CO2加富對鹽生杜氏藻(Dunaliella salina)葉綠素熒光參數的影響*
2017-12-09 01:20:54黃致遠趙新宇胡順鑫
海洋與湖沼 2017年5期
臧 宇 黃致遠 趙新宇 胡順鑫,
(1.中國海洋大學海洋生命學院 青島 266003;2.山東省海洋資源與環境研究院 煙臺 264006;3.山東省青島市第五十八中學 青島 266199)
CO2加富對鹽生杜氏藻(Dunaliella salina)葉綠素熒光參數的影響*
臧 宇1黃致遠3趙新宇1胡順鑫1,2①
(1.中國海洋大學海洋生命學院 青島 266003;2.山東省海洋資源與環境研究院 煙臺 264006;3.山東省青島市第五十八中學 青島 266199)
大氣中CO2濃度不斷升高導致的海水酸化,已經引起了廣泛的環境、生態和氣候問題。本實驗采用實驗生態學的方法,以鹽生杜氏藻(Dunaliella salina)為研究對象,分析其在CO2加富的條件下葉綠素熒光參數的變化。研究表明,CO2加富對鹽生杜氏藻光系統Ⅱ最大光化學量子產額(Fv/Fm)和最大相對電子傳遞速率(rETRmax)無顯著影響(P>0.05),顯著促進了光系統Ⅱ實際光合效率(P<0.05)和光能利用效率(α)(P<0.05),并且降低了飽和光強(Ek)(P<0.05)。然而,CO2升高增加了鹽生杜氏藻的光抑制參數(β)(P<0.05)和非光化學淬滅(NPQ)(P<0.05),這說明在光照充足的情況下,CO2加富會對鹽生杜氏藻產生負面效應,使其更容易受到光抑制。
CO2;海水酸化;鹽生杜氏藻;葉綠素熒光
因為煤炭、石油及天然氣等燃料的使用以及森林破壞的加劇,大氣中 CO2含量正急劇升高,并在環境、生態以及氣候等方面導致諸多問題(Hughes,2000)。海洋作為地球表面最大的碳源之一,可以吸收人類活動排放 CO2總量的一半,而 CO2排放量上升,也導致了一系列的負面效應,而這其中最嚴峻的一個海洋化學事件就是海洋酸化(Ocean Acidification)(Fabry,2008)。在以歐洲為代表的工業革命以前,海水中的pH為8.2,截至目前,海水pH下降了0.1個單位,此時大氣CO2濃度為390ppmv。根據推斷,到2100年,海表pH將下降 0.4個單位,pCO2將上升200%,屆時CO2濃度將達到1000ppmv (Caldeiraet al,2003;Doneyet al,2009)。
海洋酸化改變了海洋化學環境,截至目前,大量研究表明海洋酸化可以降低珊瑚、珊瑚藻類以及貝類的鈣化量(Gaoet al,1993);海洋酸化對于非鈣化生物也會產生影響,CO2濃度升高可以促進海洋硅藻三角褐指藻(Phaeodactylum tricornutum)的生長和光合作用,并且下調了其無機碳濃縮機制(Wuet al,2010)。同時,利用葉綠素熒光技術探討海洋酸化對藻類影響的研究也有很多:徐金濤等(2016)研究表明,CO2加富條件能夠顯著促進塔瑪亞歷山大藻(Alexandrium tamarense)PSⅡ最大光化學量子產量(Fv/Fm),提高最大相對電子傳遞效率(rETRmax)和光能轉換效率。當營養條件及光照條件適宜時,海洋酸化可促進壇紫菜(Pyropia haitanensis)與龍須菜(Gracilaria lemaneiformis)的光合作用,并在一定程度上有利于壇紫菜對病害的抗性(陳斌斌,2015)。
