魁蚶母源大防御素在子代發育早期的動態變化*
2018-01-12 04:57:24劉志鴻周麗青孫秀俊王澤江楊愛國
海洋與湖沼 2017年2期
吳 彪 劉志鴻 周麗青 孫秀俊 王澤江 楊愛國①
(1. 中國水產科學研究院黃海水產研究所農業部海洋漁業可持續發展重點實驗室 青島 266071;2. 山東省五蓮縣許孟鎮水利站 日照 262315)
魁蚶(Scapharca broughtonii)是一種大型冷溫性蚶類, 廣泛分布于太平洋西部沿岸, 因其個體大、肉味鮮美, 并富含蛋白質和多種維生素, 在國內外市場深受喜愛, 是我國黃、渤海區的重要經濟貝類(吳彪等,2012)。最近幾年, 由于消費市場及增殖放流活動對魁蚶的需求量激增, 帶動了苗種繁育產業蓬勃發展。魁蚶營體外受精、體外發育的生殖方式, 卵子排出體外后的受精、發育等過程完全暴露于水環境中。處在開放水體中的胚胎或者幼體非常容易受到病原微生物的侵染, 導致胚胎孵化率下降、幼體生長緩慢, 甚至死亡, 嚴重影響人工繁育魁蚶的幼苗出庫率和苗種質量。胚胎發育早期, 個體自身免疫系統尚未形成或發育不完善, 除受精膜第一道免疫防線外, 多數水產動物能夠通過卵子從母體獲得免疫因子而具備一定的免疫力, 即母源性免疫。因此, 母源免疫因子對胚胎早期的正常發育具有重要的免疫保護作用。
母源免疫研究已在哺乳類、鳥類、爬行類等多種動物中廣泛開展(Bandrick et al, 2014; Blanco et al,2015), 但在水產動物中的相關研究起步較晚, 已有的報道主要集中在魚、蝦等少數物種(Huang et al,1999; Swain et al, 2009)。在高等的脊椎動物中, 通過免疫母體來增強子代免疫力已經得到廣泛應用, 如用黏液囊病病毒疫苗免疫母雞, 其子代卵黃囊中有不同水平的黏液囊病病毒抗體存在, 能夠對子代早期發育發揮免疫保護作用(Grindstaff et al, 2006)。目前高等動物的相關研究多關注于抗體的傳遞和作用,但是關于水產動物的一些研究已經證明, 除抗體外,補體、溶菌酶、凝集素等非特異性免疫因子也能夠通過卵子傳遞給子代, 發揮重要的免疫功能(Yousif et al, 1991; Olafsen, 1996; L?voll et al, 2006; Wang et al,2009), 尤其是對主要依靠非特異性免疫系統進行機體免疫反應的軟體動物更重要。目前關于貝類的研究報道較少, 母源性免疫因子的轉移特點及在子代早期的變化規律尚不清楚, 明確該問題是開發利用母源免疫的前提和基礎, 對提高貝類胚胎和幼蟲早期免疫力具有重要的理論意義和實用價值。
大防御素(big-defensin), 是一種富含半胱氨酸的抗菌肽, 最先在鱉(Saito et al, 1995)中發現, 后來在菲律賓蛤仔(Ruditapes philippinesis)(Wei et al, 2003;Zhao et al, 2010)、海灣扇貝(Argopecten irradians)(Zhao et al, 2007)、蝦夷扇貝(Patinopecten yesoensis)(于赫男, 2012)、長牡蠣(Crassostrea gigas)(Rosa et al,2015)、三角帆蚌(Hyriopsis cumingii)(Wang et al, 2014)等多種貝類中也被證實存在, 且具有廣譜的抗革蘭氏陰性菌、革蘭氏陽性菌及真菌的生物活性, 發揮重要的免疫保護作用。Li等(2012)克隆獲得了魁蚶大防御素基因cDNA序列, 為本研究的開展奠定了前期基礎。本研究擬通過實時熒光定量 PCR(quantitative real-time PCR, qRT-PCR)和酶聯免疫吸附測定(enzyme linked immunosorbent assay, ELISA)技術從mRNA和蛋白質水平上檢測魁蚶胚胎及幼蟲發育早期大防御素的表達變化, 以明確大防御素從母體向子代傳遞和表達規律。
