高溫干旱對髯毛箬竹的葉和細根的生理生態影響
2017-12-16 05:44:17李偉成盛海燕高貴賓田新立
環境科學研究 2017年12期
李偉成, 盛海燕, 高貴賓, 溫 星, 田新立*
1.國家林業局竹子研究開發中心, 浙江省竹子高效加工重點實驗室, 浙江 杭州 310012 2.西南林業大學生命科學學院, 云南 昆明 650224 3.杭州環境保護科學研究院, 浙江 杭州 310005
高溫干旱對髯毛箬竹的葉和細根的生理生態影響
李偉成1,2, 盛海燕3, 高貴賓1, 溫 星1, 田新立1*
1.國家林業局竹子研究開發中心, 浙江省竹子高效加工重點實驗室, 浙江 杭州 310012 2.西南林業大學生命科學學院, 云南 昆明 650224 3.杭州環境保護科學研究院, 浙江 杭州 310005
為了解全球變暖及其導致的高溫干旱氣候對地被竹生長產生的影響,通過模擬自然高溫干旱條件,以髯毛箬竹為研究對象,分析其葉片氣體交換、細根和葉片水勢、抗氧化酶等生理生態反應過程與高溫干旱的關系. 結果表明:①不同高溫干旱條件對髯毛箬竹的影響不同,對照組中常溫條件下植株的光合速率為(7.141±0.072) μmol(m2·s),高溫可明顯提升蒸騰速率和氣孔導度,極端高溫條件下表現為非氣孔限制因素導致光合速率〔(4.898±0.395) μmol(m2·s)下降〕,并且收窄光合日進程的變幅;②中 度干旱、高溫+中度干旱、極端高溫+中度干旱的條件下,光合速率的降低是由氣孔限制因素導致;③重度干旱、高溫+重度干旱、極端高溫+重度干旱的環境將嚴重影響植株生理進程,高溫加劇這一進程,具體表現為蒸騰速率、光合速率、氣孔導度和水分利用效率顯著下降;④對照組、高溫組和極端高溫組的細根水勢谷值下降范圍分別為(-0.111±0.033)~(-0.961±0.086)、(-0.173±0.060)~(-0.970±0.072)和(-0.304±0.061)~(-1.225±0.166) MPa,降幅比成熟葉片強烈;⑤高溫和干旱對細根、成熟葉片3種抗氧化酶活性的影響一致,細根SOD活性大于成熟葉片,而CAT和POD活性則相反,因此,細根是髯毛箬竹適應高溫干旱環境的重要器官,水分吸收器官比蒸騰器官敏感. 研究顯示,髯毛箬竹具有較好的抗高溫和抗旱能力,一定范圍內的高溫和干旱能提升其葉片、細根的生理生態適應能力,特別是復合條件下,高溫與干旱對其生長表現出協同作用,極端高溫+中度干旱并不威脅其生存,高溫、中度干旱及高溫+中度干旱有利于馴化栽培,甚至在一定程度上能促進其生長;重度干旱則使植株葉片氣體交換、葉片和細根的水分生理進程受損,不適宜長期重度干旱的環境,高溫使這種損傷更加嚴重.
復合條件; 地被竹; 氣體交換; 水勢; 抗氧化酶
竹類植物有較高園林美學價值,地被竹是竹類植物中一個新穎且富有特色的重要類群,是園林景觀和城市林業建設的新寵.目前對地被竹的光合、蒸騰和氣孔特性僅限于單一環境因子的研究[6-7]. 李娟等[8]對地被竹——黃條金剛竹的研究表明,在單因素干旱脅迫25 d后植株的光合機構被破壞,葉綠素含量下降,丙二醛含量上升,最大光化學效率(FvFm)和PSⅡ 潛在熱耗散能力下降,氣孔是光合參數變化的主導因素. Beebe等[9]認為,隨著干旱脅迫程度和時間增加,植物凋落量增加,同時高溫延長植物生長期,但二者并不是簡單的疊加作用;XU等[10]的試驗結果表明,高溫干旱復合脅迫條件下促進刺槐幼苗的光合、蒸騰速率和氣孔導度,而單一條件下,生理指標上升并不明顯. 可見,高溫干旱復合脅迫對不同植物的影響不同. 而植物作為一個整體,在應對復雜的氣候環境變化時,地上與地下器官對不同環境因子交互作用的協同響應的方式如何?因此,同時考慮影響植物生長、發育和功能的溫度和水分因子,才能正確推斷未來氣候條件下植物的生理生態過程. 國內外對高溫或干旱單一因素的脅迫研究有大量報道[11-12],但二者復合脅迫的研究較少[9,13].
髯毛箬竹(IndocalamusbarbatusMcClure),葉大色綠,繁殖能力較強,地被應用景觀價值高,適合于我國南方中低海拔地區進行大面積栽培和造景. 該研究通過模擬自然高溫干旱環境,進行髯毛箬竹的溫室盆栽試驗,以其細根和成熟葉片為對象,研究高溫干旱脅迫與髯毛箬竹相關生理生態反應過程的關系,分析其高溫干旱耐受能力和恢復能力,為髯毛箬竹在園林物種配置和造景維護方面提供數據支撐,為篩選適合園林美化與道路綠化的地被竹種進行馴化栽培提供理論依據.
1 材料與方法
1.1 試驗地概況與試驗材料
試驗地位于浙江省湖州市長興縣小浦鎮葡萄岕苗圃地(119°45′51.044 8″E、30°58′59.198 1″N),屬亞熱帶海洋性季風氣候. 年均氣溫(15.6±0.5)℃,有效積溫 5 750 ℃,無霜期239 d. 年均降水量 1 309 mm. 年均日照時數 1 810.3 h,歷年平均日照百分率為41%,光照分配較均勻. 土壤以發育于酸性巖漿巖和沉積巖的紅壤為主. 試驗地土壤層深厚,土壤層可達1.5 m. 林地土壤有機質平均含量為30.2 gkg,林地土壤中w(TN)為1.16~2.24 gkg,w(有效磷)為98.00~101.76 mgkg,w(速效鉀)為54.75~92.61 mgkg.