鹽生杜氏藻(Dunaliella salina)是一種單細胞綠藻,由于其結構簡單、可以在高鹽環境下生存,抗逆性較強。作為一種良好的模式生物,其已經被廣泛應用于生物工程和遺傳工程中。但關于鹽生杜氏藻對環境脅迫的基礎研究仍然不夠系統全面,尤其是鹽生杜氏藻光合作用對海洋酸化的響應尚未見報道。因此,本實驗以鹽生杜氏藻為實驗對象,擬利用葉綠素熒光技術,探究 CO2濃度升高對鹽生杜氏藻對葉綠素熒光參數的影響,以期為預測未來海洋酸化條件下海洋浮游植物的適應性提供理論基礎。
1 材料與方法
1.1 藻種來源及培養條件
實驗所用微藻——鹽生杜氏藻(Dunaliella salina)取自中國科學院海洋研究所微藻培養室,培養于改良后的f/2培養基中。所用的海水取自青島市魯迅公園附近的自然海水,經脫脂棉粗過濾、M-50抽濾器過濾(濾膜直徑 5cm,孔徑 0.45μm)細過濾,然后高壓滅菌。光照強度為80μmol/(m2·s),光暗周期比為12h︰12h,培養溫度(22±1)°C,鹽度為32。
1.2 實驗設計
實驗選取了兩種 CO2濃度的氣體,分別為390ppmv(目前大氣中CO2濃度)和1000ppmv (2100年的預測值)。390ppmv濃度的CO2取自室外空氣(氣體采集處應注意通風);1000ppmv濃度的CO2氣體是高純二氧化碳氣體(99.99%)和空氣在二氧化碳光照培養箱(HP400G,瑞華,武漢)內混合而成,二氧化碳光照培養箱可以控制培養箱內CO2氣體的濃度。借助空氣壓縮泵將達到預設濃度的CO2氣體通過0.22μm的針式過濾器泵入培養瓶內,以保證通入培養液的氣體無菌。實驗時先將目標微藻培養至指數生長期,將處于指數生長期的微藻接種 300mL錐形瓶,實驗初始密度為5×104cell/mL。微藻接種前,預先向培養液中通各濃度的CO2氣體24h,使培養液中的碳酸鹽系統達到穩定。在培養24、48、72和96h后進行相關指標的測定。
1.3 葉綠素熒光參數的測定
采用調制葉綠素熒光儀(Water-PAM,Walz,德國)測定390和1000ppmv CO2條件下鹽生杜氏藻誘導光曲線和快速光響應曲線,來反映 CO2濃度升高對鹽生杜氏藻葉綠素熒光參數的影響。
測定誘導光曲線時,先將樣品暗適應 20min,使得光PSⅡ處于完全開放狀態。打開測量光,測定初始熒光值(F0),隨后打開飽和脈沖,持續時間為0.8s,測得Fm和Fv/Fm值。40s之后打開光化光,每隔 20s打開飽和脈沖,連續進行13次,直至最大熒光值(Fm′)達到穩定。所得熒光值按照公式(1)計算光系統Ⅱ的實際光化學效率(Yield):
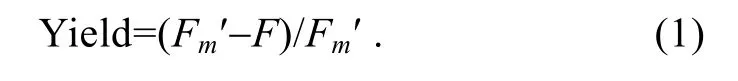
快速光響應曲線測定方法:打開測量光[0.01μmol/(m2·s)]測定初始熒光值F0,然后打開飽和脈沖[4000μmol/(m2·s),0.8s],測定Fm和Fv/Fm值。在預實驗的基礎上,光化光強度分別設定為0、90、150、210、265、295、350、405、480μmol/(m2·s),共 8 個梯度。在每個光化光梯度下,測得飽和脈沖前的熒光值(F)和打開飽和脈沖后測得的Fm′。相對電子傳遞速率(rETR)通過公式(2)計算:
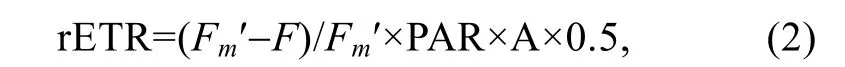