1 材料與方法
1.1 魁蚶苗種繁育及樣品收集
實驗用的魁蚶親貝取自山東省長島縣海區, 在親本自然繁育期前一個月從海區捕獲后轉移至育苗車間升溫促熟。性腺成熟后, 利用升溫過濾海水刺激以誘導親本產卵、排精, 建立兩個同父異母半同胞家系。同一母本的卵細胞在受精后即刻被平均分成兩組,一組在22°C的充氣海水中孵化、培育, 為對照組; 另一組則在含有5×108CFU/L鰻弧菌(Vibrio anguillarum)的同條件海水中培育, 為鰻弧菌脅迫處理組。培育期間, 以投喂金藻為主, 配合小球藻等其他單胞藻, 日換水2—3次。
實驗樣品的收集時間分別為: 卵細胞期、受精卵期、多細胞期、囊胚期、擔輪幼蟲前期、擔輪幼蟲后期、D形幼蟲期和殼頂幼蟲期。用500目或300目篩絹在上述各幼體發育時期隨機收集實驗材料。材料固定方法如下, RNA提取材料: 加入Trizol后, 液氮速凍后保存于–80°C; 蛋白質提取材料: 樣品先置于預冷的無菌Tris-HCl緩沖液中(pH8.0), 4°C下超聲波細胞破碎儀將樣品充分破碎, 6000×g離心30min后, 取上清液氮速凍, 保存于–80°C。
1.2 RNA的提取及qRT-PCR檢測
總RNA的提取參考Zheng等(2015)的方法, 并稍作修改。樣品去除Trizol后, 分別經過Solution D、氯仿/異戊醇、β-巰基乙醇充分裂解, 水飽和酚/氯仿/異戊醇兩次抽提, 異丙醇/醋酸鈉沉淀、75%乙醇兩次洗滌后, RNA溶于RNA-free水中, 并加入DNaseⅠ去除基因組DNA。運用超微量分光光度計(A260/A280值)和瓊脂糖電泳檢測RNA濃度及完整性。根據試劑盒PrimeScript? RT reagent Kit(TaKaRa)說明書合成cDNA第一條鏈用于qRT-PCR實驗。
以上各時間點樣品中大防御素 mRNA表達水平通過 qRT-PCR進行檢測, 具體操作方法參照 Wu等(2015)的方法進行。根據已有的魁蚶大防御素基因的cDNA序列(GenBank登錄號: JQ782659), 用 Primer Premier 5.0設計 qRT-PCR 引物(序列為 F:ACCTTTTGTTGTCCACG, R: TGTTCTACACCACCC TC), 選用魁蚶 β-actin基因作為內參基因(引物序列為F: GGTTACACTTTCACCACCACAG, R: ACCGG AAGTTTCCATACCTAAGA)。qRT-PCR在儀器LineGene 9600(BIOER)上進行, 反應體系為25μL, 具體包括: ExTaq12.5μL、正反向引物(10μmol/L)各0.5μL、cDNA模板2μL、滅菌蒸餾水9.5μL。具體反應程序為: 94°C預變性 2min, 94°C變性30s, 62°C退火30s, 72°C延伸30s, 40個循環。內參基因與目的基因分管同時進行, 每個樣品重復3次, 利用儀器軟件計算各樣品 Ct值, 數據取平均值。將卵細胞時期mRNA含量作為參照1, 采用2–Ct△△法對基因表達量進行相對定量分析(Livak et al, 2001)。運用SPSS軟件, 采用t檢驗法檢測每個相同發育時期處理組與正常組之間的差異性, 單因素方差分析不同發育時期表達量差異, 差異顯著水平設為P<0.05, 極顯著水平為P<0.01。