2015年10月上旬在白色塑料薄膜搭建的溫室大棚中,選取健康無病蟲害的兩年生髯毛箬竹進行試驗. 將苗高35~40 cm的長髯箬竹植株,僅留1分枝附3~5片葉,帶5~10 cm的根鞭,移栽入長×寬×高為3.0 m×1.2 m×0.20 m苗床(四周用客土砌成擋墻高30 cm,以保證透氣),株距20~30 cm,行距10~15 cm;植苗后,采用滲灌供水技術[14],PVC滲灌管平鋪于苗床,管的間距為10~15 cm,在10~40 cm水頭壓力下滲水速率為(22.09±4.77)cm3(m·min). 滲灌至飽和,土壤表層先覆蓋2~3 cm厚的河沙,再覆蓋1~2 cm 厚的腐殖土,以保持苗床內水分. 期間常規管理與施水,試驗苗栽培130 d. 放置大棚從地面至40 cm高處不覆蓋薄膜,起到通風的作用,頂棚封膜,晴天開啟頂棚,雨天關閉頂棚薄膜. 土壤本底養分:w(有機質)為(20.08±3.61)gkg,w(TN)為(2.64±0.55)gkg,w(有效磷)為(15.29±3.88)mgkg,w(速效鉀)為(40.50±8.37)mgkg,pH為5.1±0.3.
1.2 試驗設計
于2016年3月20日開始模擬自然高溫干旱失水與復水過程,設置對照組(對照、中度干旱和重度干旱)、高溫組(高溫、高溫+中度干旱和高溫+重度干旱)和極端高溫組(極端高溫、極端高溫+中度干旱、極端高溫+重度干旱),共9個處理. 溫度脅迫的實施方案:在各處理的苗圃地上方離地面高1.6 m處架設硅碳發熱管(黑管,直徑1.8 cm,長1.0 m,4根串聯成一行,每行相隔0.5 m,并聯成3行排列),在距離地面1.0 m處的中間位置懸設溫度計以方便數控溫度[15]. 各處理組的晝夜溫度控制見表1. 干旱脅迫的土壤水勢設為-30~0 kPa(對照,土壤水分含量為37.86%±5.95%)、-60~-30 kPa(中度干旱,土壤水分含量為27.95%±2.64%)和-90~-60 kPa(重度干旱,土壤水分含量為18.25%±1.73%)3個處理,土壤水勢由土壤水分張力計(內置傳感器)監測并采集數據,重度干旱及其復合處理以每日中午試驗用苗不發生萎蔫為標準.
1.3 試驗測量與數據處理
1.3.1蒸騰速率、氣孔導度和凈光合速率測定
3.企業財務管理能準確地了解企業的資金運用情況,從而保證企業的良好發展。企業的財務管理的水平直接決定著企業能否健康穩定的發展。因此,企業需要將財務管理貫穿在日常的經營中,保證財務管理的科學合理,從而有效地分配企業的資產。此外,企業的財務管理可以保證日常經營活動順利進行,強化企業生產經營過程中的資金運行情況以及資產分配情況,保證企業的日常運行。
2016年5月1日,在模擬試驗進行42 d后,在各自試驗狀態下,選取每株當年生苗最長枝條從上往下數的第3片完整展開的成熟葉片,選擇晴朗天氣的04:00—20:00每隔1 h采用Li6400(LI-COR,USA)測定葉片蒸騰速率(Tr)、凈光合速率(Pn)和氣孔導度(Gs)指標,每處理重復5株;由于試驗中04:00—06:00和18:00以后凈光合速率都為0 μmol(m2·s),故僅取06:00—18:00的凈光合速率數據進行分析;同時,根據蒸騰速率與光合速率的比值計算葉片水分利用效率(WUE)[4].
1.3.2葉片與細根的水勢測定
葉片氣體交換測定的同時,選取同一處理的另一部分試驗栽培苗,在04:00—20:00每隔2 h測定細根(≤2 mm)水勢和成熟葉片水勢動態. 采用WP4-T露點水勢儀(Decagon,USA)測定成熟葉片水勢,采用壓力室法測定細根水勢,各處理每次測定的重復株數為5株.
1.3.3抗氧化酶測定
在試驗結束后立即取樣,在17:00—18:00采集髯毛箬竹的中部葉片進行抗氧化酶活性測定,共采樣5次,取平均值. 測定SOD(超氧化物歧化酶)、CAT(過氧化氫酶)和POD(過氧化物酶)的活性. 取0.2 g葉片洗凈后置于研缽中,加入1.6 mL 50 mmolL預冷的磷酸緩沖液(pH為7.8)在冰浴上研磨成勻漿,轉入離心管中在 4 ℃、12 000 rmin 下離心20 min,取上清液測定酶活性,其中SOD活性采用氮藍四唑法測定,CAT活性采用紫外吸收法測定,POD活性采用愈創木酚比色法測定[16].
1.3.4數據處理
使用SPSS 11.0進行數據處理,采用單因素方差分析(one-way ANOVA)和最小顯著差異法(LSD)比較不同處理間的差異顯著性.
2 結果與分析
2.1 不同處理條件下髯毛箬竹葉片的蒸騰速率、光合速率、氣孔導度及水分利用效率
在高溫、干旱及高溫干旱復合條件下,髯毛箬竹蒸騰作用、光合作用和氣孔導度的日進程見圖1. 在對照組(常溫)、高溫組(33~36 ℃)和極端高溫組(37~40 ℃)中,蒸騰速率、光合速率和氣孔導度三者都表現為對照>中度干旱>重度干旱、高溫>高溫+中度干旱>高溫+重度干旱、極端高溫>極端高溫+中度干旱>極端高溫+重度干旱,說明各處理組的土壤水分條件對髯毛箬竹的蒸騰、光合速率和氣孔導度有顯著影響. 高溫+重度干旱和極端高溫+重度干旱兩種復合處理的蒸騰、光合速率和氣孔導度日進程都呈現“單峰”,而且重度干旱處理下的氣孔導度也出現“單峰”狀態.
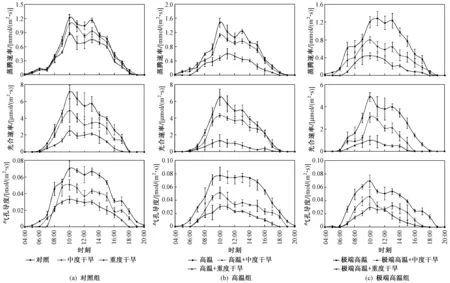
圖1 不同處理組中髯毛箬竹葉片的蒸騰速率、光合速率及氣孔導度日進程Fig.1 Diurnal processes of transpiration rate, photosynthesis rate and stomatal conductance of I. barbatus′s leaves under different treatment groups
與重度干旱處理相比較,高溫+重度干旱和極端高溫+重度干旱兩種復合處理的植株蒸騰速率峰值顯著下降(P<0.05)(見表2),僅為(0.606±0.148)和(0.422±0.058) mmol(m2·s),而其他處理都在10:00和13:00左右出現峰值,呈現“雙峰”態勢,說明這兩種處理條件下,蒸騰作用被顯著抑制. 高溫處理的蒸騰速率峰值為(1.480±0.129) mmol(m2·s),與對照有顯著差異(P<0.05),說明土壤水分供應充足的高溫條件可促進髯毛箬竹植株的蒸騰作用. 極端高溫條件下,其峰值的平均值較對照處理高,但差異不顯著(P>0.05).
