燈盞花乙素對J774A.1巨噬細胞中ATP誘導的炎癥小體活化和細胞焦亡的影響
2018-01-24 06:25:50景艷蕓李陳廣徐麗慧白文靜歐陽東云何賢輝
中國藥理學通報 2018年2期
關鍵詞:小鼠
景艷蕓,李陳廣,顏 亮,徐麗慧,白文靜,歐陽東云,何賢輝
(暨南大學生命科學技術學院 1. 免疫生物學系、2. 細胞生物學系,廣東 廣州 510632)
炎癥小體是存在于免疫細胞中的多亞基蛋白質復合體,可以感應入侵的病原體或損傷組織釋放的危險信號,在協助機體抵御病原體感染和組織損傷中發揮重要作用[1-2]。NLRP3(NOD-like receptor family,pyrin domain-containing 3)炎癥小體是目前研究最廣泛的炎癥小體之一,其活化一般需要兩個信號,第一信號是病原體相關分子模式(pathogen-associated molecular pattern,PAMP),如脂多糖(lipopolysaccharide, LPS)被細胞膜模式識別受體(pattern recognition receptor,PRR)識別并結合,激活NF-κB信號通路,誘導NLRP3、白細胞介素-1β前體(pro-interleukin-1β,pro-IL-1β)等炎癥性相關蛋白的表達;第二信號是損傷相關分子模式 (danger-associated molecular pattern,DAMP)如三磷酸腺苷(ATP)等激活NLRP3,通過接頭蛋白ASC(apoptosis-associated speck-like protein containing a CARD),募集半胱天冬氨酸蛋白酶-1前體(pro-cysteinyl aspartate specific protease-1,pro-caspase-1),進而自我催化加工形成活化的caspase-1;活化的caspase-1切割pro-IL-1β形成成熟的IL-1β并分泌至細胞外,引起炎癥反應[1, 3]。炎癥小體活化會誘導細胞發生炎癥性死亡——細胞焦亡(pyroptosis)。細胞焦亡是近年來新發現的一種不同于細胞凋亡與細胞壞死的新型細胞死亡方式,它依賴于caspase-1活化,并通過切割gasdermin D (GSDMD)產生其N端產物(GSDMD-NT),在細胞膜上成孔,使炎癥因子如成熟IL-1β、高速泳動族框蛋白1(high-mobility group box 1,HMGB1)等分泌至胞外,導致炎癥反應[4]。因此,正常的NLRP3炎癥小體的活化是機體固有免疫防線的重要組成部分,而NLRP3炎癥小體的異常活化則與多種炎癥性疾病的發生密切相關,是治療相關疾病的重要藥物靶標[5-6]。
燈盞花乙素(scutellarin)又名野黃芩苷,是從燈盞花等中草藥中分離提取的一種黃酮類物質,具有多種藥理學作用,如抗炎、抗凋亡、抗腫瘤等[7]。在臨床上用于缺血和缺氧性心腦血管疾病的治療以及腦血管病后癱瘓的治療[8]。研究顯示,燈盞花乙素可以通過抑制鈣離子調節的鈣調磷酸酶和鈣蛋白激酶的信號通路,改善心腦血管疾病[9]。燈盞花乙素也具有明顯的抗炎作用,能抑制炎癥相關信號通路和炎癥性細胞因子的表達,從而減輕炎癥相關疾病的癥狀[10]。缺血性心腦血管疾病涉及組織損傷及損傷相關炎癥反應,可能原因之一是損傷引起的內源性危險信號(如ATP等)釋放到細胞外,激活固有免疫細胞中NLRP3活化和IL-1β等炎癥性細胞因子的分泌[11]。但目前燈盞花乙素對NLRP3炎癥小體活化的影響尚未見報道。在本研究中,我們以LPS致敏的小鼠巨噬細胞 J774A.1作為炎癥細胞模型,探討燈盞花乙素對NLRP3炎癥小體活化和細胞焦亡的影響,為進一步了解其藥理學機制提供實驗依據。
1 材料
