CMAH基因在小鼠不同組織中的轉錄水平分析
2018-01-31 06:30:00李一帆尹德琦桑曉宇漢麗梅
中國比較醫學雜志 2018年1期
李一帆,楊 娜,王 瑤,尹德琦,趙 旭,桑曉宇*,漢麗梅*
(1.沈陽農業大學畜牧獸醫學院,沈陽 110866; 2.遼寧省人獸共患病研究重點實驗室,沈陽 110866)
唾液酸(sialic acid)是通過N末端或O末端連接在糖蛋白或糖脂末端單糖家族的總稱。唾液酸的核心結構包含9個碳原子,并且通常在第4、5、7、8和9號位的碳原子發生修飾可形成不同的唾液酸衍生物[1]。根據5號位的碳原子連接的基團不同,可以將唾液酸初步分為N-乙酰神經氨酸(N-acetylneuraminic acid,Neu5Ac)、N-羥乙酰神經氨酸(N-glycolylneuraminic acid,Neu5Gc)和脫氨神經氨酸(deaminoneuraminic acid,KDN)[2]。在哺乳動物細胞中唾液酸的主要存在形式包括Neu5Ac和Neu5Gc,并且在胞苷單磷酸-N-乙酰神經氨酸羥化酶(cytidine monophospho-N-acetylneuraminic acid hydroxylase,CMAH)的作用下CMP-Neu5Ac可生成Neu5Gc[3]。研究者已經在豬氣管上皮細胞[4]、鴨結腸的隱窩上皮細胞、馬氣管上皮細胞和鴿子的結腸與氣管中檢測到Neu5Gc。不僅如此,研究者應用高效液相色譜(HPLC)在雞的氣管和腸道表面也檢測到Neu5Gc[5]。由于人體內的CMAH基因丟失了92 bp,導致人失去了合成Neu5Gc的能力[4]。最近研究表明,與人類的情況類似,雪貂也不能合成Neu5Gc[6]。
研究表明多種病原微生物都能利用在哺乳動物細胞表面廣泛存在的唾液酸作為受體,包括細菌、寄生蟲和病毒等[7],與之結合并入侵宿主細胞,但不同的病原選擇不同的唾液酸衍生物作為受體[8]。A型和B型流感病毒主要識別宿主細胞表面以2, 3糖苷鍵或2, 6糖苷鍵連接的Neu5Ac,而馬流感病毒主要結合受體是Neu5Gc[9]。研究表明在A549細胞表面外源表達的Neu5Gc受體能夠抑制A型流感病毒的感染[10]。不僅如此,有研究表明人體紅細胞表面缺少Neu5Gc受體的表達,能夠限制諾氏瘧原蟲的跨種傳播[11]。相反,多瘤病毒、輪狀病毒等同時能夠識別Neu5Ac和Neu5Gc并作為入侵宿主的受體[12]。由此可見,宿主細胞表面唾液酸受體的類型與病原的入侵密切相關。