與對照組比較,極端高溫組各處理的光合速率峰值對應地顯著下降(P<0.05),而高溫組處理則不顯著(P>0.05)(見表2),重度干旱與極端高溫+重度干旱處理的光合速率峰值差異顯著(P<0.05),說明高溫對髯毛箬竹的影響不明顯. 而中度干旱與高溫+中度干旱光合速率峰值差異不顯著(P>0.05),中度干旱與極端高溫+中度干旱、高溫+中度干旱與極端高溫+中度干旱處理光合速率峰值都表現為差異顯著(P<0.05),說明在極端高溫條件下,溫度對髯毛箬竹的光合速率有明顯影響,而高溫+中度干旱條件下,溫度對光合速率的峰值負面影響不顯著,且對整體日進程有積極影響. 日進程中,重度干旱、高溫+重度干旱、極端高溫+重度干旱處理的植株在15:00左右的光合速率已經基本降為0,說明重度干旱是髯毛箬竹光合作用的主要影響因素.
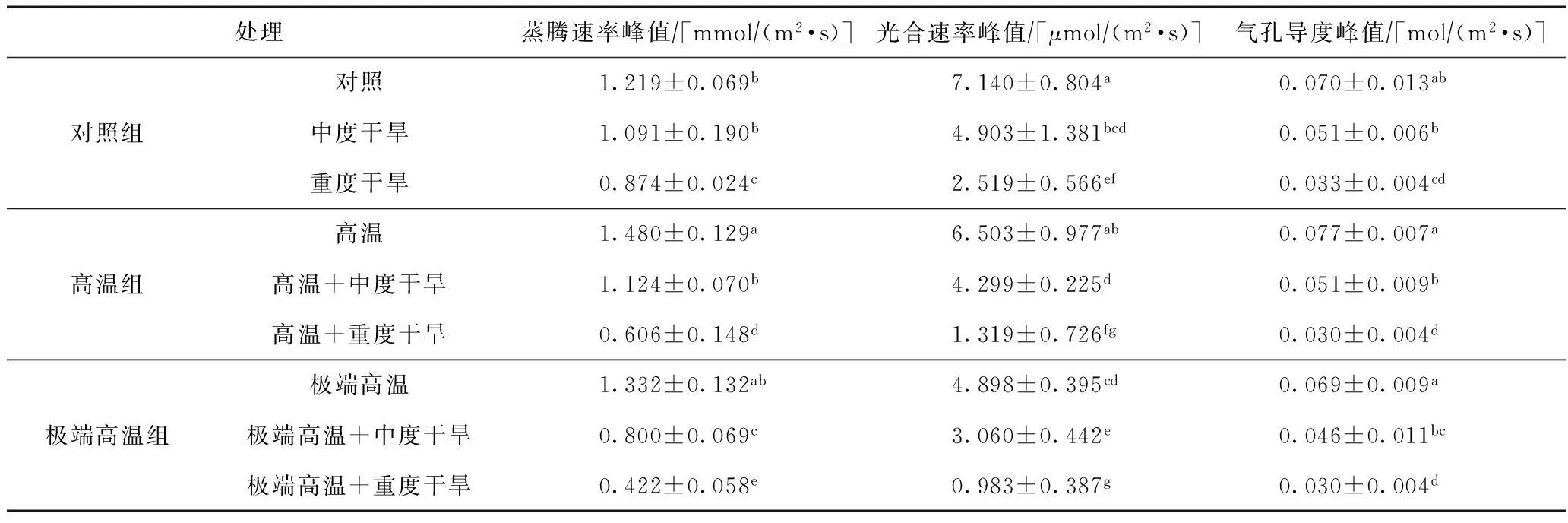
表2 不同處理組中髯毛箬竹葉片的蒸騰速率、光合速率及氣孔導度峰值
注: 數據為平均值±標準差;不同小寫字母表示差異顯著(P<0.05).
與對照組相比較,高溫組和極端高溫組的氣孔導度峰值差異不顯著(P>0.05)(見表2),且高溫處理日進程每個時間點測得數據的均值都有一定提升,而極端高溫處理的均值則有所下降,說明高溫提高了髯毛箬竹葉片氣孔的開放程度,而極端高溫處理對氣孔導度有一定的限制,但不至于對生理過程形成危害. 重度干旱、高溫+重度干旱、極端高溫+重度干旱3個處理的氣孔導度峰值差異不顯著(P>0.05),但相對于常溫對照、高溫和極端高溫處理而言,氣孔導度急劇下降,這與蒸騰速率和光合速率的情況類似.
由各處理的葉片水分利用效率日進程(見圖2)可知,植株葉片在07:00—08:00的水分利用效率較高,然后波動下降;高溫+中度干旱處理的葉片水分利用效率日進程具有較高的平均值,變化幅度較小,大部分數據與高溫處理的日進程有顯著差異(P<0.05),且與12:00以后中度干旱處理的水分利用效率亦有顯著差異(P<0.05),說明該處理的葉片水分利用效率較高. 重度干旱、高溫+重度干旱、極端高溫+重度干旱3個處理的葉片水分利用效率日進程與各自組內其他處理有顯著差異(P<0.05). 高溫組中,葉片水分利用效率日進程的均值表明對照>中度干旱>重度干旱. 而極端高溫組中,極端高溫、極端高溫+中度 干旱處理之間差異不顯著(P>0.05),這與對照組的情況類似.
