MITFa及TYR基因在紅色錦鯉體色發生不同階段的表達分析
2017-12-18 08:43:10龐小磊王良炎郭生強李學軍
水產科學 2017年2期
關鍵詞:研究
田 雪,龐小磊,王良炎,顧 靚,郭生強,李學軍
( 河南師范大學 水產學院, 河南 新鄉 453007 )
MITFa及TYR基因在紅色錦鯉體色發生不同階段的表達分析
田 雪,龐小磊,王良炎,顧 靚,郭生強,李學軍
( 河南師范大學 水產學院, 河南 新鄉 453007 )
本研究描述了紅色錦鯉從出膜到體色形成的過程,總結和歸納不同發育階段體色變化異同點,篩選出6個體色變化較顯著時期,分別為1、2、3、4、12、48 日齡。利用熒光定量PCR分析MITFa及TYR基因在紅色錦鯉6個體色變化時期的表達情況。結果顯示,MITFa基因在1日齡時表達量最高,顯著高于48日齡時(P<0.05),極顯著高于其他4個時期(P<0.01)。48日齡時表達量次之,亦極顯著高于2、3、4、12日齡 4個時期(P<0.01)。2、3、4、12日齡 4個時期MITFa基因表達量較低且不存在顯著差異(P>0.05)。TYR基因在1日齡時表達量最高(P<0.01),4日齡時表達量顯著高于12、48日齡(P<0.05),與2、3日齡不存在顯著差異(P>0.05)。TYR基因表達量在2、3、12、48日齡 4個時期差異不顯著(P>0.05)。MITFa和TYR基因在紅色錦鯉體色形成過程中,表達水平整體呈現降低的趨勢,其中MITFa基因表達量表現為先降后升,TYR基因則先降后升再降。以上結果顯示MITFa、 TYR基因與錦鯉體色形成具有一定相關性,但MITFa和TYR基因在體色發生中的相互作用還有待進一步研究證實。
紅色錦鯉;MITFa;TYR;表達分析;體色發生
魚類體色豐富多彩,種類、性別、個體差異、甚至個體發育的不同階段和部位都可能表現出不同顏色。除了種類、生理生化因素使魚類體色產生差異,其生存的水域環境,攝食的餌料等一系列因素也可影響魚類體色的形成[1-2]。環境、理化因子對魚類體色的影響主要因其皮膚和鱗片上的色素細胞種類、數量和分布不同所致[3-6]。魚類具有6種色素細胞,分別是黑色素細胞、紅色素細胞、黃色素細胞、虹彩細胞、白色素細胞和藍色素細胞。6種色素細胞含有不同的色素顆粒,色素顆粒的顏色、反光作用和運動遷移,使魚類呈現從白到黑的多種顏色[7]。
魚類色素細胞由神經嵴細胞分化形成,大量基因參與神經嵴細胞向色素細胞的遷移和分化[8]。MITF基因是黑色素細胞分化形成的重要轉錄因子,在魚類色素細胞分化和成熟過程中發揮重要作用。TYR是黑色素合成的關鍵限速酶,調控黑色素顆粒的生成,MITF基因結合在下游TYR基因啟動子區域,促進TYR基因的表達[9-12]。
錦鯉(Cyprinuscarpio),屬鯉形目、鯉科、鯉屬,是鯉魚人工選育過程形成的觀賞品種[13-14]。作為高檔觀賞魚,錦鯉在其體色鑒賞上已形成一定系統[15],常見種類主要包括紅白、大正三色、別光等。但是,這些類型體色多成斑塊狀或鑲嵌型,遺傳背景復雜。因此,本試驗選用體色單一的紅色錦鯉作為研究對象,觀察其體色發生過程,并對其個體發育不同階段MITFa和TYR基因進行熒光定量分析,探索MITFa和TYR基因與錦鯉體色發生的關系,為研究錦鯉體色形成機制提供理論依據,也為魚類色素細胞系的建立提供基礎研究資料。