1.1細胞與試劑小鼠巨噬細胞系J774A.1細胞購自中國科學院昆明動物研究所細胞庫。燈盞花乙素購于廣東省藥品檢驗所;ATP、碘化丙錠(propidium iodide,PI)、Hoechst 33342、LPS、MDL12330A,購于Sigma-Aldrich公司;DMEM、Opti-MEM、胎牛血清(fetal bovine serum,FBS)、青霉素、鏈霉素為Thermo Fisher公司產品;抗caspase-1抗體購自Santa Cruz Biotechnology公司;抗NLRP3抗體購于Adipogen AG公司;抗IL-1β、HMGB1、ASC、β-tubulin、phospho-(Ser/Thr) PKA substrate、辣根過氧化物酶標記二抗等抗體以及Protein G agarose beads購于Cell Signaling Technology公司;H89、PMSF購自碧云天公司;Cytometric Bead Array (CBA) Mouse IL-1β Flex Set試劑盒為BD公司的產品。
1.2儀器Labofuge 400R低溫高速離心機(美國Thermo Scientifc公司);FACSCalibur流式細胞儀(美國BD公司);Axio Observer D1倒置熒光顯微鏡(德國Zeiss公司);MINI Protean 2電泳系統(美國 Bio-Rad公司);FluorChem 8000凝膠圖像成像儀(美國AlphaInnotech公司)。
2 方法
2.1細胞培養小鼠J774A.1細胞為貼壁型細胞,培養于完全培養基(含10% FBS、100 kU·L-1青霉素、100 mg·L-1鏈霉素的DMEM)中,置37℃、5% CO2培養箱中培養,取對數生長期細胞進行實驗。
2.2細胞焦亡的檢測參考相關文獻[12],J774A.1細胞接種于24孔板,培養過夜后,用0.5 mg·L-1LPS刺激4 h,加入燈盞花乙素(0.1、0.2、0.4 mmol·L-1)作用1 h,補加4 mmol·L-1的ATP反應1 h,最后加入PI(2 mg·L-1)和Hoechst 33342(5 mg·L-1)室溫下孵育染色10 min,倒置熒光顯微鏡觀察并拍照,統計PI陽性細胞占總細胞(即Hoechst陽性)的百分比。
2.3免疫印跡分析將J774A.1細胞接種于6孔板,培養過夜后,用0.5 mg·L-1LPS刺激4 h,加入燈盞花乙素(0.1、0.2、0.4 mmol·L-1)作用1 h,補加4 mmol·L-1的ATP反應1 h。分別提取細胞和上清中(樣品的體積相同)的蛋白,進行SDS-PAGE電泳,電泳結束后,將分離的蛋白質轉至PVDF膜上。取出PVDF膜,加入適量的封閉液(含3% FBS和0.1% 吐溫20的PBST),常溫下封閉1 h,然后加入相應比例的一抗,4℃搖床孵育過夜。加入辣根過氧化物酶標記的二抗,室溫搖床孵育1 h。加入ECL化學發光液顯色,成像儀拍照記錄結果,最后用Alpha EaseFC 4.0軟件(Alpha Innotech)分析條帶的灰度值。
2.4可溶性細胞因子IL-1β的檢測小鼠J774A.1細胞接種于24孔板,培養過夜后,用0.5 mg·L-1LPS刺激4 h,加入燈盞花乙素(0.1、0.2、0.4 mmol·L-1)作用1 h,補加4 mmol·L-1的ATP反應1 h。最后收集細胞培養上清液,按照試劑盒說明書定量檢測上清中IL-1β含量。用流式細胞儀檢測,通過CellQuest軟件(BD公司)進行數據獲取與分析。
2.5cAMP-PKA信號通路的抑制小鼠J774A.1細胞接種于24孔板,培養過夜后,用0.5 mg·L-1LPS刺激4 h,加入燈盞花乙素(0.1、0.2、0.4 mmol·L-1)作用30 min后,補加腺苷酸環化酶特異性抑制劑MDL12330A 10 μmol·L-1作用30 min,或者加入PKA的選擇性抑制劑H89 20 μmol·L-1作用30 min后,加燈盞花乙素(0.1、0.2、0.4 mmol·L-1)作用1 h,最后加入4 mmol·L-1的ATP反應1 h。
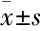
3 結果