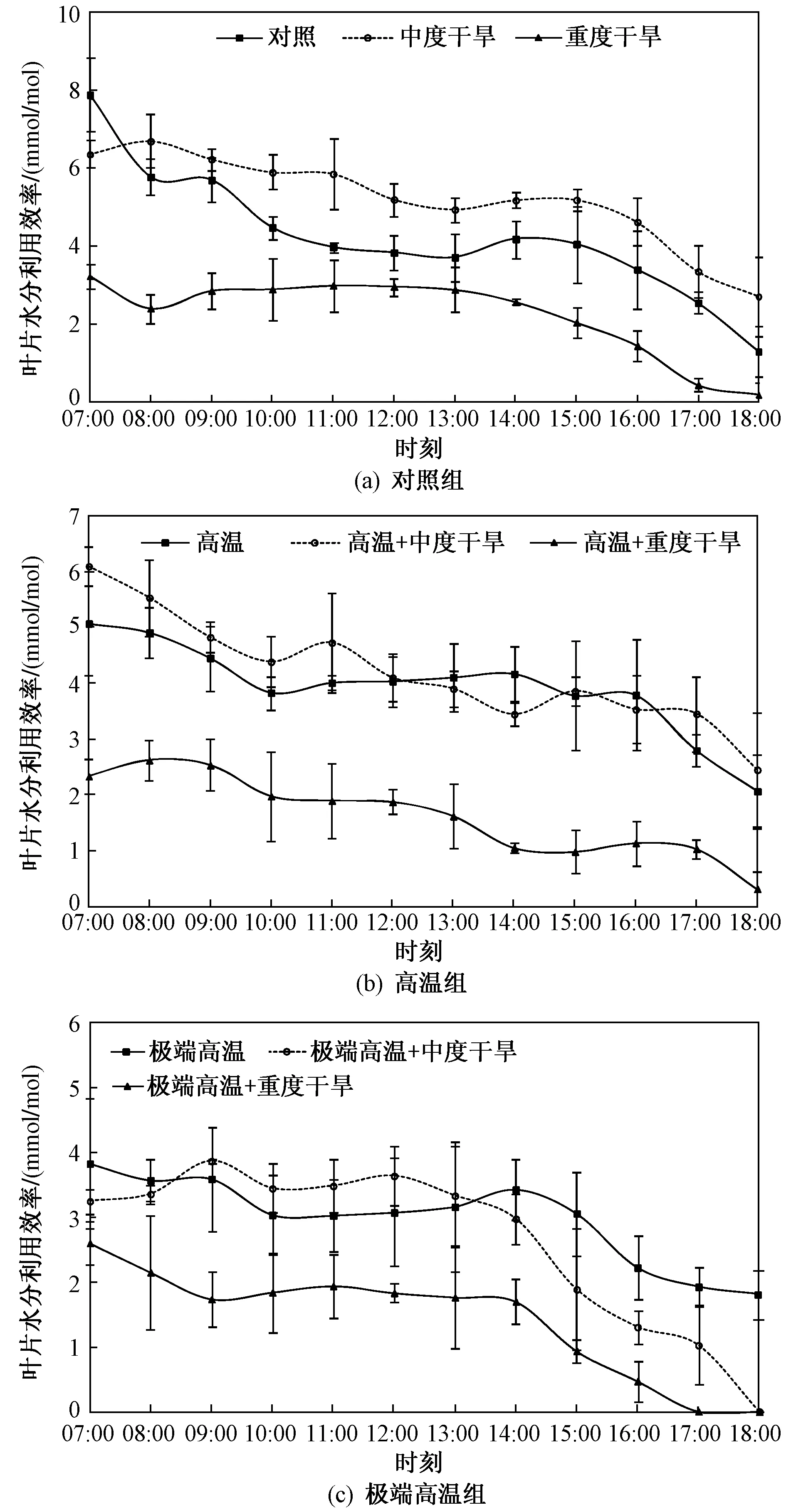
圖2 不同處理組組中髯毛箬竹葉片的水分利用率日進程Fig.2 Diurnal processes of water use efficiency of I. barbatus′s leaves under different treatment groups
2.2 不同處理條件下髯毛箬竹細根與成熟葉片的水勢動態變化
在蒸騰作用的拉力影響下,細根水勢的日動態模式見表3,其水勢變化表現為不同深淺的谷型. 對照、高溫和極端高溫這3種土壤水勢較高的處理下,細根水勢在一天中總是處于較高的動態水平,其日變幅不大,平均細根水勢分別為-0.010~-0.111、-0.011~-0.173 和-0.016~-0.304 MPa. 細根水勢在08:00開始下降,14:00達到谷值,對照和高溫處理細根水分虧缺在午后基本可以恢復,其在08:00—18:00的谷形亦不明顯,極端高溫處理與前二者在08:00—18:00有顯著差異(P<0.05),說明極端高溫增強了蒸騰作用,降低了細根水勢;隨著各處理組內的干旱程度的增加(土壤水勢的下降),細根水勢的動態水平亦下降,且差異顯著(P<0.05),隨著降幅的快速增大,形成深谷,下午細根水分虧缺的恢復過程明顯延遲,如對照和高溫處理的細根水勢平均值的日變動降幅相對其他各處理而言并不明顯,在16:00左右基本得到恢復;而中度干旱、高溫+中度干旱和極端高溫+中度干旱處理的細根水勢的日變動區間分別為-0.014~-0.604、-0.015~-0.723和-0.041~-1.028 MPa,降幅分別是對照處理的5.80、6.97和9.73倍,中度干旱與極端高溫+中度干旱處理之間差異顯著(P<0.05),20:00時僅中度干旱處理接近恢復到04:00的水平,說明隨著溫度的升高,加劇了蒸騰作用,上述涉及中度干旱的3個處理的細根水勢對溫度有明顯的響應;重度干旱、高溫+重度干旱和極端高溫+重度干旱這3個涉及重度干旱的處理,細根水勢的日變動降幅分別是對照處理的9.22、9.93和9.86倍,3個處理之間差異不顯著(P>0.05),20:00時3個處理的植株細根水分虧缺都沒有得到恢復,說明重度干旱已嚴重影響髯毛箬竹的日常生理進程.