其中,PAR代表光化光強度;A表示藻類的吸光系數0.84;0.5代表有50%的吸收光能被分配到光系統Ⅱ。隨后,以光化光強度為X軸,rETR為Y軸制作相對電子傳遞速率隨 PAR變化的曲線,即快速光響應曲線(rapid light curve,RLC)。隨后,對快速光響應曲線進行最小二乘法擬合(Plattet al,1980),得到初始斜率(α)、最大相對電子傳遞速率(rETRmax)、半飽和光強(Ek)等參數:
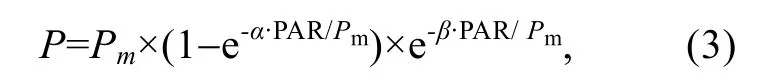
其中,P代表給定光化光強度下的相對電子傳遞速率(rETR);α代表快速光曲線的初始斜率,能夠反映光能的利用效率;Pm代表最大相對電子傳遞速率(rETRmax);Ek=Pm/α,代表半飽和光強,反映藻類對光強的耐受能力;β代表光抑制參數。
2 結果與分析
由圖1可知,隨著PAR強度的提升,鹽生杜氏藻的相對電子傳遞速率先線性上升到最大值后保持不變。24、48、72、96h時,390和1000ppmv CO2條件下鹽生杜氏藻快速光響應曲線擬合參數如表1所示:24h時,1000ppmv處理組與390ppmv處理組相比較,α值無顯著性差異(P>0.05);48、72、96h時,1000ppmv處理組的α值顯著增加(P<0.05),與390ppmv CO2條件下相比,分別增加12.5% (P<0.05)、21.7%(P<0.05)、8.333% (P<0.05)。
在前72小時內,390和1000ppmv CO2條件下鹽生杜氏藻光抑制參數β無顯著性差異(P>0.05);在96h時,1000ppmv CO2條件下生長的鹽生杜氏藻的β發生了顯著變化,比 390ppmv CO2條件下的值增加了45.45%。
對于rETRmax值來說,前72小時內,1000ppmv CO2條件下的rETRmax值均高于390ppmv CO2,但是無顯著差異(P>0.05);在96h時,390ppmv CO2條件下的的 rETRmax略高于對照組,但兩者間仍無顯著性差異(P>0.05)。
在24h時,390和1000ppmv CO2條件下鹽生杜氏藻的半飽和光強Ek無顯著性差異(P>0.05),但是通過比較48、72、96h的Ek值發現,其在1000ppmv CO2條件下均顯著降低(P<0.05),分別為390ppmv CO2條件下的90%、86.79%、87.10%。
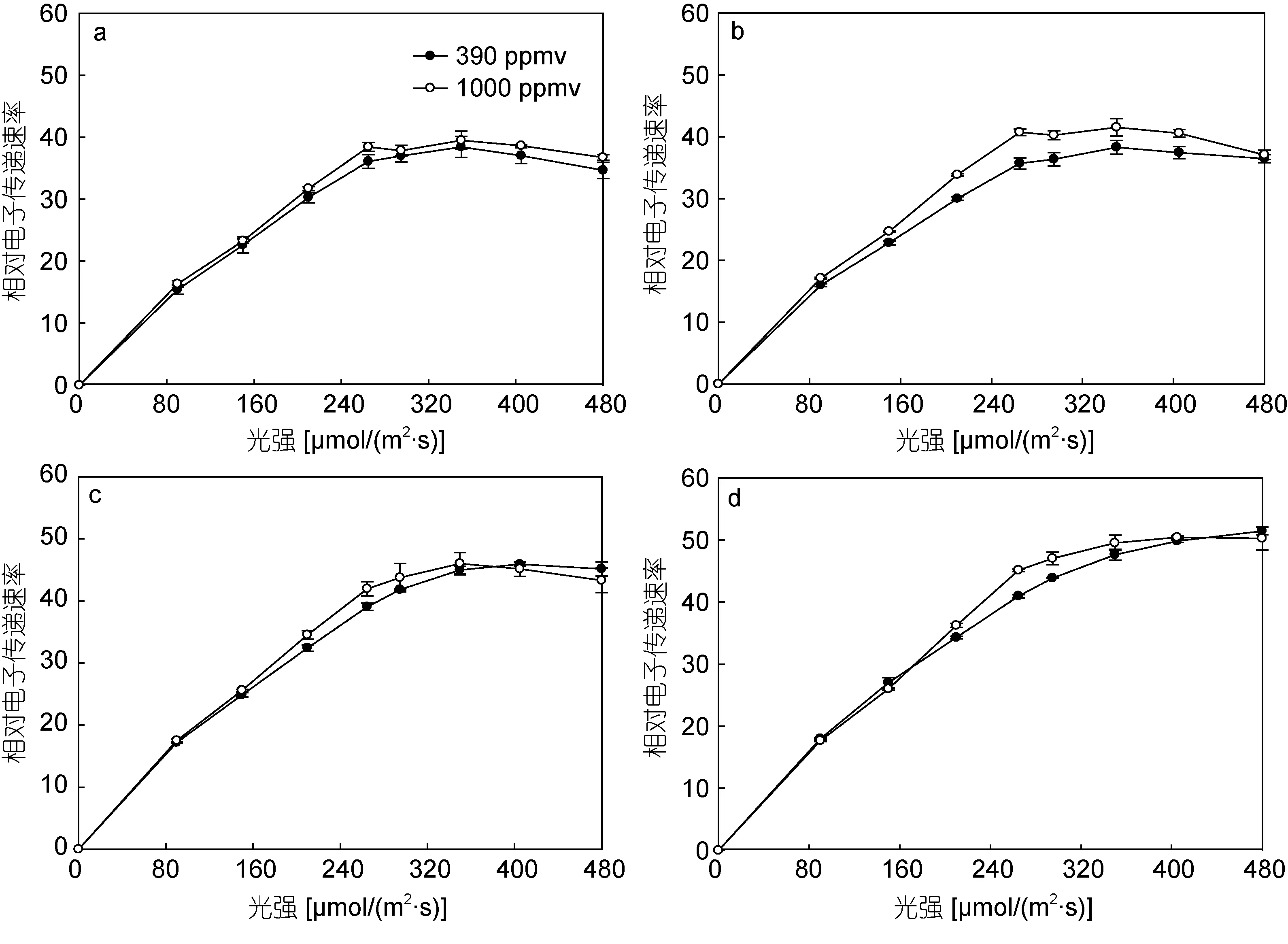
圖1 不同CO2濃度下鹽生杜氏藻快速光響應曲線Fig.1 The curves of rapid response to light by D.salina under different pCO2 in different exposure durations注:a:光照時間24h;b:48h;c:72h;d:96h
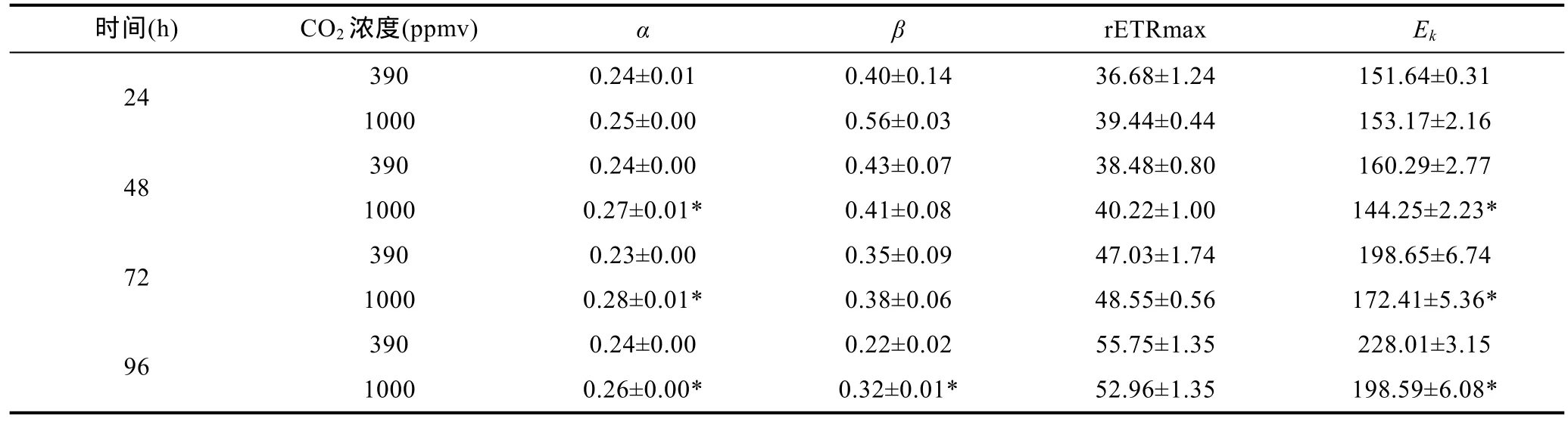
表1 24、48、72、96h時不同CO2濃度下鹽生杜氏藻快速光響應曲線擬合參數Tab.1 Parameter derived from curves of rapid response to light by D.salina under different pCO2 in 24h,48h,72h and 96h