1 材料與方法
1.1 試驗魚
2015年3月從河南師范大學水產養殖基地挑選性腺發育良好的紅色錦鯉親魚6尾(♀∶♂=1∶1)進行人工干法授精,受精卵置于孵化桶孵化,用于仔魚體色發生觀察。
1.2 仔魚體色發生觀察和樣品收集
仔魚出膜后,0~25 日齡每日觀察、取樣一次,25~60 日齡每2 d取樣一次。隨機取樣30尾仔魚,10尾在OLYMPUS BX51研究級顯微鏡下觀察、拍照,20尾用RNAlater(Life technologies公司,美國)保存于-80 ℃,用于RNA提取。
1.3 總RNA提取和RT-PCR擴增
依據體色發生觀察結果,對1、2、3、4、12、48日齡的紅色錦鯉仔魚進行RNA提取,1%瓊脂糖凝膠電泳和ND-2000核酸蛋白儀檢測RNA完整性、純度和OD值。PrimeScriptTMRT reagent Kit with gDNA Eraser(Takara公司,大連)合成cDNA用于PCR擴增。
利用Primer3Plus在線軟件設計MITFa和TYR引物(http://www.primer3plus.com/)(表1),由蘇州金唯智生物公司合成。取適量cDNA為模板,進行PCR擴增。PCR產物經瓊脂糖凝膠電泳檢測,連接pMD18-T載體(Takara公司,大連),轉化DH5α感受態細胞,經藍白斑篩選及雙酶切驗證的陽性克隆送蘇州金唯智生物有限公司進行測序。

表1 試驗用PCR 引物及序列
1.4 實時熒光定量PCR (qRT-PCR)
參照TaKaRa熒光定量PCR試劑盒說明書(TaKaRa,大連),將6個發育階段的紅色錦鯉cDNA作為模板,設3個重復,體系為10 μL: 上下游引物各0.3 μL,SYBR Premix Ⅱ TaqTM(2×) 5 μL,cDNA 1 μL,RNase free H2O 3.4 μL。qRT-PCR反應條件如下:95 ℃預變性30 s,95 ℃變性5 s,62 ℃ 20 s, 40 個循環。反應結束分析溶解曲線和Ct值,2-△△Ct法計算每個階段MITFa和TYR基因的相對表達水平,△Ct=Ct目的基因-Ctβ-actin,△△Ct=(Ct目的基因-Ctβ-actin)試驗組-(Ct目的基因-Ctβ-actin)對照組。
1.5 統計學分析
利用SPSS 20.0單因素方差分析表達量數據并進行LSD和Duncan氏比較,結果用平均值±標準差表示。
2 結 果
2.1 紅色錦鯉體色發生觀察
紅色錦鯉出膜前時期,受精卵依靠卵黃囊提供能量用來發育,此時胚胎呈現卵黃囊顏色,表現為微弱黃色。1日齡仔魚除眼部呈現黑色外,其他部位無色素細胞,魚苗整體表現為無色透明。仔魚發育至2日齡,顯微鏡下可觀察到體色變化,色素細胞集中在錦鯉背部,從頭部逐漸向尾部擴散。發育至3日齡,仔魚背部顏色加深,腹部開始出現色素細胞。4日齡,色素細胞完全分布在背部及腹部。5~14日齡,體色逐漸加深,發育至48日齡,鱗片完全形成,魚體呈現紅色,基本同成魚顏色一致(圖1)。

圖1 不同發育時期的紅色錦鯉a,1日齡; b,2日齡;c,3日齡;d,4日齡;e,12日齡;f,48日齡.1,背部;2,腹部;3,眼.