3.1燈盞花乙素抑制ATP誘導的NLRP3炎癥小體活化ATP是NLRP3炎癥小體的常用活化劑,而活化的caspase-1(10 ku)與成熟IL-1β(17 ku)是NLRP3炎癥小體活化的兩個重要標志。免疫印跡檢測顯示,單獨LPS處理能夠促進pro-IL-1β(31 ku)與NLRP3的表達,而對組成型表達的pro-caspase-1(45 ku)與ASC無影響(Fig 1A);單獨LPS處理的巨噬細胞的培養上清液中未檢測到活化的caspase-1p10與成熟的IL-1β(17 ku)的釋放。經第二信號ATP刺激后,細胞培養液上清中能夠明顯檢測到活化的caspase-1p10以及成熟IL-1β(17 ku)(Fig 1A-1C)。而燈盞花乙素預處理能夠劑量依賴性地抑制ATP誘導的caspase-1活化以及IL-1β的成熟與分泌(Fig 1A-1C);同時,LPS+燈盞花乙素處理并不會促進caspase-1活化以及IL-1β的成熟與分泌(Fig 1A-1C)。另外,用 CBA檢測細胞培養液上清中IL-1β的分泌水平,顯示燈盞花乙素能夠劑量依賴性地抑制ATP誘導的IL-1β的釋放(Fig 1D),這與免疫印跡法的檢測結果相一致。這些結果表明,燈盞花乙素能夠抑制ATP誘導的小鼠巨噬細胞中NLRP3炎癥小體的活化。
3.2燈盞花乙素抑制巨噬細胞中ATP誘導的細胞焦亡NLRP3炎癥小體活化會導致細胞焦亡的發生。而細胞焦亡可以通過PI染色法以及標志性蛋白HMGB1的釋放進行分析。如Fig 2所示,當加入第二信號ATP后,細胞明顯變圓腫脹,甚至破裂成細胞碎片,從而使PI染料穿過胞膜進入細胞核內,發出紅色熒光,表明ATP誘導LPS致敏的巨噬細胞發生了焦亡,其焦亡率約為45%(Fig 2A、2B)。而燈盞花乙素預處理能夠明顯抑制ATP誘導的細胞焦亡(Fig 2A、2B);單獨LPS或LPS+燈盞花乙素聯合作用并不會誘導細胞焦亡的發生。免疫印跡法分析顯示,燈盞花乙素能夠劑量依賴性地抑制ATP誘導的巨噬細胞中HMGB1分泌至細胞培養上清中;然而在無ATP刺激時,單獨LPS或LPS+燈盞花乙素并不會導致HMGB1的釋放(Fig 2C、2D)。上述結果表明,在J774A.1細胞中,燈盞花乙素能夠抑制ATP誘導的細胞焦亡的發生。
3.3燈盞花乙素抑制巨噬細胞發生細胞焦亡依賴于cAMP-PKA信號研究表明,燈盞花乙素可以上調細胞內cAMP的水平[13],而cAMP能夠激活下游的PKA,活化的PKA可以負調控NLRP3炎癥小體的活化[14]。因此,我們首先選用了腺苷酸環化酶特異性抑制劑MDL12330A抑制胞內cAMP水平,以探討燈盞花乙素對細胞焦亡的影響。經MDL12330A處理后,不僅能夠促進ATP誘導的細胞焦亡,還能逆轉燈盞花乙素對巨噬細胞中ATP誘導的細胞焦亡的抑制作用(Fig 3A、3B)。以上結果提示,燈盞花乙素抑制巨噬細胞發生細胞焦亡與cAMP水平密切相關。
進一步利用PKA的選擇性抑制劑H89抑制PKA活性,通過PI染色法對細胞焦亡進行檢測分析。結果顯示,燈盞花乙素預處理劑量依賴性地抑制J774A.1巨噬細胞中ATP誘導的細胞焦亡,而H89處理不僅能夠促進ATP誘導的細胞焦亡,還能逆轉燈盞花乙素對ATP誘導的細胞焦亡抑制作用(Fig 4A、4B)。上述結果提示,燈盞花乙素對NLRP3炎癥小體活化與細胞焦亡的抑制作用依賴于PKA信號。

Fig 1 Scutellarin inhibited ATP-induced activation of NLRP3 inflammasomes (±s, n=3)
A: Western blot analyses of the levels of indicated proteins in cell lysates and culture supernatants, respectively. β-tubulin was used as a loading control for cell lysates; B, C: Histograms showing the relative intensity of caspase-1p10 or IL-1β of Western blot. The intensity of caspase-1p10 or IL-1β bands in ATP group was set to 1.0; D: Cytometric bead array (CBA) assay for IL-1β levels in the culture supernatants.**P<0.01vsATP group. Scu: Scutellarin.