不同物種或同一物種的不同組織中細胞表面多聚糖末端結合的Neu5Gc和Neu5Ac所占比例都不同。在小鼠和大鼠組織中Neu5Gc和CMP-Neu5Ac的表達比例主要依靠CMAH的活性水平[13]。盡管腦組織中的唾液酸含量非常高[14],但在大多數哺乳動物腦組織內很少能夠檢測到Neu5Gc。在神經組織中起到非常重要作用的神經節苷脂和神經細胞粘附分子(neural cell adhesion molecule,NCAM)中的多聚唾液酸僅包含Neu5Ac。本研究通過應用相對熒光定量PCR的方法檢測Neu5Cc合成途徑中關鍵酶(CMAH)的基因在不同小鼠組織中轉錄水平,為進一步分析不同組織中所含Neu5Gc含量差異提供理論參考。
1 材料和方法
1.1 實驗動物
采用遼寧省實驗動物資源中心培育的5只SPF級雌性BALB/c小鼠,體重18~20 g [SCXK (遼) 2015-0001],[SYXK (遼) 2015-0001],質量檢測單位:中國食品藥品檢定研究院。
1.2 主要試劑與儀器
BIOZOL Total RNA Extraction Reagent(TRIzol)購于Bio Flux公司;PrimeScriotTMRT Reagent Kit with gDNA Eraser(反轉錄試劑盒)與SYBR? Premix Ex TaqTMII(熒光定量PCR試劑盒)均購于Takara公司;2× RNA Loading Buffer購于Solarbio公司;DEPC購于Amresco公司。
實時熒光定量PCR儀器:Thermo Fisher公司QuantStudio? 6 Flex;電泳儀:北京六一生物技術有限公司DYY-7C型;凝膠成像儀:UVP公司GelDoc-It2 Imager型;低溫離心機:德國Eppendorf公司Centrifuge 5427R型。
1.3 實驗方法
1.3.1 實驗樣本的采集
組織樣品采集于5只健康的SPF級BALB/c鼠。實驗過程中按實驗動物使用的3R原則給予人道主義關懷。各樣品均采集于不同臟器的相同部位。為保證提取RNA的質量,用乙醚致暈小鼠后對其斷頸處死,立即采集其肝臟、心臟、脾臟、肺臟、腎臟、小腸、骨骼肌、氣管及腦組織,并用預冷的生理鹽水漂洗血液,后迅速剪成小塊(約0.1 g),浸入液氮速凍后,立刻投入裝有TRIzol的2 mL去酶離心管,于-80℃凍存備用。

表1 CMAH與β-actin基因熒光定量PCR引物
