural levels,thus this provided insights for better management of this destructive virus in potato crops.
Potato virus Y (PVY); Potato varieties; Strains; Symptoms; Ultrastructure
2017-01-18
2017-04-06
Modern Agriculture Potato Industry Technology System(No.CARS-10-P14).
BAI Yanju,female,research fellow,doctoral student.Research area:plant pathology.E-mail: yanjubai@163.com
日期:2017-11-17
網絡出版地址:http://kns.cnki.net/kcms/detail/61.1220.S.20171117.1101.036.html
2017-01-18
2017-04-06
國家現代農業馬鈴薯產業技術體系(CARS-10-P14)。
白艷菊,女,研究員,博士研究生,研究方向為植物病理。E-mail:yanjubai@163.com
文景芝,女,教授,博士生導師,研究方向為植物病理。E-mail:jzhwen2000@163.com
S435.32
A
1004-1389(2017)11-1713-08
猜你喜歡
初中生學習指導·提升版(2023年8期)2023-09-12 10:26:19
保健醫苑(2022年1期)2022-08-30 08:39:40
中老年保健(2021年12期)2021-08-24 03:30:44
今日農業(2020年17期)2020-10-27 03:10:52
今日農業(2020年16期)2020-09-25 03:05:08
家庭醫學(下半月)(2020年2期)2020-05-11 02:07:18
基層中醫藥(2020年10期)2020-02-13 15:45:52
吉林蔬菜(2017年10期)2017-11-01 07:47:04
獸醫導刊(2016年6期)2016-05-17 03:50:35
中國醫學影像學雜志(2015年9期)2015-12-15 11:03:26