以 PAR強度為橫坐標,以相應的非光化學淬滅值為縱坐標作圖(圖2),得到不同 CO2濃度下,鹽生杜氏藻非光化學淬滅值隨 PAR的變化規律。結果表明,隨著PAR強度的上升,所有實驗組中的NPQ值均逐漸增大。通過對比24、48、72、96h時,390和1000ppmv CO2條件下鹽生杜氏藻非光化學淬滅值發現,24、48、72h 內,當 PAR 大于 150 μmol/(m2·s)μmol/(m2·s)時,1000ppmv CO2條件下的 NPQ 值均大于390ppmv CO2條件下;96h時,在PAR較低時[0—150μmol/(m2·s)],390ppmv CO2條件下的 NPQ 顯著高于1000ppmv CO2(P<0.05),隨著PAR的上升,390ppmv CO2條件下 NPQ值增長速度逐漸低于1000ppmv CO2條件下,當PAR值大于295μmol/(m2·s)時,1000ppmv CO2條件下 NPQ顯著高于 390ppmv CO2條件下NPQ值。
24、48、72、96h 時,390 和1000ppmv CO2條件下鹽生杜氏藻誘導光曲線中 PSⅡ的潛在Fv/Fm和PSⅡ的實際光合效率如圖3和圖4所示。結果表明,在24、48、72、96h時,390和1000ppmv CO2條件下鹽生杜氏藻 PSⅡ的潛在Fv/Fm均無顯著差異。然而,在24、48、96h時,與對照組相比,1000ppmv CO2條件下鹽生杜氏藻的 PSⅡ的實際光合效率分別被促進了 5.8% (P<0.05)、6.9% (P<0.05)和3.8% (P<0.05)。
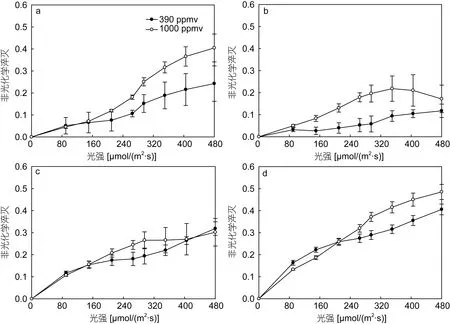
圖2 不同CO2濃度下不同光照時間鹽生杜氏藻非光化學淬滅Fig.2 NPQ (non-photochemical quenching)of D.salina under different pCO2 in different light exposure durations
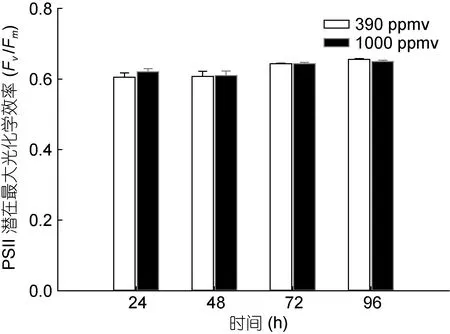
圖3 不同CO2濃度下不同光照時間對鹽生杜氏藻PSⅡ的潛在最大光合作用效率(Fv/Fm)的關系Fig.3 Fv/Fm of D.salina under different pCO2 in different lightexposure length
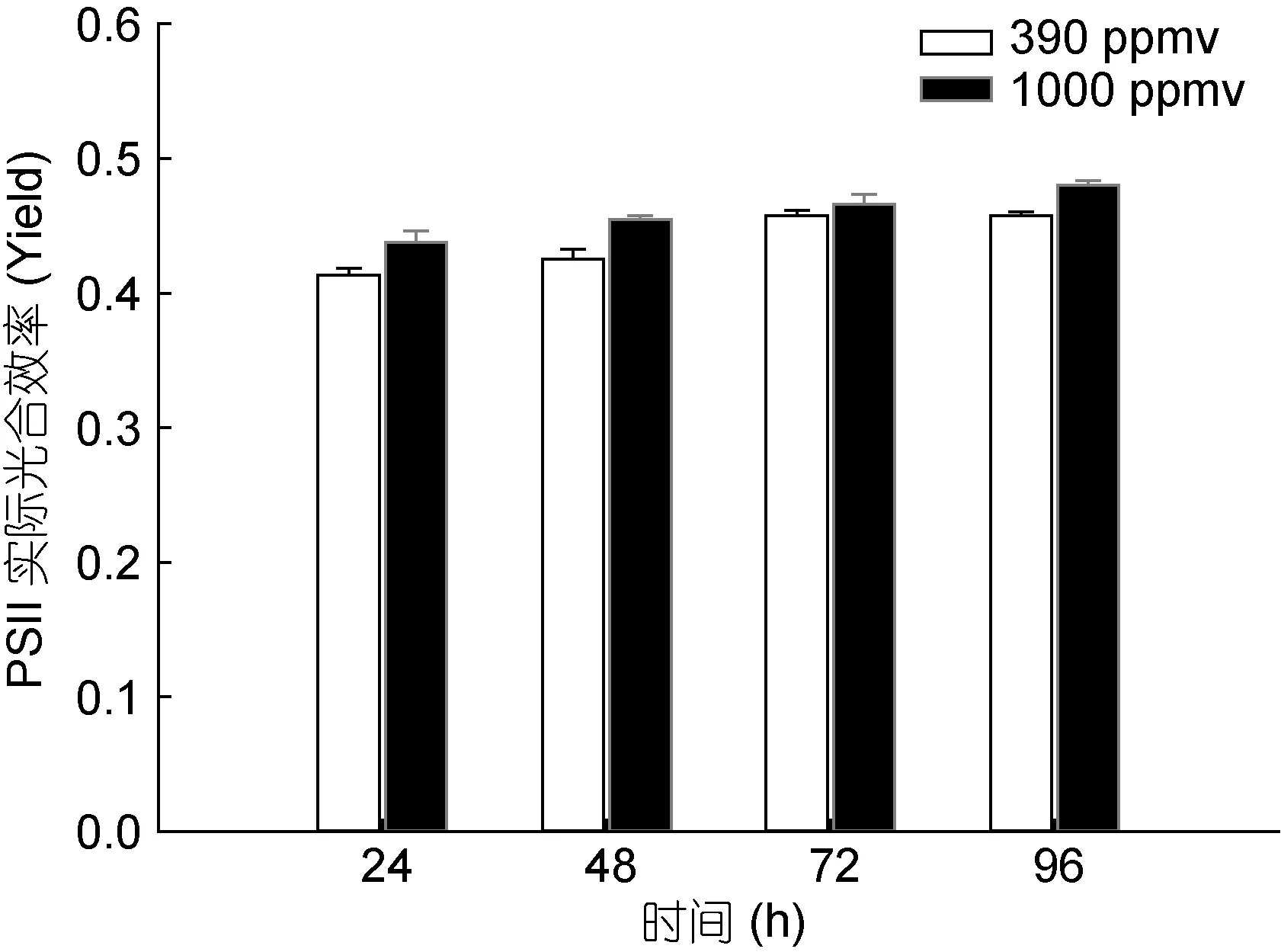
圖4 24、48、72、96h時不同CO2濃度下鹽生杜氏藻PSⅡ的實際光合效率Fig.4 Yield of D.salina under different pCO2 at 24h,48h,72h,96h.
3 討論
當外界環境發生變化時,葉綠素熒光是反映環境變化對光合生物影響的重要指標之一(Maxwelletal,2000),能夠客觀地反映光合生物本身的光合特征及其對環境變化的適應能力(Jianget al,2003;李鵬民等,2005)。通過葉綠素熒光技術測定光合生物的快速光響應曲線和誘導曲線,不但可以反映細胞在實驗條件下對光能的分配和響應能力,同時也可以反映細胞在不同光合環境條件下的潛在光合特征(Franket al,1996)。本研究結果表明,CO2濃度升高并沒有改變鹽生杜氏藻 PSⅡ的Fv/Fm和rETRmax,這與徐金濤等(2016)對塔瑪亞歷山大藻的研究結果相反,當CO2濃度由370ppmv增加至1000ppmv時,塔瑪亞歷山大藻的Fv/Fm值和rETRmax顯著增加。盡管 CO2升高并沒有改變鹽生杜氏藻的Fv/Fm和rETRmax,但是從光能利用效率、半飽和光強以及 PSⅡ的實際光化學量子產額的角度上來講,CO2濃度升高對鹽生杜氏藻的 PSⅡ產生了積極的促進作用。