2.2 錦鯉體色發生過程中MITFa基因的表達
通過紅色錦鯉的體色發生觀察,篩選出6個關鍵時期。檢測MITFa基因在1、2、3、4、12、48日齡的表達情況,熒光定量PCR結果顯示,MITFa基因在1日齡時表達量是1.053±0.409,顯著高于48日齡(P<0.05),1、48 日齡 MITFa表達量均極顯著高于其他4個時期(P<0.01)。12日齡時表達量最低,但與2、3、4日齡 3個發育階段不存在顯著差異(P>0.05)(圖2)。
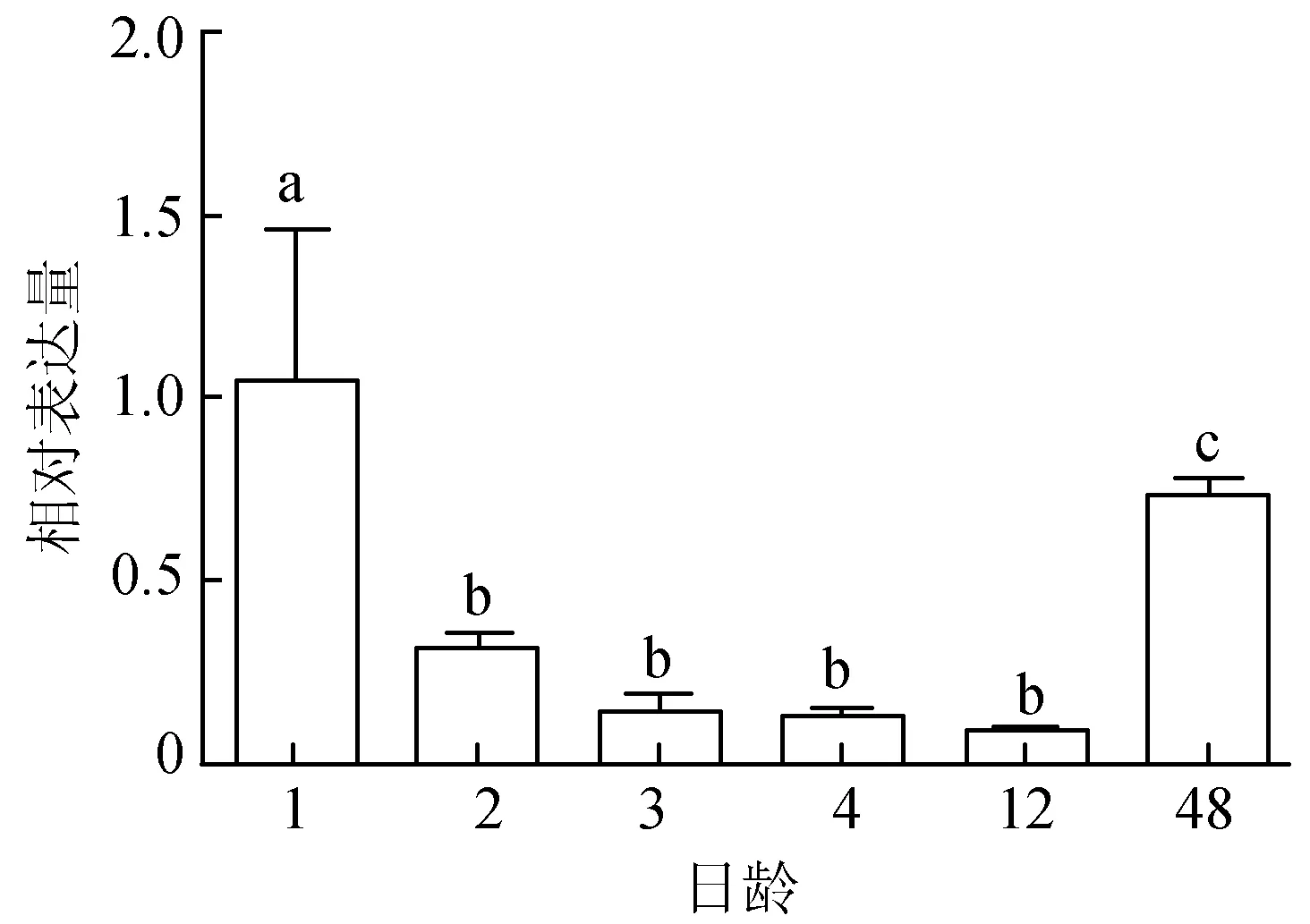
圖2 錦鯉體色發生過程中MITFa基因的表達
2.3 錦鯉體色發生過程中TYR基因的表達
熒光定量PCR結果顯示,TYR基因在紅色錦鯉上述6個體色發生階段的表達情況整體呈降低趨勢。1日齡TYR基因表達量是1.088±0.572,極顯著高于其他5個時期(P<0.01),4日齡表達量顯著高于12日齡、48日齡(P<0.05),而與2日齡、3日齡這兩個時期不存在顯著差異(P>0.05),2、3、12、48日齡 4個時期TYR基因表達量不存在顯著差異(P>0.05),48日齡 TYR基因的表達量最低,為0.0388±0.00613(圖3)。
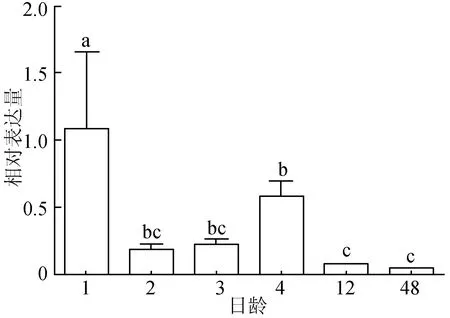
圖3 錦鯉體色發生過程中TYR基因的表達
3 討 論
3.1 紅色錦鯉不同發育時期體色發生觀察
利用顯微觀察方法對紅色錦鯉出膜前、出膜后及體色完全形成等一系列體色發生過程進行了詳細描述。出膜前時期受精卵依靠卵黃囊提供能量進行發育,胚胎呈現卵黃囊顏色,表現為微弱黃色,與一般魚類胚胎顏色相近[4]。出膜后1日齡仔魚除眼部呈現黑色,其他部位無色素細胞,身體透明,可能是魚體內色素細胞還未開始分化發育或剛開始分化,色素顆粒尚未合成,導致整體表現為無色透明[16]。發育至2日齡,隨著黃色素和紅色素細胞不斷分化、發育,可在顯微鏡下觀察到體色變化,色素細胞集中呈現于錦鯉背部,從頭部逐漸向尾部擴散[17]。這說明色素細胞分化存在時空差異,隨著時間發展,不斷向尾部遷移擴散[18]。仔魚發育至3日齡,背部顏色加深,腹部開始出現色素細胞,這可能是由于色素細胞分化的同時,伴隨著色素顆粒的合成和積累。