1.3.2 RNA的提取、檢驗與反轉錄
采用TRIzol法提取各組織中的RNA;采用Nano Drop 2000c檢測其濃度,通過OD260/OD280與OD260/OD230的比值初步判斷RNA是否有降解和污染;并運用RNA甲醛變性瓊脂糖凝膠(1%)電泳對其完整性進行檢驗,電泳條件為:預電泳80 V(15 min),正式電泳50 V(50 min)。采用PrimeScriotTMRT Reagent Kit with gDNA Eraser反轉錄試劑盒進行反轉錄:首先42℃,2 min去除基因組DNA;再選擇隨機引物經37℃,15 min進行反轉錄;最后經85℃,5 s終止反應。
1.3.3 熒光定量PCR方法
(1)引物設計:根據GenBank中登錄的小鼠CMAH基因序列(NM_001111110)和β-actin基因序列(NM_007393),應用ABI Primer Express 3.0實時熒光定量PCR引物設計軟件設計熒光定量PCR引物,其中以β-actin作為內參基因。對所設計引物的特異性和擴增效率等因素進行分析,選擇引物序列如表1所示。引物由上海捷瑞生物工程有限公司合成。
(2)制作相對熒光定量PCR標準曲線:根據實驗需求與條件選用SYBR Green,2-ΔCt相對定量法進行熒光定量PCR檢測。將原始cDNA模板作3倍梯度稀釋后進行熒光定量PCR擴增,分別制作2對引物標準曲線,探索退火溫度使各對引物的擴增效率在其它各項指標的允許范圍內接近100%,最終確定反應體系為SYBR Premix Ex Taq II(2×)10 μL,ROX Reference Dye II(50×)0.4 μL,PCR forward primer(10 μmol/L)0.8 μL,PCR reverse primer(10 μmol/L)0.8 μL,template 2 μL,ddH2O 6 μL;最佳反應條件為:預變性95℃,30 s;PCR反應95℃,5 s,60℃,30 s,72℃,30 s,共40個循環,在每個循環結束時采集熒光信號;之后進行溶解曲線分析,95℃,15 s,60℃,60 s,95℃,15 s。
(3)qPCR相對定量檢測:陰性對照以水為模板,20 μL反應體系,分別做5個生物學重復及3個組內重復。
1.4 統計學方法
所得qPCR相對定量數據應用SPSS軟件進行One-way ANOVA分析,以P< 0.05為差異有顯著性,應用GraphPad Prism 5軟件制圖。