通常來說,光能利用效率(α)反映了光合生物對光能的吸收和利用效率(Fuet al,2007,2008)。對鹽生杜氏藻來說,高濃度 CO2條件下鹽生杜氏藻的光能利用效率明顯高于對照組,這可能是由于對照組中,更多的能量被用來濃縮CO2以滿足核酮糖-1,5二磷酸羧化酶的需要,因此在吸收光能不變的情況下,高濃度 CO2下更多的能量被用于光合固碳,從而提高了光能利用效率(Olischl?geret al,2013)。 類 似 的 結 果在Synechococcussp.和Ulva prolifera中也同樣被發現(Fuet al,2007;Liuet al,2012)。Olischl?ger等(2013)發現,在10°C條件下,Neosiphonia harveyi的光能利用效率在1411ppmv CO2條件下被顯著促進,但是當溫度升高至17.5°C時,CO2升高對其光能利用效率無顯著影響,這說明其他環境因子(如溫度)在一定程度下會影響光能利用效率對CO2升高的響應。
半飽和光強Ek代表光合生物保持捕獲光能和處理光能的最佳平衡點(Olischl?geret al,2013)。本實驗中,在高濃度CO2條件下,光能利用效率的升高和未發生顯著變化的最大相對電子傳遞速率引起鹽生杜氏藻Ek的降低。CO2濃度升高導致的Ek降低并非普遍存在于所有浮游植物類群中,Suárez-álvarez 等(2012)的實驗表明,Hypnea spinella的Ek在高濃度CO2條件下顯著增加,這可能是CO2升高使得最大相對電子傳遞速率被顯著促進所導致的;另外,也有研究表明,CO2升高并不改變浮游植物的Ek(Wuet al,2010;Yanget al,2012)。從這個角度上講,未來在海洋酸化條件下,鹽生杜氏藻對光照強度的依賴程度將會減弱,大大降低了在自然環境中遭受光強限制的可能性。與酸化情況下Ek不變或升高的浮游植物類群相比,鹽生杜氏藻在種群競爭和群落演替過程中將比現在更具競爭優勢。
不斷加劇的海洋酸化將會對海洋中的鈣化生物產生嚴重的威脅,主要表現為降低鈣化生物的鈣化速率、表面鈣片脫落等(Gaoet al,1993;Riebesellet al,2000)。對海洋中的非鈣化生物來說,雖然 CO2升高導致的海洋酸化促進了非鈣化生物的生長、光合和初級生產力,但是酸化對非鈣化生物同樣也是一個潛在的脅迫因子。本研究發現,在1000ppmv CO2條件下,鹽生杜氏藻光系統Ⅱ的光抑制率(β)顯著增加,并且伴隨著較高的NPQ值。Tchernov等(2001)指出,浮游植物可以通過無機碳濃縮機制(CCMs)的運行耗散多余的光能、分流電子以起到緩解光抑制的作用。因此,鹽生杜氏藻光抑制程度增加可能是由于高濃度CO2下調了CCMs的運行所致。在光限制條件下,下調 CCMs所節省的能量可能會促進浮游植物的生長和光合作用;但是在光充足的情況下,這部分能量不但不會補充藻細胞對光能的需求,反而增加了過剩光能引起的光抑制程度,類似的結果在三角褐指藻和滸苔幼苗中也同樣被發現(Wuet al,2010;Liuet al,2012)。
4 結論
基于本實驗的結果,CO2導致的海洋酸化將會對鹽生杜氏藻光系統Ⅱ產生促進作用,主要表現為增加的光能利用效率和PSⅡ的實際光化學量子產額以及降低的半飽和光強;但是在光能充足的條件下,CO2升高導致的海洋酸化增加了鹽生杜氏藻的光抑制程度,并且導致了更高的 NPQ值。海洋酸化對鹽生杜氏藻的正面效應和負面效應的凈值將決定其最終對海洋酸化的響應。比較之前的研究可以發現,浮游植物對海洋酸化的響應存在種間差異,而這種種間差異很可能會改變浮游植物的種間競爭關系,甚至會影響浮游植物類群的進化。