Hultman等[19]研究證實魚類色素細胞是由神經嵴細胞通過背腹軸模式遷移,本研究中紅色錦鯉體色發生同樣存在背腹軸模式。發育至4日齡,色素細胞完全分布在背部及腹部,同樣也是黃色素細胞和紅色素細胞相互作用的過程[20]。5~14日齡,隨著色素顆粒的沉著,體色逐漸加深。發育至48日齡,鱗片完全形成,魚體呈現紅色,基本同成魚顏色一致。
3.2 MITFa基因和TYR基因在發育過程中的表達差異
MITFa基因在紅色錦鯉體色發生的6個時期均有一定量的表達。這與Liu等[12]研究結果相似,即在無黑色素細胞的錦鯉囊胚、剛出膜仔魚、紅白錦鯉和黃金錦鯉中均檢測到MITFa基因。已有的研究表明,MITFa基因不僅參與黑色素細胞發育與分化,而且在體色形成相關的非黑色素細胞分化及成體體色維持中扮演重要角色[9]。發育至1日齡, MITFa基因表達量最高,可能是由于色素前體細胞向色素細胞分化過程中MITFa基因發揮一定作用導致[11]。隨著色素細胞不斷分化成熟,色素細胞遷移和擴散逐漸減慢,MITFa基因表達水平也呈降低趨勢[21]。
TYR是一種廣泛存在于植物、動物和細菌中的跨膜轉運蛋白,主要存在于黑色素小體中,僅有一小部分在細胞質。TYR基因在紅色錦鯉體色發生階段都有存在,與王巍等[10,22]研究結果相似。甌江彩鯉、純色及黑色斑塊錦鯉皮膚中均存在TYR基因的表達,說明TYR不僅在黑色素細胞中起作用,在其他類型色素細胞中也發揮一定功能[23]。MITFa和TYR基因均在1日齡表達量達到最高,顯著高于其他5個時期,進一步表明在紅色錦鯉體色形成初期,MITFa基因對TYR具有上調效應。隨著體色的形成,MITFa基因表達量呈遞減趨勢,TYR基因表達量同樣表現為逐漸降低。但4日齡TYR基因表達量相對增加,原因可能由于錦鯉體內存在多種酪氨酸酶形式,部分抑制狀態在這個時期被激活,導致表達量增高[24- 25]。
近年來,關于錦鯉體色研究主要集中在色素著色物質及理化因素對魚類體色的影響方面[1-2,26-27],而對于色素相關基因的研究大多從黑色素細胞色素合成關鍵基因入手,涉及色素細胞遷移和分化的研究,較為鮮見[9-10,12],關于MITF及TYR基因的研究主要集中在哺乳動物黑色素細胞方面[28],MITF基因可調控酪氨酸基因家族的表達,從而參與黑素生成的調控, MITF基因在生物體內能調控黑色素合成關鍵限制酶——酪氨酸酶基因的表達,影響黑色素細胞的分化[29]。