1.3 魁蚶大防御素蛋白水平的ELISA檢測
魁蚶大防御素抗體由艾比瑪特生物醫藥有限公司提供, 利用抗原蛋白免疫健康兔子七次后純化獲得。不同發育時期大防御素蛋白含量通過 ELISA檢測, 具體操作參照劉帥帥等(2013)的方法進行。主要過程為: 運用 BCA法測定每份樣品總蛋白濃度并調整至濃度一致后, 取200μL樣品加入酶標板, 4°C過夜后棄去殘液, 之后用PBST洗滌3次; 用300μL10%的脫脂奶粉于 37°C中孵育 2h, 去掉封閉液并用PBST洗板3次; 加入1:1000稀釋的魁蚶大防御素抗體 100μL, 37°C 孵育 2h, PBST洗滌 3次; 加入 1∶2000稀釋的辣根過氧化物酶標記的羊抗兔二抗100μL, 37°C 孵育 2h, PBST 洗滌 4次; 加入 100μL 辣根酶底物 TMB溶液, 避光條件下 37°C條件下反應10min后, 加入2mol/L的硫酸50μL以終止反應。用PBS溶液代替樣品作為陰性對照, 其余條件與上述相同。每個樣品設置三個平行, 酶標儀讀取OD450吸光值。以制備抗體時所用的蛋白抗原為標準品制作標準曲線和曲線方程, 通過方程用OD值計算各樣品中大防御素蛋白的濃度。將卵細胞時期大防御素蛋白含量視為參照 1, 計算每個發育時期蛋白含量的相對值,繪制蛋白表達變化趨勢圖。差異顯著性檢驗與上述qRT-PCR差異分析方法相同。
2 結果
2.1 mRNA水平的動態變化
魁蚶大防御素 mRNA在兩個魁蚶家系的幼蟲發育早期表達變化如圖1所示。從圖1中可以看出, F1、F2兩家系的對照組和脅迫處理組 mRNA的變化趨勢基本相同, 即先降后升, 但家系之間、以及同家系對照組和處理組之間在變化幅度和時間上有一定差別。結合表 1的差異顯著性分析結果發現, 在 F1家系中,大防御素在卵細胞受精后開始降低, 對照組和處理組分別在囊胚期和多細胞期達到最低值, 分別為卵細胞期的0.27倍和0.23倍, 之后開始上調表達; 正常組中, 多細胞期、囊胚期的表達量極顯著低于卵細胞期(P<0.01), 經多細胞期的最低值后上調, 至 D形幼蟲期達到最高值, 差異達到顯著水平(P<0.05); 而脅迫處理組, 除了受精卵期, 其余各發育期的表達量與卵細胞期相比均達到了極顯著差異, 經囊胚期最低值后開始不斷升高, 殼頂幼蟲期達到卵細胞期的 3.8倍; 對照組與處理組相比, 對照組在多細胞期極顯著高于處理組, 在殼頂幼蟲期極顯著低于處理組, 囊胚期之后的各階段, 處理組高于正常組, 但差異不顯著。F2家系與F1表現出基本相同的變化趨勢, 也是先下降至最低, 之后擔輪幼蟲前期開始顯著上調; 擔輪幼蟲期至殼頂幼蟲期, 正常組和處理組的表達量與卵細胞期的表達均達到了顯著或極顯著水平; 兩組之間, 受精卵期、囊胚期、殼頂幼蟲期具有顯著差異。
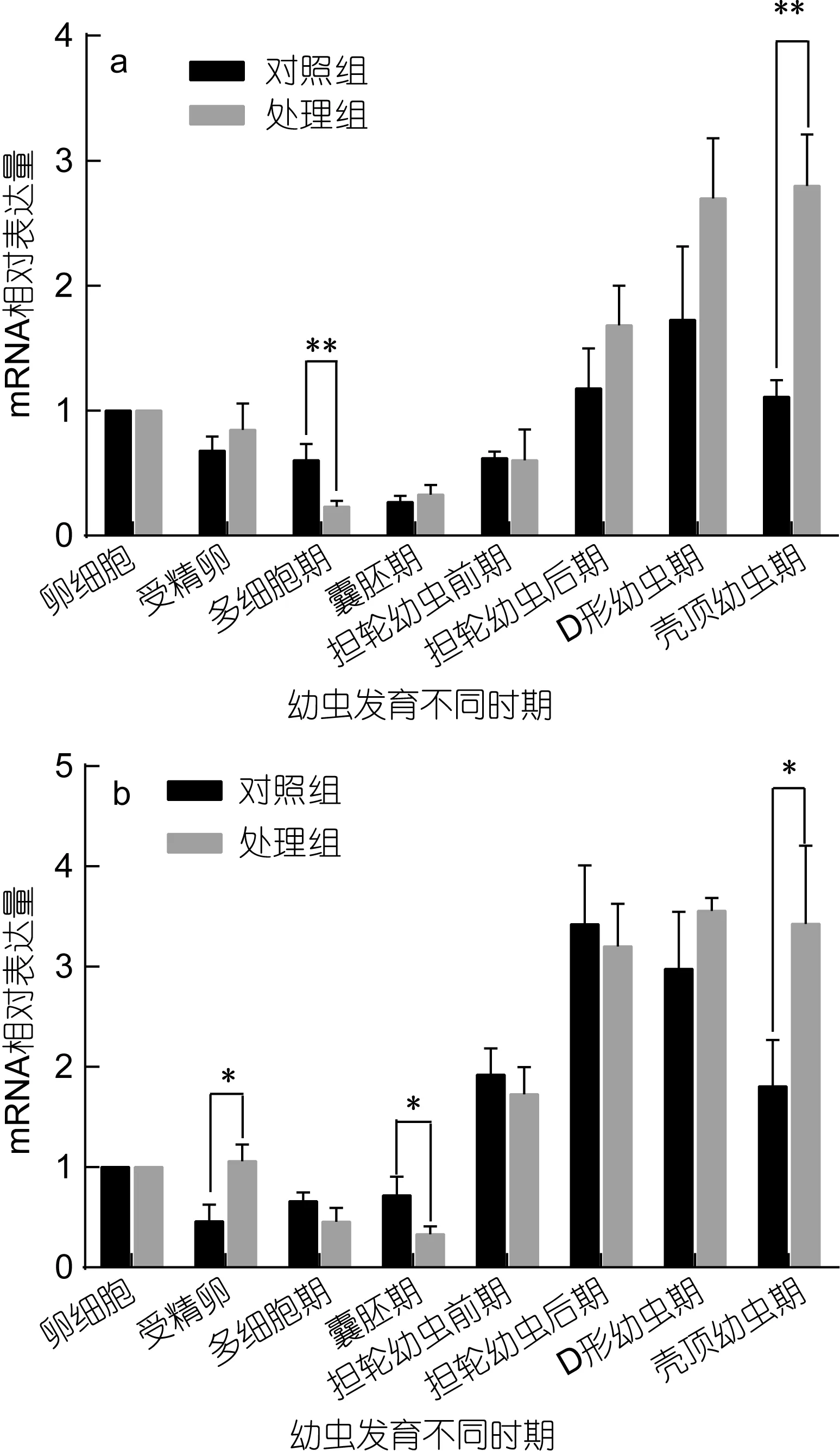
圖1 魁蚶大防御素mRNA在魁蚶幼蟲不同發育時期的變化Fig.1 The big-defensin mRNA during the early-larvae developmental stage of S. broughtonii
2.2 蛋白水平的動態變化