由表3可知,在較高土壤水勢支持下,髯毛箬竹成熟葉片水勢始終維持相對較高的動態水平,且葉片水分虧缺后能得到有效恢復;而土壤低水勢則導致葉片水勢的相對較低水平波動,在葉片虧缺后不能得到迅速恢復. 對照組中,3個處理的葉片水勢日變幅分別為-0.057~-2.053、-0.060~-2.227和-0.123~-2.677 MPa,其日變幅與高溫組各處理接近,對照與中度干旱處理的差異不顯著(P>0.05),而對照、中度干旱處理分別與重度干旱處理的日變幅有顯著差異(P<0.05),高溫組和極端高溫組的成熟葉片水勢日變幅情況與此類似(僅14:00數據差異不顯著,P>0.05),其中極端高溫+重度干旱處理的谷值在各處理中最低,為(-2.967±-0.244)MPa. 對照組中,對照與中度干旱處理的日變化過程類似,與重度干旱處理的日變化有較大不同:對照和中度干旱處理主要表現為午間降幅大,低谷期較短,在大氣蒸散力較弱的04:00—08:00時段變幅較小,在蒸散力和蒸騰速率迅速增大的09:00—14:00時段持續快速下降,午后降至最低值,隨后回升形成窄而尖的谷型;而重度干旱處理在04:00—08:00時段即快速下降,10:00—14:00時段基本維持在最低水平,然后緩慢回升,形成寬而緩的谷型,其午間降幅雖小,但低谷期卻明顯提前并后延. 總體而言,隨著土壤有效水分供應程度(土壤水勢)的遞降,髯毛箬竹成熟葉片水勢的動態水平亦降低,延長了低谷期,延遲了葉片水分虧缺的時間,這種情況與高溫組和極端高溫組相似,極端高溫+重度干旱處理表現的尤為明顯,不僅植株葉片水勢在高蒸騰作用時段(10:00—14:00)處于最低水平,且04:00和20:00的成熟葉片水勢亦顯著低于極端高溫和極端高溫+中度干旱處理.
對照、高溫和極端高溫處理在主要蒸騰時段(10:00—16:00)內表現出強烈的抑制,而成熟葉片水勢則基本維持在較低的水平,表明植株的葉片啟動自身的水流調節機制,沒有繼續降低水勢,對保護包括光合組織以及其他細胞內部結構起到關鍵作用. 中度干旱及其復合處理、重度干旱及其復合處理午間細根水勢的強烈降低,表明在土壤水分虧缺狀態下植株細根水流調節或水勢維持機制尚未形成響應.
2.3 不同處理條件下髯毛箬竹細根與成熟葉片的抗氧化酶活性
由圖3可知,在對照組中,細根SOD活性以中度干旱最高,但與對照處理差異不顯著(P>0.05),而后隨著干旱程度的加劇,SOD活性顯著下降(P<0.05),說明重度干旱下,SOD系統受到了破壞,這與高溫組和極端高溫組中SOD活性的表現相同,各組之間的相同處理差異不顯著(P>0.05). 成熟葉片的SOD活性與細根類似,但在極端高溫組中,極端高溫+中度干旱處理的SOD活性均略低于極端高溫,可能在處理下,SOD系統受到了一定的破壞. 對照組的細根CAT活性在中度干旱時平均值升高,而重度干旱處理則出現顯著下降(P<0.05),高溫組的CAT活性與對照組相同,即先升后降的趨勢,說明在常溫和高溫的情況下,髯毛箬竹在重度干旱下會出現生理過程受抑制的現象. 而極端高溫組的細根CAT活性在中度干旱時就出現了顯著下降(P<0.05),在重度干旱時極顯著下降(P<0.01),說明高溫對細根CAT活性有顯著影響. 而極端高溫組的成熟葉片在極端高溫+重度干旱處理時CAT活性平均值下降,仍表現為差異不顯著(P>0.05). 細根和成熟葉片的POD與CAT活性的情況類似,在對照組和高溫組中,細根的POD活性都表現出先升后降的趨勢,且在重度干旱、高溫+重度干旱處理時顯著下降(P<0.05),細根與成熟葉片POD活性平均值在極端高溫+中度干旱處理時有所下降. 因此,就3種抗氧化酶活性的平均值而言,高溫、極端高溫、中度干旱和高溫+中度干旱處理對提升細根與成熟葉片的抗逆能力有一定的積極作用,而重度干旱及其復合處理的平均值表明其對細根和成熟葉片3種抗氧化酶有抑制影響.

表3 不同處理組中髯毛箬竹細根與成熟葉片水勢日變化
注:數據為平均值±標準差;以絕對值大小進行LSD小寫字母編排,不同小寫字母表示差異顯著(P>0.05).

注:數據為平均值±標準差;不同小寫字母表示差異顯著(P>0.05).圖3 不同處理組中髯毛箬竹細根和成熟葉片的抗氧化酶系統Fig.3 Antioxidant enzyme system of I. barbatus′s fine roots and mature leaves under different treatment groups
3 討論
3.1 高溫干旱對髯毛箬竹蒸騰、光合速率、氣孔導度和水分利用效率的影響
植物處于最適溫度與土壤水分條件時表現出最大的光合反應速率[17]. 該研究表明,重度干旱及其復合處理嚴重影響髯毛箬竹的正常生理進程,高溫與極端高溫加劇了這一影響,具體表現為蒸騰速率、光合速率、氣孔導度和水分利用效率顯著下降幾倍或幾十倍;與對照處理相比,但高溫明顯提升了髯毛箬竹葉片的蒸騰速率,氣孔導度也隨之略微升高;極端高溫則導致光合速率的明顯下降,并收窄光合速率日進程變化的幅度,而氣孔導度和蒸騰速率并沒有下降的趨勢,反而二者的平均值有所上升,可能是因為高溫、極端高溫試驗條件下供水充足有關,這與Ranney[18]在25~40 ℃葉溫范圍內,對水分供應充足的白樺(Betulaplatyphylla)葉片的蒸騰作用、光合作用和氣孔行為的研究結果類似.這是在供水充足的條件下,植物通過增加氣孔導度來提高蒸騰作用并降低葉片溫度是植物對高溫環境的一種積極保護措施[10]. 同時,作為協調碳同化和水分耗散關系的葉片水分利用效率亦佐證了非氣孔限制,該試驗發現,高溫處理從10:00以后和極端高溫處理07:00—18:00的水分利用效率平均值小于對照處理,說明僅蒸騰速率得到了提升,固定碳水化合物能力則下降,即比正常環境的植物遭受了較高的蒸發需求,而植物水分利用效率則較低[12]. 在涉及干旱及其復合處理的條件下,光合速率與蒸騰作用、氣孔導度同時降低,光合速率比蒸騰速率減小的幅度大,表明水分利用效率降低使葉綠體內CO2的供應受阻,即氣孔限制因素導致[19-20].