2 結果
2.1 RNA的質量與完整性檢測結果
檢測所提取各組織總RNA的OD260/OD280比值均介于1.90~2.10之間,OD260/OD230的比值均介于1.90~2.20之間,且稍大于OD260/OD280的比值,其濃度也滿足反轉錄需要。將BALB/c小鼠各組織總RNA進行甲醛變性瓊脂糖凝膠,完整性檢測如圖1所示,各樣品中的28s、18s、5s三條特異性條帶均清晰可見,且28s亮度約為18s的2倍,說明所提取的總RNA完整性較好,無明顯降解,滿足后續實驗需求。
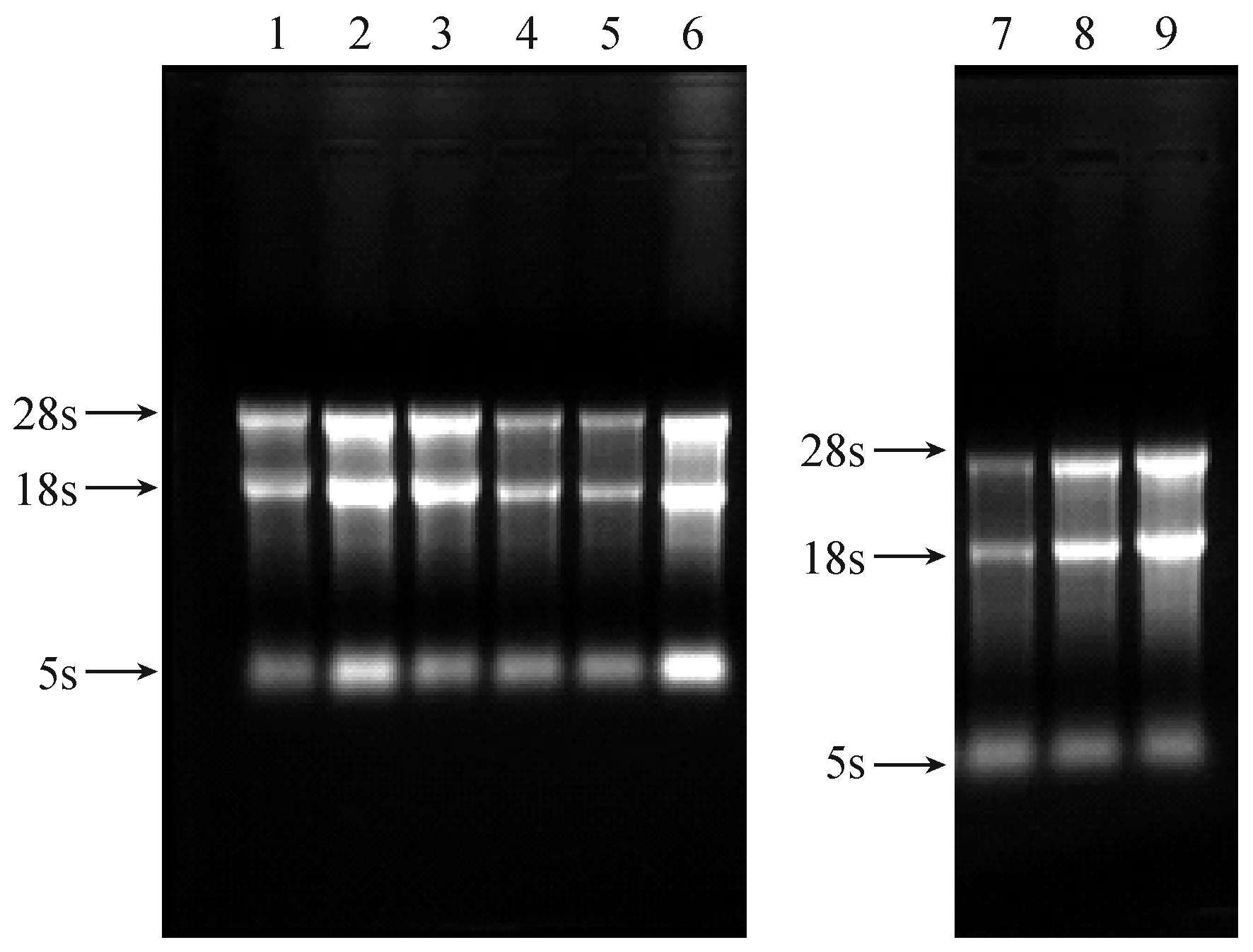
注:1:心肌;2:肝臟;3:氣管;4:小腸;5:腎臟;6:腦組織;7:骨骼肌;8:肺臟;9:脾臟。圖1 總RNA完整性檢驗Note. 1: Myocardium; 2: Liver; 3: Trachea; 4: Small intestine; 5: Kidney; 6: Brain; 7: Skeletal muscle;8: Lung; 9: Spleen.Fig.1 Integrity of the total RNA
2.2 相對標準曲線的建立
本實驗選擇SYBR Green染料法,以3倍梯度稀釋肝臟組織的原始cDNA,不少于5個稀釋點,以水為模板作陰性對照,均重復三個加樣孔,分別制作CMAH與β-actin兩對引物的相對標準曲線。擴增曲線如圖2A、2B所示,模板3倍梯度稀釋擴增曲線重復性好,間距一直。溶解曲線如圖2C、2D所示,各引物均為特異性的單峰,無非特異性擴增,說明各對引物特異性良好,并且無引物二聚體等結構。兩基因相對標準曲線如圖2E、2F所示,β-actin與CMAH引物的擴增效率分別為101.728%、104.210%,接近100%,組內差距小,重復性好。
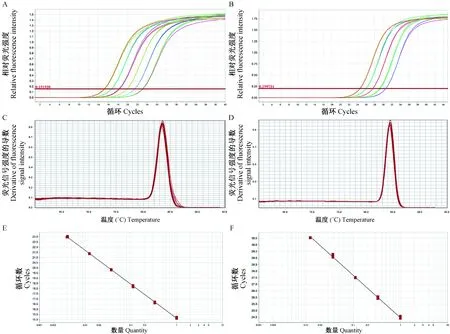
注:A:β-actin基因的擴增曲線;B:CMAH基因的擴增曲線;C:β-actin基因的溶解曲線;D:CMAH基因的溶解曲線;E:β-actin基因的相對標準曲線;F:CMAH基因的相對標準曲線。圖2 相對標準曲線的建立Note. A: Amplification curve of β-actin. B: Amplification curve of CMAH. C: Dissolution curve of β-actin. D: Dissolution curve of CMAH. E: Standard curve of β-actin. F: Standard curve of CMAH.Fig.2 Establishment of the relative standard curves