大防御素蛋白水平的表達變化如圖2所示, 顯著性檢驗結果見表 2。結果表明, 大防御素蛋白在魁蚶幼體早期發育階段與 mRNA的變化趨勢基本相似,同樣表現為先下降后升高, 而且在升高階段, 處理組表達量普遍高于正常組。F1家系中, 正常組蛋白從受精后開始下降, 多細胞期、囊胚期下降至極顯著水平,之后開始逐漸升高, 但均未達到顯著水平; 而處理組,多細胞期、囊胚期、擔輪幼蟲前期、殼頂幼蟲期與卵細胞期有極顯著差異, D形幼蟲期則具有顯著差異;兩組之間, 處理組從囊胚期之后高于對照組, 但只有囊胚期和擔輪幼蟲前期時差異達到顯著(P<0.05)。F2家系趨勢與F1基本相同, 囊胚期前, 處理組低于正常組; 擔輪幼蟲前期之后, 處理組高于對照組, 其中擔輪幼蟲前期、擔輪幼蟲后期和殼頂幼蟲期的差異達到顯著水平(P<0.05)。
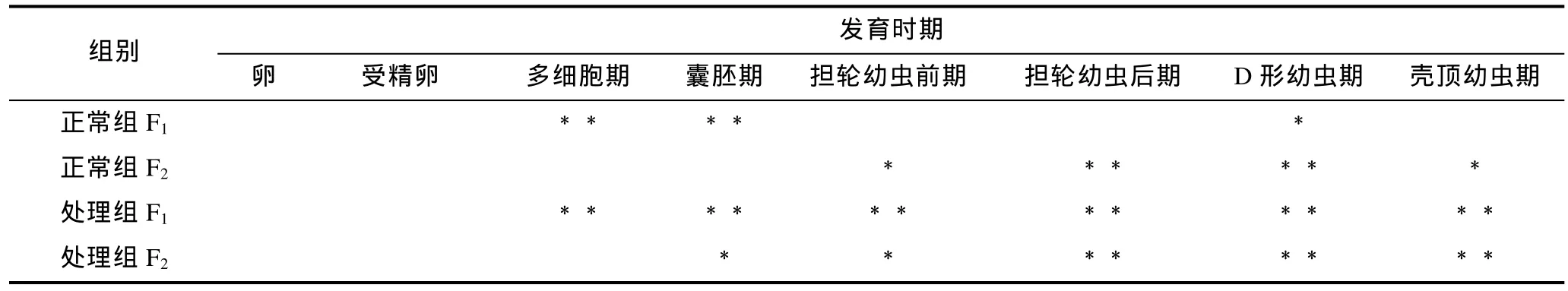
表1 魁蚶大防御素mRNA變化的顯著性檢驗Tab.1 Significance test on big-defensin mRNA
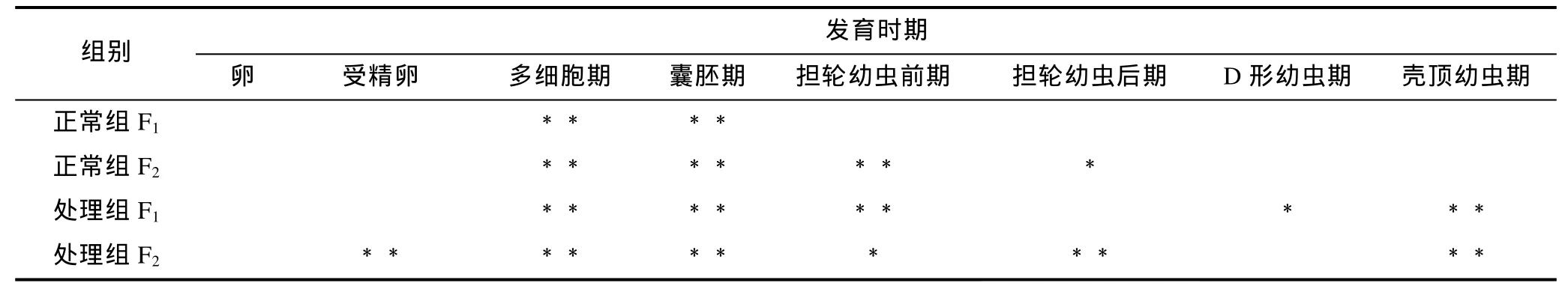
表2 魁蚶大防御素蛋白變化的顯著性檢驗Tab.2 Significance test on big-defensin protein
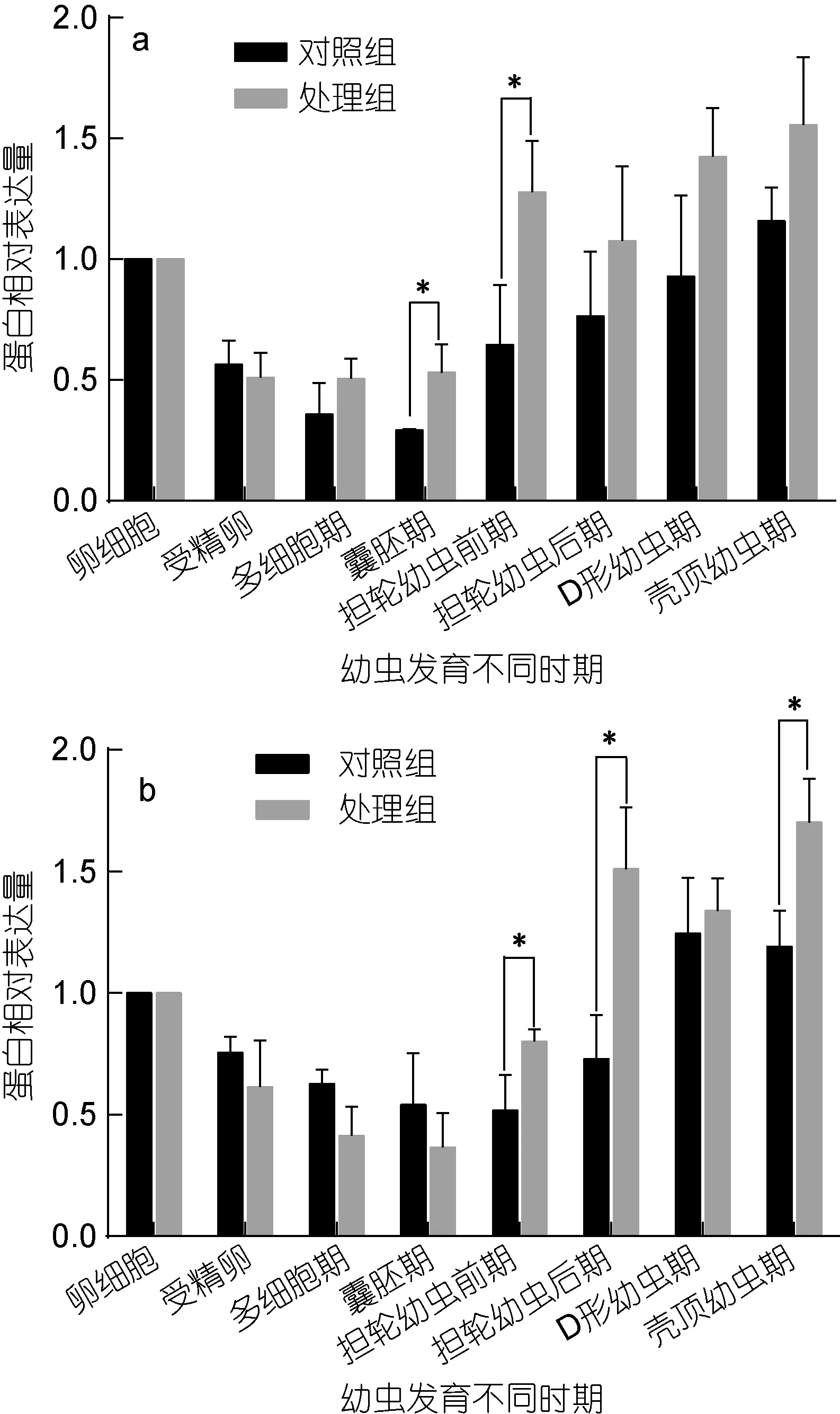
圖2 魁蚶大防御素蛋白在魁蚶幼蟲不同發育時期的動態變化Fig.2 The big-defensin protein during the early-larvae developmental stage of S. broughtonii
3 討論
傳代免疫效應是母體免疫力通過母源性物質傳遞給后代, 從而使子代擁有一定的免疫能力。母體產生的一些抗體以及凝集素、溶菌酶等先天性免疫因子從母體向子代傳遞, 并發揮免疫保護作用, 已經在很多動物中得到證實(張士璀等, 2007; Wanget al, 2009,2010, 2012)。與脊椎動物研究相比, 目前有關無脊椎動物母源免疫方面的研究報道較少。不過, 也有研究證明一些頭索動物、軟體動物也同樣存在這種被動的母體免疫。如, 刁明月(2015)在文昌魚(Branchiostoma lanceolatum)1—2細胞期的受精卵液中發現了以蛋白和mRNA存在的α2巨球蛋白, 并證實這種母源性蛋白發揮了重要的抑菌作用; 岳峰(2013)在櫛孔扇貝(Chlamys farreri)卵細胞中發現有 CfCu/Zn-SOD、CfLBP/BPI、CfLBP、CfLec-3和CfLYZ的mRNA和蛋白表達。本研究在魁蚶卵細胞期檢測到了大防御素mRNA和蛋白, 這與上述的研究結果相一致, 進一步證實了無脊椎動物先天免疫因子的母源傳遞現象。
認識免疫因子在子代的傳遞表達規律是母源免疫的重要研究內容, 也是開發利用母源免疫的前提和基礎。通過對兩個魁蚶家系幼蟲不同發育階段的檢測, 本研究發現大防御素 mRNA和蛋白的表達規律基本一致, 即自受精后開始持續下降, 一般至囊胚期都維持在比較低的水平, 從擔輪幼蟲期開始, 含量開始迅速上升, 這與已有的研究報道結果相似。大西洋鮭(Salmo salar)卵細胞中的抗體在受精后開始下降,經過孵化期、仔魚期, 直至幼魚自身合成抗體, 水平開始迅速上升(Olsenet al, 1997); 王鴻淼(2012)運用Western檢測發現斑馬魚(Danio rerio)母源性 IgM從受精后開始減少, 至第5天檢測不到抗體。這些結果說明, 母源性的免疫因子在卵細胞受精后開始被逐步消耗, 含量不斷下降, 直至幼體開始自身合成, 含量開始升高。本研究中, 魁蚶大防御素含量基本在擔輪幼蟲期開始升高, 而且 mRNA的變化幅度比蛋白水平更大, 這表明此時的魁蚶幼蟲可能已經開始自身合成大防御素。