相比較各自對照組的各處理(即對照、高溫和極端高溫),中度干旱及其復合處理的葉片水分利用效率沒有明顯下降,高溫+中度干旱處理甚至顯著大于高溫處理,這可能是水分可利用性的降低導致通過增強氣孔關閉來減弱蒸騰作用,從而提高了植物的葉片水分利用效率. 因此,通過適當的增溫來馴化和栽培抗高溫物種可改變植物氣體交換過程[17,20]. 該研究發現,髯毛箬竹蒸騰、光合速率和氣孔導度對不同高溫干旱條件的響應不同,髯毛箬竹的抵抗極端高溫方面表現尚可,極端高溫+中度干旱處理并不威脅其生存,但不適宜長期重度干旱的環境;高溫、中度干旱及高溫+中度干旱處理有利于髯毛箬竹的馴化栽培,特別是在高溫+中度干旱的復合作用下,并不影響植物正常生長,甚至一定長度上能促進其生長. 故適度控制環境溫度和土壤含水量,在未來園林景觀物種配置中可以得到應用.
3.2 高溫干旱對髯毛箬竹細根和成熟葉片水勢的影響
植物不同器官對適應環境的策略不同,甚至相反[21],Dichio等[22]研究橄欖樹(Oleaeuropaea)時發現,根系和葉片的共同作用才能使其在水分脅迫下維持光合作用;而植物根系與葉片水勢受土壤水分條件和溫度等因素的綜合影響[23]. 因此,有必要深入理解植物不同器官適應高溫干旱脅迫的機制和策略[19,22,24]. 該研究發現,除了高溫+重度干旱和極端高溫+重度干旱這兩種處理已經對植株生理造成破壞,其他處理的髯毛箬竹都出現了蒸騰午休現象,但細根和成熟葉片水勢并沒有出現這種波動,二者水勢皆表現為單谷型日變化進程,這可能是由于植株本身水分交換、運輸過程和水容效應,其根系吸水與蒸騰并不完全同步,吸水滯后于蒸騰[25]. 細根與成熟葉片水勢在14:00左右開始回升,這與崔曉陽等[14]試驗結果近似,表明髯毛箬竹各組織吸釋水更替時間在14:00左右;對照、中度干旱、高溫、高溫+中度干旱和極端高溫條件下髯毛箬竹細根和成熟葉片水勢在20:00時可以得到有效恢復,隨著干旱程度上升,細根和成熟葉片水勢顯著下降,高溫加劇這一趨勢,增加了細根和成熟葉片恢復正常水勢的成本. 同時,土壤水勢對細根水勢的日動態谷型跨度及恢復進程的影響與成熟葉片水勢類似,但土壤水勢對二者日變幅的影響卻不同,即較高的土壤水勢使成熟葉片水勢和細根水勢的高值出現小幅變動,較低的土壤水勢導致葉水勢的低值出現小幅變動、根水勢的低值出現大幅變動,這在10:00—16:00之間表現得最為明顯,尤其是在重度干旱及其復合處理條件(毛管水斷裂的土壤水分狀態)下,細根水勢的下降要比葉水勢更強烈. 因此,在水勢方面,吸收器官對提升的大氣蒸散力的間接反應比蒸騰器官的直接反應敏感,這意味著細根吸水遇到了較大阻力或者呈現虧缺狀態[12,24].
3.3 高溫干旱對髯毛箬竹保護酶系統的影響
研究[26]表明,當植物長期處在逆境環境中,體內的活性氧代謝失調,SOD、POD和CAT的活性水平會發生變化. 筆者發現,隨著溫度的增加,髯毛箬竹通過增加體內SOD、POD和CAT活性來消除體內的活性氧,免受高溫傷害,在極端高溫處理下其酶防御系統仍未受損;中度干旱及其復合處理條件下,SOD、POD和CAT活性未顯著下降,說明高溫和極端高溫對土壤水分和植株蒸騰拉升作用并未嚴重影響植株的保護酶系統;而重度干旱及其復合處理的3種保護酶均表現出顯著下降,葉片的光合能力大幅下降,活性氧物質超出了保護酶調節能力的限度. 因此,髯毛箬竹具有較好的抗高溫和抗旱能力,髯毛箬竹的抗氧化酶活性證明了其可以在中度干旱、高溫或極端高溫中生存,一定范圍內的高溫和干旱能刺激其抗氧化酶活性的提升,其器官組織受到的氧化損傷并不是不可修復;對于重度干旱而言,用于光化學反應的能量大量下降,形成更多的活性氧物質使其細胞膜系統受損程度嚴重,而高溫使這種損害更加嚴重.
細根的活力在很大程度上決定著植物的存活和抗逆能力. 安玉艷等[19]對杠柳(Peripiocasepium)幼苗的研究中發現,幼葉、老葉和根的SOD、CAT和POD活性及變化趨勢不同,以細根的保護酶反應最為靈敏. 該研究發現,高溫和干旱對髯毛箬竹細根和成熟葉片SOD、POD和CAT活性的影響雖然較為一致,但是細根與成熟葉片3種抗氧化酶的活性差異顯著,其中細根的SOD活性明顯大于成熟葉片,而CAT和POD活性則相反,在保護系統中處于核心地位的SOD活性顯著大于成熟葉片,表明植株細根在活性氧代謝及清除活性氧的能力提升[16],而后者則表明成熟葉片在生理生化代謝及生長發育情況優于細根,說明細根是髯毛箬竹適應高溫干旱環境的重要器官,器官間的生理整合作用與協調使其能有效地適應干濕交替的高溫環境. 因此,在逆境下,髯毛箬竹優先保護生命力旺盛的組織,以提高逆境下的生存能力[13],不同組織和器官在抵抗高溫干旱脅迫中的作用不同. 髯毛箬竹細根高效地調節保護酶系統的能力,是其適應未來極端環境的重要生理生態機制.
4 結論
a) 在土壤水分充足的情況下,高溫提升了髯毛箬竹蒸騰速率和氣孔導度,非氣孔限制因素是極端高溫條件下光合速率下降的主要原因;氣孔限制因素是干旱及其復合條件下導致光合速率降低的主要原因.
b) 細根和成熟葉片水勢日進程皆表現為單谷型,吸釋水更替時間均在14:00左右;在土壤毛管水斷裂狀態下,吸收器官細根對提升的大氣蒸散力的間接反應比蒸騰器官成熟葉片的直接反應更為敏感.
c) 高溫和極端高溫并未嚴重影響植株的保護酶系統,一定范圍內的高溫和干旱能提升其抗氧化酶活性的提升;細根在活性氧代謝及清除活性氧的能力優于成熟葉片,是適應高溫干旱環境的重要器官.
d) 生理生態參數、器官水勢指標和抗氧化酶活性證明了髯毛箬竹具有較好的抗高溫和抗旱能力;重度干旱及其復合處理嚴重影響植株生理進程,高溫加劇這一進程.