以上數據證明本試驗實驗所選2對熒光定量PCR引物及其相應反應條件可滿足SYBR Green、2-ΔCt相對定量法進行熒光定量PCR檢測的條件,實驗結果真實可信。
2.3 qPCR相對定量檢測結果
根據上文中所建立的相對熒光定量PCR的實驗條件,分別對小鼠的肝臟、脾臟、腎臟、氣管、肺臟、心肌、骨骼肌、小腸和腦組織的CMAH與β-actin基因進行熒光定量PCR檢測,檢測結果如表2所示,并選擇2-ΔCt進行差異表達分析。實驗結果如圖3所示:CMAH基因在BALB/c小鼠肝臟內表達最為豐富,后依此為脾臟、腎臟、氣管、肺臟、心肌、骨骼肌和小腸,而腦組織中表達量極低。其中肝臟組織CMAH基因表達量分別達到脾臟的2.46倍;腎臟的3.17倍;氣管的5.14倍;肺臟的11.70倍;心肌的21.12倍;骨骼肌的31.37倍;小腸的66.90倍;腦組織的1055.99倍。
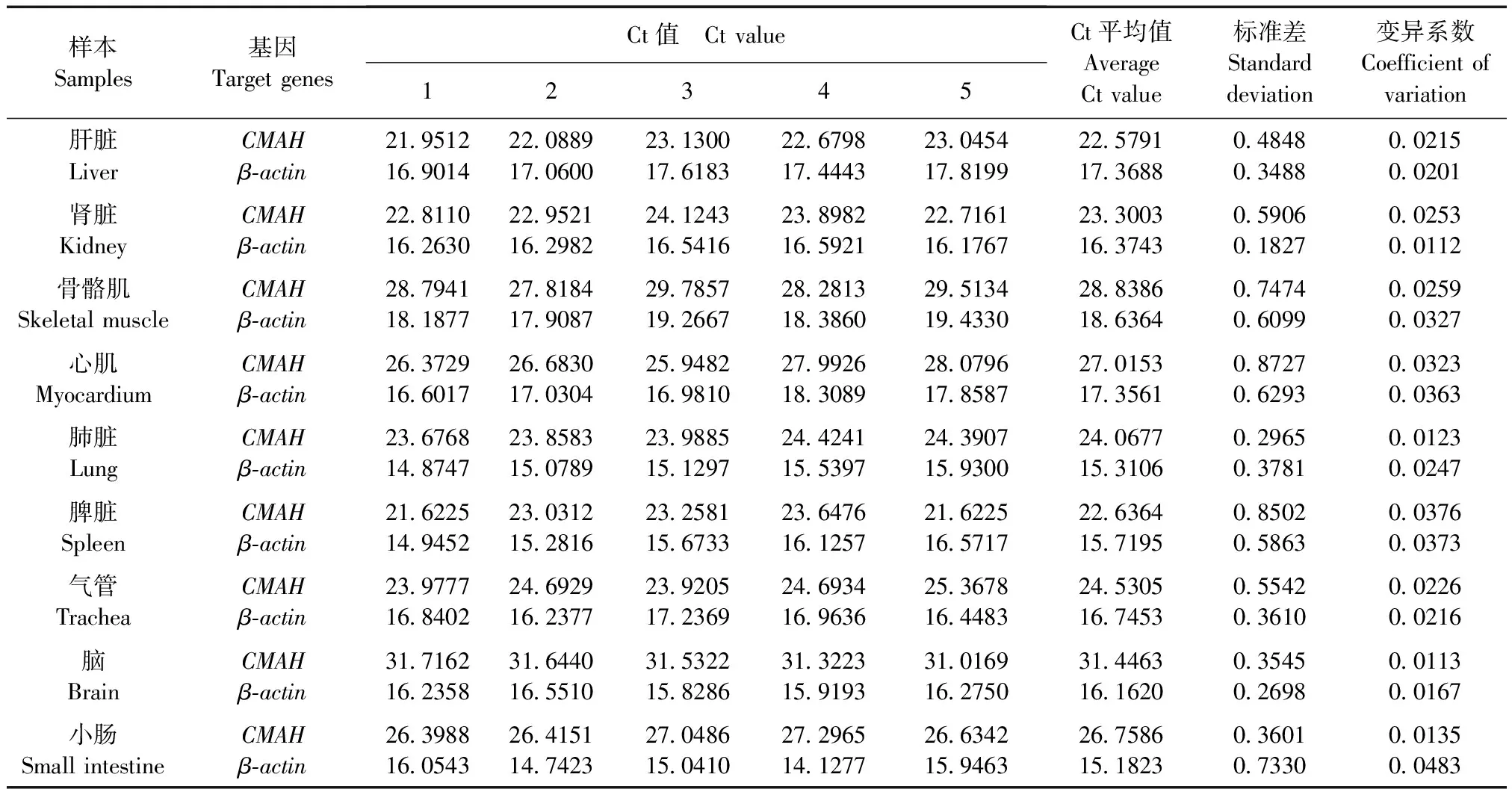
表2 BALB/c 小鼠各組織臟器CMAH/β-actin基因熒光定量PCR檢測結果
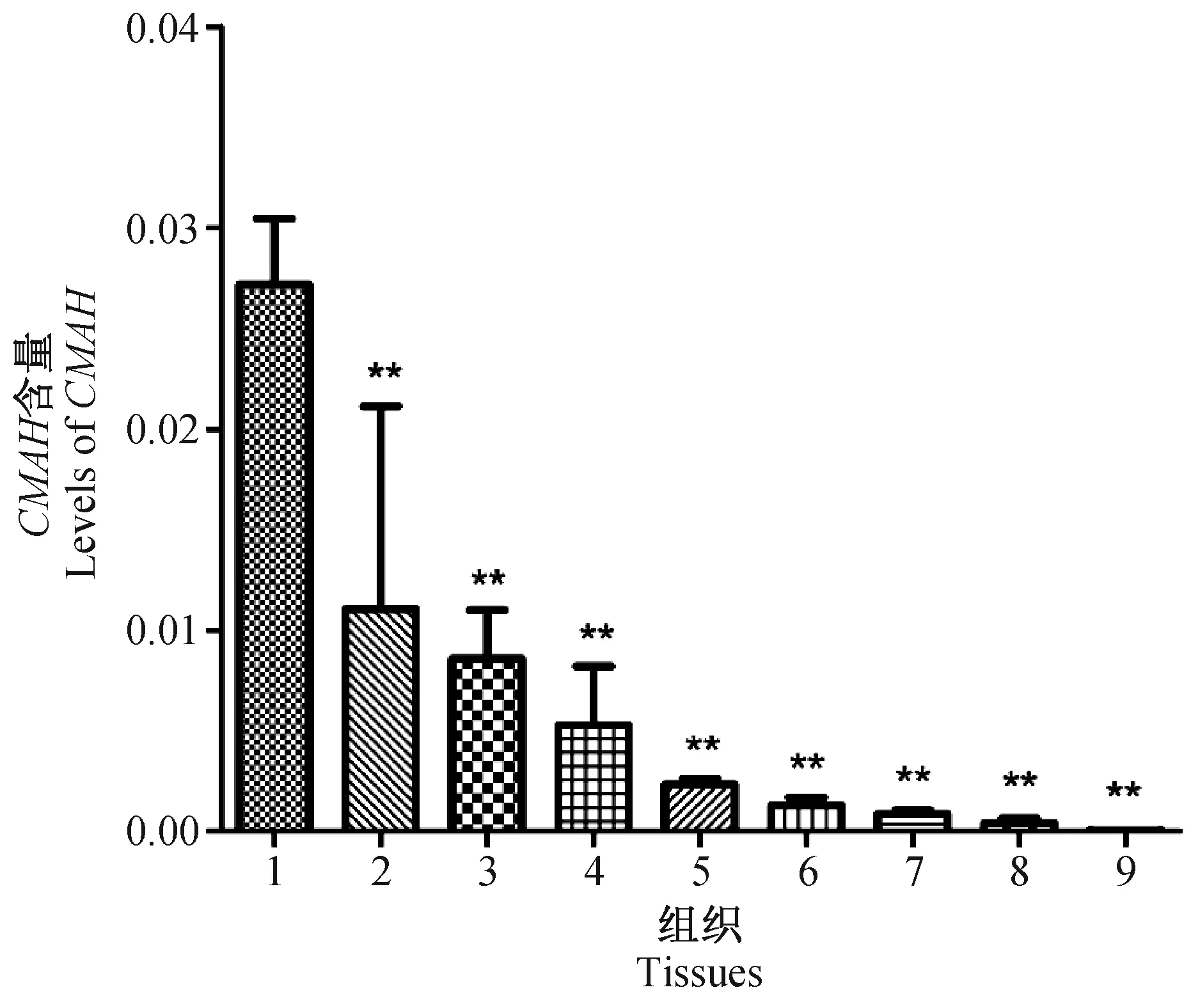
注:1:肝臟;2:脾臟;3:腎臟;4:氣管;5:肺臟;6:心肌;7:骨骼肌;8:小腸;9:腦。以內參(β-actin)的轉錄水平為1并對其他轉錄水平進行校正。與肝臟比較,** P< 0.01。圖3 CMAH基因在不同組織中的轉錄水平Note. 1: Liver; 2: Spleen; 3: Kidney; 4: Trachea; 5: Lung; 6: Myocardium; 7: Skeletal muscle; 8. Small intestine; 9: Brain. Transcriptional levels of CMAH were normalized to those of the internal control (β-actin). Compared with the liver,** P < 0.01.Fig.3 Transcriptional levels of CMAH in different tissues
3 討論