岳峰(2013)發現櫛孔扇貝模式識別受體(pattern recognition receptor, PRR)分子CfPGRP-S1、CfLGBP、CfLec-1和CfLec-3的 mRNA在擔輪幼蟲期開始大量表達, 利用整體免疫熒光定位進一步證實了其表達部位最早是出現在擔輪幼蟲基部的兩側對稱細胞中, 認為此時是扇貝免疫系統的最早形成期。這可以解釋本研究中大防御素含量為什么在擔輪幼蟲期開始升高, 同時也說明魁蚶的免疫系統也可能在此時形成, 當然, 這還需要更多的證據來證明。
免疫刺激能夠增強免疫因子的表達, 這在許多物種中已有報道。如, 鰻弧菌刺激能夠增強蝦夷扇貝(Patinopecten yessoensis)卵黃蛋白原(Wu et al,2015)、魁蚶鐵蛋白(Zheng et al, 2016)及半乳糖凝集素(鄭利兵等, 2015)這些免疫因子的表達等。本研究中, 擔輪幼蟲之前, 處理組與對照組大防御素的表達量基本相當, 互有高低; 而之后的時期, 處理組表達量基本高于對照組, 但差異未全部都達到顯著水平。這與魁蚶成體受到鰻弧菌刺激后的響應情況有差別。Li等(2012)研究了鰻弧菌刺激魁蚶成體后,血細胞和肝胰腺中大防御素mRNA的動態變化, 結果表明, 刺激 8h后血細胞中的表達量顯著升高, 至16h時上調至最高值; 肝胰腺中的表達量在24h達到最高值。而且, 最高值分別是對照組的 10.25倍和5.14倍。可見, 擔輪幼蟲期后的幼體對鰻弧菌刺激的響應與成體相似, 受刺激后大防御素均上調表達,但是響應程度不如成體。這可能是由于幼體免疫系統發育并不完善, 再加上鰻弧菌沒有進行滅活而具有致病性, 可能會使得部分幼蟲機體發育受阻而影響其免疫活性因子的合成。岳峰(2013)的研究表明,用滅活的鰻弧菌對櫛孔扇貝親貝進行免疫刺激后,許多免疫分子的蛋白表達在子代卵細胞、胚胎或幼蟲中均明顯升高, 且免疫刺激組的抗菌活力顯著高于對照。這給我們新的提示, 在充分了解先天免疫因子在子代表達規律前提下, 結合母源免疫開發提高母源免疫因子含量的方法, 能夠有效增強子代的免疫抵抗力, 可以為科學制定提高子代早期成活率的策略提供理論依據, 對魁蚶苗種繁育產業發展具有重要的現實意義。
4 結論
本文通過qRT-PCR和ELISA檢測了魁蚶胚胎及幼體早期發育過程中大防御素在 mRNA水平和蛋白質水平上的動態變化, 發現母源性大防御素能夠通過卵細胞傳遞給子代, 并在卵子受精后被逐漸消耗。幼體發育至擔輪幼蟲期可能具備了自身合成大防御素的能力, 使幼體體內大防御素含量增加, 并且鰻弧菌刺激能夠促進其增加幅度。
刁明月, 2015. α2巨球蛋白在文昌魚受精卵中的存在及其抑菌作用分析——頭索動物母源性免疫新證據. 青島: 中國海洋大學碩士論文, 14—51
于赫男, 2012. 蝦夷扇貝大防御素基因的克隆、表達及G—型溶菌酶基因的啟動子分析. 大連: 遼寧師范大學碩士學位論文, 14—23
王鴻淼, 2012. 斑馬魚(Danio rerio)母源性抗體的傳遞及其對子代的保護作用. 青島: 中國海洋大學碩士論文, 23—54
劉帥帥, 姚 琳, 馬麗萍等, 2013. 貝類中 3種組織血型抗原ELISA檢測方法的建立與分型. 中國水產科學, 20(1): 211—216
吳 彪, 梁 超, 楊愛國等, 2012. 基于 SSR 標記的魁蚶(Scapharca broughtonii)不同群體遺傳結構的分析. 海洋與湖沼, 43(4): 863—869