[1] WHITE T A,CAMPBELL B D,KEMP P D.Impacts of extreme climatic events on competition during grassland invasions[J].Global Change Biology,2001,7(7):1-13.
[2] 王維,王文杰,李俊生,等.基于歸一化差值植被指數的極端干旱氣象對西南地區生態系統影響遙感分析[J].環境科學研究,2010,23(12):1447-1455.
WANG Wei,WANG Wenjie,LI Junsheng,etal.Remote sensing analysis of impacts of extreme drought weather on ecosystems in southwest region of china based on normalized difference vegetation index[J].Research of Environmental Sciences,2010,23(12):1447-1455.
[3] LLORET F,ESCUDERO A,IRIONDO J M,etal.Extreme climatic events and vegetation:the role of stabilizing processes[J].Global Change Biology,2012,18(3):797-805.
[4] 李偉成,周妍,丁雨龍,等.毛竹種子萌發及幼苗生長對環境條件的可塑性響應[J].竹子研究匯刊,2008,27(4):23-27.
LI Weicheng,ZHOU Yan,DING Yulong,etal.The plastic response of seed germination and seedling growth to environment,a study on moso bamboo[J].Journal of Bamboo Research,2008,27(4):23-27.
[5] WANG Jinchuan,DUAN Baoli,ZHANG Yuanbin.Effects of experimental warming on growth,biomassallocation,and needle chemistry ofAbiesfaxonianain even-aged monospecific stands[J].Plant Ecology,2012,213(1):47-55.
[6] 劉國華,王福升,郭婷婷,等.4種地被竹的蒸騰特性及蒸發速率的比較[J].南京林業大學學報(自然科學版),2012,36(6):147-150.
LIU Guohua,WANG Fusheng,GUO Tingting,etal.Comparison of the transpiration parameters and evaporation rates of four dwarf bamboos with each other[J]. Journal of Nanjing Forestry University(Natural Science Edition),2012,36(6):147-150.
[7] 高貴賓,鐘浩,吳志莊,等.不同混生地被竹光合和熒光特征比較[J].福建農林大學學報(自然科學版),2016,45(5):515-521.
GAO Guibin,ZHONG Hao,WU Zhizhuang,etal.Comparative study on photosynthetic and fluorescence characteristics of different amphipodial dwarf bamboo[J].Journal of Fujian Agriculture and Forestry University (Natural Science Edition),2016,45(5):515-521.
[8] 李娟,彭鎮華,高健,等.干旱脅迫下黃條金剛竹的光合和葉綠素熒光特性[J].應用生態學報,2011,22(6):1395-1402.
LI Juan,PENG Zhenhua,GAO Jian,etal.Photosynthetic parameters and chlorophyll fluorescence characteristics ofPleioblastuskongosanensf.aureostriausunder drought stress[J].Chinese Journal of Applied Ecology,2011,22(6):1395-1402.
[9] BEEBE S E,RAO I M,DEVI M J,etal.Commonbeans,biodiversity,and multiple stresses:challenges of drought resistance in tropical soils[J].Crop and Pasture Science,2017,65(7):667-675.
[10] XU Zhenzhu,ZHOU Guangsheng,HAN Guangxuan,etal.Photosynthetic potential and its association with lipid peroxidation in response to high temperature at different leafages in maize[J].Journal of Plant Growth Regulation,2011,3(1):41-50.
[11] MITTLER R.A biotic stress,the field environment and stress combination[J].Trends in Plant Science,2006,11(1):15-19.
[12] KURSAR T A,ENGELBRECHT B M J,BURKE A,etal.Tolerance to low leaf water status of tropical tree seedlings is related to drought performance and distribution[J].Functional Ecology,2009,23(1):93-102.
[13] CARVALHO L C,COITO J L,GON?ALVES E F,etal.Differential physiological response of the grapevine varieties Touriga Nacional and Trincadeira to combined heat,drought and light stresses[J].Plant Biology,2015,18(S1):101-107.
[14] 崔曉陽,宋金鳳,屈明華.土壤水勢對水曲柳幼苗水分生態的影響[J].應用生態學報,2004,15(12):2237-2244.
CUI Xiaoyang,SONG Jinfeng,QU Minghua.Effect of soil water potential on hydraulic parameters ofFraxinusmandshuricaseedlings[J].Chinese Journal of Applied Eco1ogy,2004,15(12):2237-2244.
[15] 張佳華,張健南,姚鳳梅,等.開放式增溫對東北稻田生態系統作物生長與產量的影響[J].生態學雜志,2013,32(1):15-21.
ZHANG Jiahna,ZHANG Jiannan,YAO Fengmei,etal.Effects of free air temperature increasing on the rice growth and grain yield in northeast China[J].Chinese Journal of Ecology,2013,32(1):15-21.
[16] 郝文芳,趙潔,蔡彩虹,等.3種胡枝子抗氧化酶和滲透調節物質對干旱和增強UV-B輻射的動態響應[J].環境科學學報,2013,33(8):2349-2358.
HAO Wenfang,ZHAO Jie,CAI Caihong,etal.Dynamic response of antioxidant enzymes and osmotic adjustment substances to drought and enhanced UV-B radiation in three Lespedezas[J].Acta Scientiae Circumstantiae,2013,33(8):2349-2358.
[17] TAYLOR S H,FRANKS P J,HULME S P,etal.Photosynthetic pathway and ecological adaptation explain stomatal trait diversity amongst grasses[J].New Phytologist,2012,193:387-396.
[18] RANNEY T G.Heat tolerance of five taxa of birch (Betula):physiological responses to supraoptimal leaf temperatures[J].American Horticulture Science,1994,119(2):243-248.
[19] 安玉艷,郝文芳,龔春梅,等.干旱-復水處理對杠柳幼苗光合作用及活性氧代謝的影響[J].應用生態學報,2010,21(12):3047-3055.
AN Yuyan,HAO Wenfang,GONG Chunmei,etal.Effects of drying and re-watering on the photosynthesis and active oxygen metabolism ofPeripiocasepiumseedlings[J].Chinese Journal of Application Eco1ogy,2010,21(12):3047-3055.
[20] 朱玉,黃磊,鄭云普,等.高溫對高叢越橘葉片氣孔特征和氣體交換參數的影響[J].果樹學報,2016,33(4):444-456.
ZHU Yu,HUANG Lei,ZHENG Yunpu,etal.Effects of high temperatures on leaf stomatal traits and gas exchanges of highbush blueberries[J].Journal of Fruit Science,2016,33(4):444-456.