Fig2ScutellarininhibitedATP-inducedpyroptosisinJ774A.1macrophages
4 討論
NLRP3是固有免疫系統識別病原體的一類重要感受器,識別進入機體內的PAMP或DAMP,從而引起強烈的炎癥反應。細菌感染時,不僅細菌本身能釋放ATP,也能誘導單核/巨噬細胞釋放ATP,從而激活NLRP3炎癥小體,而引起急性期強烈炎癥反應,并誘發固有免疫細胞(包括巨噬細胞)的焦亡;同樣,器官受到損傷時也能導致細胞內DAMP(如ATP等)釋放到胞外,進而激活NLRP3炎癥小體和細胞焦亡,進一步加強損傷相關的炎癥反應。神經性疾病的發生和發展也與NLRP3有關,如多發性硬化癥、阿爾茨海默病、帕金森病等[6]。此外,NLRP3的功能失調與多種代謝疾病和免疫紊亂(如2型糖尿病、動脈粥樣硬化、痛風等)密切相關[6]。因此,抑制NLRP3炎癥小體的活化與細胞焦亡的發生,將有利于緩解上述NLRP3異常活化相關炎癥性疾病的癥狀[1]。本研究發現,在小鼠巨噬細胞J774A.1中,燈盞花乙素處理可以抑制ATP誘導的NLRP3炎癥小體的活化與細胞焦亡,從而抑制caspase-1的活化以及成熟IL-1β和HMGB1等炎癥因子的釋放,提示燈盞花乙素可作為一種治療NLRP3炎癥小體異常活化相關的炎癥性疾病的潛在藥物。
由于NLRP3的活化能夠誘發強烈的炎癥反應,因而受到多種調控分子或信號通路的調控。最近的研究發現,cAMP-PKA信號對NLRP3炎癥小體的活化也具有重要的調控作用。如膽汁酸通過其細胞膜受體TGR5提高胞內cAMP水平,從而激活PKA,活化的PKA誘導NLRP3的Ser 291(小鼠)發生磷酸化,進而抑制NLRP3活化[14];又如,前列腺素E2通過其受體EP4激活cAMP-PKA通路,促使NLRP3的Ser 295(人類)發生磷酸化,負調控NLRP3炎癥小體活化[15]。考慮到以往研究顯示燈盞花乙素可以上調cAMP的水平[13],在本研究中我們初步探討了cAMP-PKA信號在燈盞花乙素抑制NLRP3炎癥小體活化與細胞焦亡過程中的作用,發現腺苷酸環化酶抑制劑MDL12330A和PKA抑制劑H89都能完全逆轉燈盞花乙素對ATP誘導的細胞焦亡的抑制作用,提示燈盞花乙素可能通過調節PKA的活性而抑制ATP誘導的NLRP3炎癥小體活化與細胞焦亡。


A: Cell death in J774A.1 cells was measured by staining with PI (red, staining dead cells) and Hoechst 33342 (blue, staining all cells). Scale bar=50 μm; B: PI-positive cells in five randomly chosen fields containing ~100 cells in each were quantified.**P<0.01vsvehicle; ns: not significant.

Fig4ScutellarininhibitedATP-inducedpyroptosisviaregulatingPKAsignaling
A: Cell death in J774A.1 cells was assayed by PI (red) and Hoechst 33342 (blue). Scale bars=50 μm; B: PI-positive cells in five randomly chosen fields containing ~100 cells in each were quantified.**P<0.01vsvehicle; ns: not significant.
總之,本研究發現,燈盞花乙素可以明顯抑制小鼠巨噬細胞J774A.1中ATP誘導的NLRP3炎癥小體的活化與細胞焦亡,這種抑制作用可能由cAMP/PKA信號通路所介導。雖然燈盞花乙素抑制NLRP3活化的具體機制還需要進一步研究闡明,本研究結果顯示燈盞花乙素可能用于預防和治療NLRP3異常活化或功能失調所引起的炎癥性疾病;同時,這也為燈盞花乙素作為抗炎藥物發揮作用提供了新的實驗證據。
[1] Lamkanfi M, Dixit V M. Mechanisms and functions of inflammasomes[J].Cell, 2014,157(5): 1013-22.