細胞表面的唾液酸可參與細胞-細胞、細胞-病原體之間的信息交換[15]、免疫反應、炎癥反應及大量粘附過程。當前研究表明,細胞表面兩種重要唾液酸衍生物就是Neu5Ac和Neu5Gc。CMAH可催化Neu5Ac發生羥基化合成Neu5Gc,此過程具有高度特異性。在不同種動物相同組織內及在同種動物不同組織內,Neu5Ac與Neu5Gc所占比例均有差異[16]。由于CMAH基因堿基缺失、CMAH蛋白表達障礙或失活等原因,包括人在內的一些動物(雪貂、海豹、部分爬行動物和鳥類等)都不能合成Neu5Gc[17],但在其他哺乳動物中CMAH基因卻可以高效表達并且Neu5Gc含量豐富[18],但是又有明顯的組織差異性。已有研究表明,小鼠血清中含有Neu5Gc,說明小鼠CMAH基因能夠表達具有生物學功能的羥化酶,但對不同組織中CMAH基因轉錄水平的研究較少[7]。本研究中,通過相對定量PCR方法能夠在正常BLAB/c小鼠的9種組織中檢測到CMAH基因的轉錄,但其轉錄情況差異較大,肝臟中的轉錄水平最高,其次依次為脾臟、腎臟、氣管,而肺臟、心肌、骨骼肌、小腸和腦組織中的轉錄水平都比較低,此結果與先前發表文章的實驗結果相一致[18]。當前,我們尚未完全闡明不同物種或者同一個體間不同組織的Neu5Gc表達差異的調控機制,但宿主細胞表面的唾液酸組成類型確實能夠影響不同病原與宿主之間的相互作用,值得我們進一步探索。
熒光定量PCR方法自建立以來已經廣泛應用檢測基因表達調控的初步探索,根據中心法則,我們知道基因轉錄情況的分析不能完全反映出蛋白表達情況,但是與Western blot等免疫學方法相比較,熒光定量PCR方法可以快速檢測出目的基因轉錄情況,以此來初步評估蛋白的表達水平。因此,本研究成功建立BALB/c小鼠CMAH基因相對熒光定量PCR檢測方法,并確定其在多種組織中均有轉錄但差異較大,此結果為進一步分析BALB/c小鼠不同組織中所含Neu5Gc差異提供參考依據。
[1] Takahashi T, Takano M, Kurebayashi Y, et al. N-glycolylneuraminic acid on human epithelial cells prevents entry of influenza A viruses that possess N-glycolylneuraminic acid binding ability [J]. J Virol, 2014, 88(15): 8445-8456.