關于這兩個基因在魚類不同發育階段的研究較少,如Darias等[30]在其研究中所報道的那樣,塞內加爾鰨(Soleasenegalensis)不同發育階段過程中,MITF基因的表達也表現為先降后升的趨勢,同時TYR基因的表達也表現為兩個峰值,且其表達趨勢也與在錦鯉中的表達顯示出一定的相似性。但這兩種基因對于其他色素細胞分化和形成是否起作用,以及如何發揮功能均未見報道。
錦鯉色素調控機制非常復雜,關于錦鯉色素調控通路、色素細胞的相互作用及其模式形成已開展相關研究[8, 19, 31-33]。在這些研究中也都提及色素細胞的調控不是由單因素決定,但對于參與黑色素細胞形成及色素合成的調控基因是否在其他類型色素細胞中發揮作用,所開展的研究相對較少。本研究對錦鯉體色發生過程進行顯微觀察,并分析MITFa和TYR基因在錦鯉體色發生過程中的表達水平,為將來深入研究色素形成相關基因在錦鯉體色形成的作用提供基礎試驗數據,但MITFa和TYR基因在非黑色素細胞中的作用及相互關系還有待進一步深入研究。
4 結 論
本研究通過顯微觀察技術,對紅色錦鯉體色發生過程進行了詳細觀察,從中篩選出6個體色變化較顯著的時期, 并對這6個時期中MITFa及TYR基因的表達進行熒光定量分析,確定了這兩個色素關鍵基因的表達變化情況。結果顯示MITFa、TYR基因與紅色錦鯉體色形成具有一定相關性,但MITFa和TYR基因在體色發生中的相互作用還有待進一步研究證實。
[1] 張春蘭, 趙文, 徐立蒲, 等. 微生態制劑和著色劑對錦鯉體色的影響[J]. 大連海洋大學學報, 2011, 26(4):328-332.
[2] 崔培, 姜志強, 王雪, 等. 不同蛋白水平的蝦青素飼料對錦鯉體色、生長及免疫的影響[J]. 上海海洋大學學報, 2012, 21(3):382-388.
[3] 王安利, 劉金海, 王維娜. 錦鯉總色素及色素組分的比較研究[J]. 水生生物學報, 2005, 29(6):694-697.
[4] 徐偉, 李池陶, 曹頂臣, 等. 幾種鯉鯽鱗片色素細胞和體色發生的觀察[J]. 水生生物學報, 2007, 31(1):67-72.