張士璀, 李 欣, 汲廣東, 2007. 魚類免疫系統的早期發生.中國海洋大學學報, 37(4): 557—562
岳 峰, 2013. 櫛孔扇貝免疫系統發生及母源免疫的初步研究.青島: 中國科學院研究生院(海洋研究所)博士學位論文,61—104
鄭利兵, 吳 彪, 劉志鴻等, 2015. 魁蚶(Scapharca broughtonii)半乳糖凝集素(SbGal)基因 cDNA的克隆及表達分析. 海洋與湖沼, 46(5): 1061—1070
Bandrick M, Theis K, Molitor T W, 2014. Maternal immunity enhances Mycoplasma hyopneumoniaevaccination induced cell-mediated immune responses in piglets. BMC Veterinary Research, 10(1): 124
Blanco J C G, Pletneva L M, Oue R O et al, 2015. Maternal transfer of RSV immunity in cotton rats vaccinated during pregnancy. Vaccine, 33(41): 5371—5379
Grindstaff J L, Hasselquist D, Nilsson J ? et al, 2006.Transgenerational priming of immunity: maternal exposure to a bacterial antigen enhances offspring humoral immunity.Proceedings of the Royal Society B: Biological Sciences,273(1600): 2551—2557
Huang C C, Song Y L, 1999. Maternal transmission of immunity to white spot syndrome associated virus (WSSV) in shrimp(Penaeus monodon). Developmental & Comparative Immunology, 23(7—8): 545—552
Li M, Zhu L, Zhou C Y et al, 2012. Molecular characterization and expression of a novel big defensin (Sb-BDef1) from ark shell, Scapharca broughtonii. Fish & Shellfish Immunology,33(5): 1167—1173
Livak K J, Schmittgen T D, 2001. Analysis of relative gene expression data using real-time quantitative PCR and the 2–CT△△Method. Methods, 25(4): 402—408
L?voll M, Kilvik T, Boshra H et al, 2006. Maternal transfer of complement components C3-1, C3-3, C3-4, C4, C5, C7, Bf,and Df to offspring in rainbow trout (Oncorhynchus mykiss).Immunogenetics, 58(2—3): 168—179