李鵬民,高輝遠,Strasser R J,2005.快速葉綠素熒光誘導動力學分析在光合作用研究中的應用.植物生理與分子生物學學報,31(6):559—566
陳斌斌,2015.海洋酸化背景下經濟海藻龍須菜與壇紫菜的生物學特性.廣州:華南理工大學博士學位論文
徐金濤,龐 敏,馬 新等,2016.CO2加富對塔瑪亞歷山大藻葉綠素熒光參數及產毒的影響.海洋與湖沼,47(3):557—563
Caldeira K,Wickett M E,2003.Oceanography:anthropogenic carbon and ocean pH.Nature,425(6956):365
Doney S C,Fabry V J,Feely R Aet al,2009.Ocean acidification:the other CO2problem.Annual Review of Marine Science,1:169—192
Fabry V J,Seibel B A,Feely R Aet al,2008.Impacts of ocean acidification on marine fauna and ecosystem processes.ICES Journal of Marine Science,65(3):414—432
Frank H A,Cogdell R J,1996.Carotenoids in photosynthesis.Photochemistry and Photobiology,63(3):257—264
Fu F X,Warner M E,Zhang Yet al,2007.Effects of increased temperature and CO2on photosynthesis,growth,and elemental ratios in marineSynechococcusandProchlorococcus(cyanobacteria).Journal of Phycology,43(3):485—496
Fu F X,Zhang Y,Warner M Eet al,2008.A comparison of future increased CO2and temperature effects on sympatricHeterosigma akashiwoandProrocentrum minimum.Harmful Algae,7(1):76—90
Gao K,Aruga Y,Asada Ket al,1993.Calcification in the articulated coralline algaCorallina pilulifera,with special reference to the effect of elevated CO2concentration.Marine Biology,117(1):129—132
Hughes L,2000.Biological consequences of global warming:is the signal already apparent? Trends in Ecology &Evolution,15(2):56—61