[2] Wegiel B, Larsen R, Gallo D, et al. Macrophages sense and kill bacteria through carbon monoxide-dependent inflammasome activation[J].JClinInvest, 2014,124(11): 4926-40.
[3] 王 偉, 戴 敏, 徐忠東. 丹皮酚對脂多糖/三磷酸腺苷誘導的小膠質細胞NLRP3炎癥小體激活的影響[J]. 中國藥理學通報, 2014,30(5): 652-6.
[3] Wang W, Dai M, Xu Z D. Effects of paeonal on lipopolysaccharide/ adenosine 5′-triphosphate induced NLRP3 inflammasome activation in primary rat microglia[J].ChinPharmacolBull, 2014,30(5): 652-6.
[4] Labbe K, Saleh M. Cell death in the host response to infection[J].CellDeathDiffer, 2008,15(9): 1339-49.
[5] Strowig T, Henao-Mejia J, Elinav E, et al. Inflammasomes in health and disease[J].Nature, 2012,481(7381): 278-86.
[6] Guo H, Callaway J B, Ting J P. Inflammasomes: mechanism of action, role in disease, and therapeutics[J].NatMed, 2015,21(7): 677-87.
[7] Li H, Huang D, Gao Z, et al. Scutellarin inhibits cell migration by regulating production of αvβ6 integrin and E-cadherin in human tongue cancer cells[J].OncolRep, 2010,24(5): 1153-60.
[8] Xing J F, You H S, Dong Y L, et al. Metabolic and pharmacokinetic studies of scutellarin in rat plasma, urine, and feces[J].ActaPharmacolSin, 2011,32(5): 655-63.
[9] 石美娜, 楊為民, 劉 璇. 燈盞花乙素藥理作用研究進展[J]. 昆明醫科大學學報, 2013,34(9): 151-4.
[9] Shi M N, Yang W M, Liu X. Research progress of scutellarin pharmacological study[J].JKunmingMedUniv, 2013,34(9): 151-4.
[10] Niu C, Sheng Y, Yang R, et al. Scutellarin protects against the liver injury induced by diosbulbin B in mice and its mechanism[J].JEthnopharmacol, 2015,164(3): 301-8.
[11] Ito M, Shichita T, Okada M, et al. Bruton’s tyrosine kinase is essential for NLRP3 inflammasome activation and contributes to ischaemic brain injury[J].NatCommun, 2015,6(6):7360.
[12] 魏紅霞, 李陳廣, 梁譯丹, 等. 二甲雙胍增強LPS激活的腹腔巨噬細胞在ATP刺激下的炎癥小體活化[J]. 中國藥理學通報, 2017,33(4):474-9.
[12] Wei H X, Li C G, Liang Y D, et al. Metformin enhances ATP-stimulated inflammasome activation in LPS-primed peritoneal macrophages[J].ChinPharmacolBull, 2017,33(4): 474-9.
[13] Tian X, Chang L, Ma G, et al. Delineation of platelet activation pathway of scutellarein revealed its intracellular target as protein kinase C[J].BiolPharmBull, 2016,39(2): 181-91.
[14] Guo C, Xie S, Chi Z, et al. Bile acids control inflammation and metabolic disorder through inhibition of NLRP3 inflammasome[J].Immunity, 2016,45(4): 802-16.
[15] Mortimer L, Moreau F, MacDonald J A, et al. NLRP3 inflammasome inhibition is disrupted in a group of auto-inflammatory disease CAPS mutations[J].NatImmunol, 2016,17(10): 1176-86.
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
東方少年·布老虎畫刊(2023年8期)2023-08-01 15:45:12
科學大眾(2021年6期)2021-07-20 07:42:44
科學(2020年3期)2020-11-26 08:18:30
學苑創造·A版(2020年9期)2020-10-13 09:41:02
娃娃樂園·綜合智能(2019年3期)2019-04-03 09:17:36
中成藥(2018年2期)2018-05-09 07:19:34
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
湖北師范大學學報(自然科學版)(2015年2期)2016-01-10 08:41:55
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