Olafsen J A, 1996. Lectins: models of natural and induced molecules in invertebrates. In: Cooper E L ed. Invertebrate Immune Responses. Berlin Heidelberg, Germany: Springer,49—76
Olsen Y A, Press C M, 1997. Degradation kinetics of immunoglobulin in the egg, alevin and fry of Atlantic salmon, Salmo salar L., and the localisation of immunoglobulin in the egg. Fish & Shellfish Immunology,7(2): 81—91
Rosa R D, Alonso P, Santini A et al, 2015. High polymorphism in big defensin gene expression reveals presence–absence gene variability (PAV) in the oyster Crassostrea gigas.Developmental & Comparative Immunology, 49(2):231—238
Saito T, Kawabata S, Shigenaga T et al, 1995. A novel big defensin identified in horseshoe crab hemocytes: isolation,amino acid sequence, and antibacterial activity. Journal of Biochemistry, 117(5): 1131—1137
Swain P, Nayak S K, 2009. Role of maternally derived immunity in fish. Fish & Shellfish Immunology, 27(2): 89—99
Wang H M, Ji D R, Shao J Z et al, 2012. Maternal transfer and protective role of antibodies in zebrafish Danio rerio.Molecular Immunology, 51(3—4): 332—336
Wang G L, Xia X L, Li X L et al, 2014. Molecular characterization and expression patterns of the big defensin gene in freshwater mussel (Hyriopsis cumingii). Genetics and Molecular Research, 13(1): 704—715
Wang Z P, Zhang S C, 2010. The role of lysozyme and complement in the antibacterial activity of zebrafish (Danio rerio) egg cytosol. Fish & Shellfish Immunology, 29(5):773—777