Jiang C D,Gao H Y,Zou Q,2003.Changes of donor and acceptor side in photosystem 2 complex induced by iron deficiency in attached soybean and maize leaves.Photosynthetica,41(2):267—271
Liu Y T,Xu J T,Gao K,2012.CO2-driven seawater acidification increases photochemical stress in a green alga.Phycologia,51(5):562—566
Maxwell K,Johnson G N,2000.Chlorophyll fluorescence-a practical guide.Journal of Experimental Botany,51(345):659—668
Olischl?ger M,Wiencke C,2013.Ocean acidification alleviates low-temperature effects on growth and photosynthesis of the red algaNeosiphonia harveyi(Rhodophyta).Journal of Experimental Botany,64(18):5587—5597
Platt T,Gallegos C L,Harrison W G,1980.Photoinhibition of photosynthesis in natural assemblages of marine phytoplankton.J Mar Res,38:687—701.
Riebesell U,Zondervan I,Rost B è et al,2000.Reduced calcification of marine plankton in response to increased atmospheric CO2.Nature,407(6802):364-367.
Suárez-álvarez S,Gómez-Pinchetti J L,García-Reina G,2012.Effects of increased CO2levels on growth,photosynthesis,ammonium uptake and cell composition in the macroalga Hypnea spinella (Gigartinales,Rhodophyta).Journal of Applied Phycology,24(4):815—823
Tchernov D,Helman Y,Keren Net al,2001.Passive entry of CO2and its energy-dependent intracellular conversion to HCO3–in cyanobacteria are driven by a photosystem I-generated ΔμH+.Journal of Biological Chemistry,276(26):23450—23455