[5] 馬雪, 董少杰, 賈艷紅. 五種觀賞魚色素細胞的初步組織學研究[J]. 天津農業科學, 2013, 19(12):70-72.
[6] 易云海, 宋一舸, 楊舟, 等. 魚鰭和鱗片色素組成的比較觀察[J]. 激光生物學報, 2014, 23(1):77-82.
[7] Leclercq E, Taylor J F, Migaud H. Morphological skin colour changes in teleosts[J]. Fish and Fisheries, 2010, 11(2):159-193.
[8] Rawls J F, Mellgren E M, Johnson S L. How the zebrafish gets its stripes[J]. Developmental Biology, 2001, 240(2):301-314.
[9] Johnson S L, Nguyen A N, Lister J A. MITFa is required at multiple stages of melanocyte differentiation but not to establish the melanocyte stem cell[J]. Developmental Biology, 2011, 350(2):405-413.
[10] 王巍, 胡紅霞, 孫向軍, 等. 錦鯉酪氨酸酶基因序列分析及其在不同錦鯉品系不同組織中的表達[J]. 水產學報, 2012, 36(11):1658-1666.
[11] 白瑞, 張美萍, 喬德瑞, 等. MITF 基因與動物毛色的研究進展[J]. 上海畜牧獸醫通訊, 2006(4):2-3.
[12] Liu J, Wen S, Luo C, et al. Involvement of the MITFa gene in the development of pigment cell in Japanese ornamental (Koi) carp (CyprinuscarpioL.)[J]. Genetics and Molecular Research:GMR, 2015, 14(1):2775.
[13] Watson C A, Hill J E, Pouder D B. Species profile: koi and goldfish[R]. Stoneville:Southern Regional Aquaculture Center Publication, 2004.
[14] 王旭霞, 邵力. 觀賞魚體色的研究現狀[J]. 水利漁業, 2008, 28(2):57-59.
[15] 霍鳳敏, 章之蓉, 鄒記興. 中國觀賞魚產業發展概況[J]. 河北漁業, 2010(1):51-53.
[16] 文勝, 劉錦輝, 張永勤, 等. 色素細胞譜系及其發育調控研究進展[J]. 湖南師范大學:自然科學學報, 2014, 37(6):24-28.
[17] 林翔, 周桂炫, 陳菲, 等. 色素細胞發育分化的分子調控機制[J]. 福州大學學報:自然科學版, 2011, 39(5):649-657.
[18] Leclercq E, Taylor J F, Migaud H. Morphological skin colour changes in teleosts[J]. Fish and Fisheries, 2010, 11(2):159-193.
[19] Hultman K A, Johnson S L. Differential contribution of direct-developing and stem cell-derived melanocytes to the zebrafish larval pigment pattern[J]. Developmental Biology, 2010, 337(2):425-431.
[20] Watanabe M, Kondo S. Is pigment patterning in fish skin determined by the Turing mechanism[J]. Trends in Genetics, 2015, 31(2):88-96.
[21] Matthews S A. Observations on pigment migration within the fish melanophore[J]. Journal of Experimental Zoology, 1931, 58(1):471-486.
[22] 楊新鑫, 王成輝, 馬玉清, 等. 甌江彩鯉酪氨酸酶基因的克隆與序列分析[J]. 上海海洋大學學報, 2012, 21(1):14-20.
[23] Parichy D M. Evolution of danio pigment pattern development[J]. Heredity, 2006, 97(3):200-210.
[24] Venizelos A, Benetti D D. Pigment abnormalities in flatfish[J]. Aquaculture, 1999, 176(1/2):181-188.
[25] Guo H, Huang B, Zhang S, et al. Biochemical and histochemical activities of TYRosinase in the skins of normal and albino turbotScophthalmusmaximus[J]. Fish Physiology and Biochemistry, 2003, 29(1): 67-76.
[26] 何志剛, 王冬武, 鄧時銘, 等. 不同飼料對錦鯉體色和著色成本的影響[J]. 中國飼料添加劑, 2015(8): 23-25.