[21] ROESSNER U,PATTERSON J H,FORBES M G,etal.An investigation of boron toxicity in barley using metabolomics[J].Plant Physiology,2006,142(3):1087-1101.
[22] DICHIO B,XILOYANNIS C,SOFO A,etal.Osmotic regulation in leaves and roots of olive trees during a water deficit and rewatering[J].Tree Physiology,2006,26(2):179-185.
[23] 康紹忠,張建華,梁建生.土壤水分與溫度共同作用對植物根系水分傳導的效應[J].植物生態學報,1999,23(3):211-219.
KANG Shaozhong,ZHANG Jianghua,LIANG Jiansheng.Combined effects of soil water content and temperature on plant root hydraulic conductivity[J].Acta Phytoecologica Sinica,1999,23(3):211-219.
[24] BLESSING C H,WERNER R A,SIEGWOLF R,etal.Allocation dynamics of recently fixed carbon in beech saplings in response to increased temperatures and drought[J].Tree Physiology,2015,35(6):585-591.
[25] MEMMI H,GIJN M C,COUCEIRO J F,etal.Water stress thresholds for regulated deficit irrigation in pistachio trees:rootstock influence and effects on yield quality[J].Agricultural Water Management,2015,164:58-72.
[26] LIU Jianjun,LI Junqi,SU Xinhong,etal.Grafting improves drought tolerance by regulating antioxidant enzyme activities and stress-responsive gene expression in tobacco[J].Environmental and Experimental Botany,2014,107(22):173-179.
ImpactofDrought,Heat,andTheirCombinationontheEco-PhysiologicalCharacteristicsofLeavesandFineRootsinDwarfBamboo(Indocalamusbarbatus)
LI Weicheng1,2, SHENG Haiyan3, GAO Guibin1, WEN Xing1, TIAN Xinli1*
1.Key Laboratory of High Efficient Processing of Bamboo of Zhejiang Province, China National Bamboo Research Center, Hangzhou 310012, China2.College of Life Sciences, Southwest Forestry University, Kunming 650224, China3.Hangzhou Environmental Protection Science Institute, Hangzhou 310005, China
Drought and high temperature are two main stress factors limiting plant productivity, and often occur simultaneously in nature. To determine the effects of high temperature and drought on plant development and productivity, we investigated the effects of abiotic stress on the growth and quality of dwarf bamboo (Indocalamusbarbatus). Specifically, we examined the relationships between simulated stress conditions (high temperature and drought) and eco-physiological processes (leaf gas exchange, water potential of leaves and fine roots, and the antioxidant enzymesystem) in dwarf bamboo. We simulated natural high temperature and drought through heating, dehydrating, and rehydrating processes, using the following three treatment groups: control group (control, moderate drought, and severe drought), high temperature group (high temperature, high temperature+moderate drought, and high temperature + severe drought), and extreme heat group (extreme heat, extreme heat + moderate drought, and extreme heat + severe drought).Our results indicated that the responses ofI.barbatusvaried significantly under the different simulated temperature and drought conditions. The photosynthetic rate of control plants under normal temperature (7-28 °C) and normal soil water content (37.86%±5.95%) was (7.141±0.072) μmolm2·s. High temperature (30-35 °C) significantly increased stomatal conductance and the transpiration rate. Under extreme heat (35-40 °C), non-stomatal-limiting factors led to a decrease in photosynthetic rate, which minimized diurnal variations in photosynthesis. Under drought conditions (soil water content 27.95%±2.64%) and its combined treatments with temperature, stomatal-limiting factors led to a decrease in photosynthetic rate. Severe drought (soil water content 18.25%±1.73%)and its combined treatments with temperature markedly affected physiological processes, and high temperature intensified this effect further, due to significant decreases in transpiration, photosynthesis, stomatal conductance, and water-use efficiency. The water potentials of fine roots in the control, high temperature, and extreme heat groups were (-0.111±0.033) to (-0.961±0.086), (-0.173±0.060) to (-0.970±0.072), and (-0.304±0.061) to (-1.225±0.166)MPa, respectively. The water potential of fine roots decreased more markedly than that of mature leaves. The influences of high temperature to three antioxidant enzymes in the fine roots and mature leaves were similartothat of drought stress. The superoxide dismutaseactivity of fine roots was higher than that of mature leaves, whereas in contrast the activities of catalaseand peroxidase were higher in mature leaves. We accordingly conclude that the water potential and antioxidant enzyme activities of leaves and fine roots are useful indicators for estimating the performance of dwarf bamboo under stress conditions. In addition, the absorbing organ (fine root) is more sensitive than the transpiration organ (leaf), and the fine roots inI.barbatusare important tissues for adapting to high temperature and drought conditions. Dwarf bamboo plants showed good tolerance to high temperature and drought stress. Within a certain range of high temperature and drought, dwarf bamboo triggered stress response mechanisms and enhanced antioxidant enzyme activity, particularly under combined treatments. High temperature and drought had no negative effects on growth, and extreme heat + moderate drought did not affect plant health. High temperature and moderate drought, individually and in combination, were found to be favorable for domestication and cultivation. Severe drought decreased leaf gas exchange, and affected leaf and fine root physiological processes, whereas high temperature aggravated the damage caused by drought. On the basis of these results, we conclude that dwarf bamboo is not suitable for growth in regions that experience extreme drought conditions.
combined conditions; dwarf bamboo; gas exchange; water potential; antioxidant enzymes
2017-04-29
2017-10-16
浙江省自然科學基金項目(LY14C030008); 浙江省科技計劃項目(2016F50008)
李偉成(1978-),男,浙江湖州人,副研究員,博士,主要從事系統生態學研究,lee_caf@163.com.
*責任作者,田新立(1981-),男,山東滕州人,高級工程師,博士,主要從事生態學研究,277918782@qq.com
李偉成,盛海燕,高貴賓,等.高溫干旱對髯毛箬竹的葉和細根的生理生態影響[J].環境科學研究,2017,30(12):1908-1918.
LI Weicheng,SHENG Haiyan,GAO Guibin,etal.Impact of drought, heat, and their combination on the eco-physiological characteristics of leaves and fine roots in dwarf bamboo (Indocalamusbarbatus)[J].Research of Environmental Sciences,2017,30(12):1908-1918.
X173;S718.4
1001-6929(2017)12-1908-11
A
10.13198j.issn.1001-6929.2017.03.57