Wu Y,Gao K,Riebesell U,2010.CO2-induced seawater acidification affects physiological performance of the marine diatomPhaeodactylum tricornutum.Biogeosciences,7(9):2915—2923
Yang G Y,Gao K S,2012.Physiological responses of the marine diatomThalassiosira pseudonanato increasedpCO2and seawater acidity.Marine Environmental Research,79:142—151
EFFECTS OF ELEVATED CO2ON CHLOROPHYLL FLUORESCENCE TODUNALIELLA SALINA
ZANG Yu1, HUANG Zhi-Yuan3, ZHAO Xin-Yu1, HU Shun-Xin1,2
(1.College of Marine Life Science,Ocean University of China,Qingdao266003,China;2.Shandong Marine Resource and Environment Research Institute,Yantai264006,China;3.Qingdao No.58High School of Shandong Province,Qingdao266199,China)
The increasing atmosphere carbon dioxide have led to a reduction in pH in the ocean and caused a wide range of environment,ecology,and climate problems.To investigate the potential effect of elevated CO2on marine microalgae photosynthesis,Dunaliella salinawas exposed to two differentpCO2levels:390 and 1000 ppmv.Results show that the elevated CO2level stimulated the light utilization efficiency (α),actual photochemical efficiency (Yield),non-photochemical quenching (NPQ),and photoinhibition rate (β),decreased the light saturation point (Ek)ofD.salina,while rETRmaxandFv/Fmremained stable.
CO2;ocean acidification;Dunaliella salina;the chlorophyll fluorescence
Q14
10.11693/hyhz20170300051
* 海洋公益性行業科研專項經費資助項目,201305027號;國家自然科學基金項目,41476091號。臧 宇,E-mail:493687012@qq.com
① 通訊作者:胡順鑫,E-mail:290166784@qq.com
2017-03-12,收修改稿日期:2017-06-14