[27] 何志剛, 王冬武, 曾國清,等. 不同飼料對紅白錦鯉生長、體色和血液生化指標的影響[J]. 廣東農業科學, 2015, 42(20):107-111.
[28] Poulaki V, Mitsiades N, Fiskus W. Bromodomain (BRD) inhibition as a novel strategy to inhibit the microphthalmia-associated transcription factor (MITF) axis in uveal melanoma (UM)[J]. Investigative Ophthalmology & Visual Science, 2015, 56(7):5337.
[29] Fang D, Setaluri V. Role of microphthalmia transcription factor in regulation of melanocyte differentiation marker TRP-1[J]. Biochemical and Biophysical Research Communications, 1999, 256(3):657-663.
[30] Darias M J, Andree K B, Boglino A, et al. Coordinated regulation of chromatophore differentiation and melanogenesis during the ontogeny of skin pigmentation ofSoleasenegalensis(Kaup, 1858)[J]. PloS One, 2013, 8(5):e63005.
[31] Schliwa M, Bereiter-Hahn J. Pigment movements in fish melanophores: morphological and physiological studies[J]. Zeitschrift für Zellforschung und Mikroskopische Anatomie, 1973, 147(1):127-148.
[32] Bullara D, De Decker Y. Pigment cell movement is not required for generation of Turing patterns in zebrafish skin[J]. Nature Communications, 2015(6):6971.
[33] Volkening A, Sandstede B. Modelling stripe formation in zebrafish: an agent-based approach[J]. Journal of the Royal Society Interface, 2015, 12(112): 106-114.
ExpressionofMITFaandTYRGeneinBodyColorFormationinRedColorKoiCarpCyprinuscarpioatDifferentStages
TIAN Xue, PANG Xiaolei, WANG Liangyan, GU Liang, GUO Shengqiang, LI Xuejun
( College of Fisheries, Henan Normal University, Xinxiang 453007, China )
This study described the process of the color formation in koi carpCyprinuscarpiowith red color from hatching to whole body color formation. We summarized and concluded the distinct changes in body color during pigmentation development and selected 6 obvious stages of pigmentation ontogenesis, including 1, 2, 3, 4, 12 and 48 dph (day post hatching), respectively. We assessed the expression of the MITFa (Microphthalmia-associtated Transcription Factor) and TYR (TYRosinase) gene in 6 stages by quantitative real-time PCR. The results showed that the expression level of MITFa was the maximum in 1 dph, higher than 48 dph (P<0.05) and significantly higher than other four stages (P<0.01). In addition, there was no significant difference among 2, 3, 4 and 12 dph (P>0.05). Also, the TYR gene was most highly expressed in the 1 dph (P<0.01). And in the 4 dph, TYR expression was significantly higher than the 12 dph and 48 dph, but no difference with 2 dph and 3 dph. There was no significant difference in TYR among the 2, 3, 12 and 48 dph (P>0.05). In conclusion, both MITFa and TYR gene expression trend were identified to decline associated with pigmentation development in koi carp with red color. The expression of MITFa gene was declined firstly and subsequently rised. But the expression trend of TYR gene was fluctuantly changed, which initially exhibited decreased and then reduced again. The findings indicated that the MITFa, and TYR gene had a relationship with the body color formation in koi carp, but the interaction between MITFa and TYR in the color formation remains to be further studied.
Cyprinuscarpio; MITFa; TYR; expression analysis; body color formation
10.16378/j.cnki.1003-1111.2017.02.014
2016-04-08;
2016-06-12.
國家自然科學基金資助項目(31402294);國家級大學生創新創業訓練計劃項目(201410476006);河南省國際科技合作計劃項目(152102410040);河南師范大學博士科研啟動費支持項目(qd13056).
田雪(1981-),女,講師;研究方向:魚類體色調控研究. E-mail: tianxue_81@126.com. 通訊作者:李學軍(1967-),男,教授;研究方向:魚類遺傳育種. E-mail: xjli@htu.cn.
S965.8
A
1003-1111(2017)02-0197-05
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