[2] Wasylnka JA, Simmer MI, Moore MM. Differences in sialic acid density in pathogenic and non-pathogenic Aspergillus species [J]. Microbiology, 2001, 147(Pt 4): 869-877.
[3] 程鋮, 高春芳. 唾液酸的生物學意義及其在肝病中的研究進展 [J]. 檢驗醫學, 2013, 28(4): 333-336.
[4] Wasik BR, Barnard KN, Parrish CR. Effects of sialic acid modifications on virus binding and infection [J]. Trends Microbiol, 2016, 24(12): 991-1001.
[5] Malykh YN, Shaw L, Schauer R. The role of CMP-N-acetylneuraminic acid hydroxylase in determining the level of N-glycolylneuraminic acid in porcine tissues [J]. Glycoconj J, 1998, 15(9): 885-893.
[6] 范曉琳, 蘇如婷, 韓曉駿. 糖尿病慢性腎臟疾病患者血清唾液酸的變化及其與血管內皮功能關系的研究 [J]. 中國糖尿病雜志, 2015, 23(8): 717-719.
[7] Ng PS, B?hm R, Hartley-Tassell LE, et al. Ferrets exclusively synthesize Neu5Ac and express naturally humanized influenza A virus receptors [J]. Nat Commun, 2014, 5: 5750.
[8] 宋宇, 周潔, 高誠, 等. 哺乳動物呼腸孤病毒外殼蛋白和受體研究進展 [J]. 實驗動物與比較醫學, 2010, 30(1): 68-72.
[9] Diaz SL, Padler-Karavani V, Ghaderi D, et al. Sensitive and specific detection of the non-human sialic acid N-glycolylneuraminic acid in human tissues and biotherapeutic products [J]. PLoS One, 2009, 4(1): e4241.
[10] Varki A. Sialic acids in human health and disease [J]. Trends Mol Med, 2008, 14(8): 351-360.
[11] Dankwa S, Lim C, Bei AK, et al. Ancient human sialic acid variant restricts an emerging zoonotic malaria parasite [J]. Nat Commun, 2016, 7: 11187.
[12] 楊威. N-乙酰神經氨酸衍生物的合成及其在快速檢測流感病毒中的應用 [D]. 東北師范大學, 2009.
[13] Kavaler S, Morinaga H, Jih A, et al. Pancreatic β-cell failure in obese mice with human-like CMP-Neu5Ac hydroxylase deficiency [J]. FASEB J, 2011, 25(6): 1887-1893.
[14] 科技日報. 斑馬魚研究首次識別出新遺傳病 唾液酸確認有助大腦與骨骼發育 [J]. 江西飼料, 2016, 23(4): 43.
[15] 宋戰昀, 丁壯. 病毒細胞膜唾液酸受體研究進展 [C]. 中國畜牧獸醫學會家畜傳染病學分會第六屆理事會第二次會議暨教學專業委員會第六屆代表大會論文集. 2006.
[16] 許永華, 張東輝, 是文輝, 等. Wistar大鼠和昆明種小鼠血清唾液酸頻率分布與參考值 [J]. 中國比較醫學雜志, 2006, 16(9): 541-543.
[17] 占玲俊, 鄧巍, 鮑琳琳, 等. 唾液酸受體并非流感病毒各亞型在雪貂組織中播散分布的決定因子 [J]. 中國比較醫學雜志, 2012, 22(4): 23-26.
[18] Kwon DN, Chang BS, Kim JH. Gene expression and pathway analysis of effects of the CMAH deactivation on mouse lung, kidney and heart [J]. PLoS One, 2014, 9(9): e107559.
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
學苑創造·A版(2020年9期)2020-10-13 09:41:02
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
海峽科技與產業(2016年3期)2016-05-17 04:32:12
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