Wang Z P, Zhang S C, Tong Z et al, 2009. Maternal transfer and protective role of the alternative complement components in zebrafish Danio rerio. PLoS One, 4(2): e4498
Wei Y X, Guo D S, Li R G et al, 2003. Purification of a big defensin from Ruditapes philippinesis and its antibacterial activity. Acta Biochimica Et Biophysica Sinica, 35(12):1145—1148
Wu B, Liu Z H, Zhou L Q et al, 2015. Molecular cloning,expression, purification and characterization of vitellogenin in scallop Patinopecten yessoensis with special emphasis on its antibacterial activity. Developmental & Comparative Immunology, 49(2): 249—258
Yousif A N, Albright L J, Evelyn T P T, 1991. Occurrence of lysozyme in the eggs of coho salmon Oncorhynchus kisutch.Diseases of Aquatic Organisms, 10(1): 45—49
Zhao J M, Li C H, Chen A Q et al, 2010. Molecular characterization of a novel big defensin from clam Venerupis philippinarum. PLoS One, 5(10): 13480
Zhao J M, Song L S, Li C H et al, 2007. Molecular cloning,expression of a big defensin gene from bay scallop Argopecten irradians and the antimicrobial activity of its recombinant protein. Molecular Immunology, 44(4):360—368
Zheng L B, Liu Z H, Wu B et al, 2016. Ferritin has an important immune function in the ark shell Scapharca broughtonii.Developmental & Comparative Immunology, 59: 15—24
Zheng L B, Wu B, Liu Z H et al, 2015. A manganese superoxide dismutase (MnSOD) from ark shell, Scapharca broughtonii:molecular characterization, expression and immune activity analysis. Fish & Shellfish Immunology, 45(2): 656—665
