綿羊iPS細胞誘導及轉錄組學分析
2018-02-25 10:56:30張譯元郭延華王聰慧南海艷王立民
新疆農業科學 2018年11期
張譯元,郭延華,王聰慧,唐 紅,南海艷,王立民,周 平
(1.新疆農墾科學院畜牧獸醫研究所/省部共建綿羊遺傳改良與健康養殖國家重點實驗室,新疆石河子 832000;2.陜西西安市閻良區動物疾病預防控制中心,西安 710000)
0 引 言
【研究意義】在綿羊的遺傳育種與品種改良進程中,快速、定向選育具有某些優質性狀的新品種或品系,是綿羊種質資源保護和綿羊生產的關鍵環節。以往轉基因技術克隆成功率低,且獲得的轉基因后代個體往往存在繁殖障礙。誘導性多能干細胞(induced pluripotent stem cells, iPS)。iPS細胞是一類具有無限增殖能力[1]和多向分化潛能[2]的細胞類群,可塑性強,能夠高效實現外源基因的導入、敲除和改造。iPS最初是由日本科學家山中伸彌(Takahashi, Yamanaka, 2006)通過病毒載體將OCT4(Octamer binding transcription factor 4)、SOX2(SRY(sex determining region Y)-box2)、KLF4(Kruppel-like factor 4)、c-Myc(Myelocytomatosis oncogene)這四個轉錄因子轉入小鼠成纖維細胞中,使之重編程構建出來的。如果把iPS細胞誘導技術和基因打靶技術、細胞核移植技術、嵌合體技術相結合,對綿羊的轉基因育種和生產意義重大。【前人研究進展】迄今為止,有關人(Homosapiens)[3,4]、小鼠(Musmusculus)[5]、大鼠(Rattusnorvegicus)[6]、猴子(Macacamulatta)[7]、豬(Susscrofa)[8]、綿羊(Ovisaries)[9]的iPS細胞系構建均已有研究報道。2006年Takahashi和Yamanaka將小鼠成纖維細胞誘導為多能干細胞,并通過四倍體囊胚嵌合技術,進一步證實小鼠iPS細胞能夠發育成完整的個體[10]。2007年Thomson和Yamanaka兩實驗室又分別將人體皮膚成纖維細胞誘導成iPS細胞[3,4]。在綿羊iPS細胞系研究方面,Bao等[5]和Liu等[11]先后獲得了能形成畸胎瘤和擬胚體并具有正常核型的綿羊iPS細胞系。Liu等[11]將綿羊iPS細胞注射到二倍體或四倍體胚胎中,形成了內細胞團的部分結構,但未形成嵌合體。Sartori等[12]將Oct4、Sox2、 Klf4、c-Myc四個轉錄因子(OSKM)轉入綿羊成纖維細胞,形成了具有正常核型的iPS細胞,并形成畸胎瘤和擬胚體;將獲得的iPS細胞植入到胚胎后,生出了嵌合體小羊。【本研究切入點】綿羊iPSCs誘導技術取得了階段性進展,但綿羊iPSCs形態特征及鑒定方法并無統一的標準,誘導方法及培養條件也形色各異。目前,綿羊iPS細胞的誘導基本采用病毒載體轉染的方式,存在一定安全風險;其次,由于物種不同,多能性基因、信號通路也會不同[13],重編程因子的選擇對iPS細胞的誘導效率會造成一定影響。因此,如何提高誘導效率、降低致癌率和抑制iPSCs的分化是目前綿羊iPSCs急需解決的問題。試驗以綿羊成纖維細胞為基礎材料,通過電轉,導入五種轉錄因子OCT4、SOX2、KLF4、l-Myc、Lin28,構建綿羊iPS細胞系;在此基礎上結合轉錄組測序技術重點對不同誘導時期的iPS細胞進行高通量測序。【擬解決的關鍵問題】分析相關因子在誘導不同階段的表達變化,研究差異基因所涉及的信號通路,選擇更為適合綿羊iPS細胞誘導的重編程因子,為豐富綿羊iPS建系機制提供一定的基礎資料,為綿羊轉基因育種及動物種質資源保護提供理論依據。
1 材料與方法
1.1 材 料
試驗動物來自新疆農墾科學院轉基因種羊場。
重組質粒:PCXLE-Hoct3/4-shp53(Oct3/4)、PCXLE-hSK(Sox2, Klf4)、PCXLE-hul(l-Myc, lin28)等,均購自美國Addgene公司,Lonza 4D-Nucleofector電轉儀與P2電轉染試劑盒購自德國LONZA公司;反轉錄試劑盒PrimeScriptTMRT reagent Kit with gDNA Eraser購自TAKARA公司;堿性磷酸酶試劑盒、免疫熒光染色所用抗體購自上海斯丹賽生物技術有限公司,DAPI 染料購自ROCHE公司。
1.2 方 法
1.2.1 綿羊iPS細胞的誘導及培養
離心收集對數期的綿羊成纖維細胞,細胞密度至5×106mL-1;添加20 μL電轉液和重組質粒,重懸細胞,選擇EH-100程序進行電轉;靜置10 min,加入80 μL細胞培養液(10%胎牛血清的DMEM培養基),混勻后全部移至培養皿,37℃、5%CO2條件下培養9~12 h后更換培養液,36 h時觀察細胞的貼壁情況。3 d后換成iPS細胞培養液(20% KSR、0.1 mol/L β-巰基乙醇、0.02 mol/L谷氨酰胺、0.01 mol/L非必需氨基酸(NEAA)、0.005 g/L b FGF的Knockout DMEM培養基),培養液中添加終濃度為0.05 mg/L VC,隔天換液。
1.2.2 堿性磷酸酶(AP)染色
用4%(M/V)多聚甲醛固定iPS克隆樣細胞,15 min后棄固定液,PBS洗滌2次,滴加染色液(現配現用),待染色液掩蓋孔底,室溫避光15~20 min,棄染色液,PBS洗滌1次,顯微鏡下觀測染色結果。
1.2.3 免疫熒光染色
4%(W/V)多聚甲醛固定細胞20 min,1×PBS浸洗2次,無水乙醇浸泡2次,每次20 min;1×PBS浸洗2次,吸水紙吸干PBS,在細胞上滴加5%(W/V)牛血清白蛋白(BSA),37℃封閉30 min;吸水紙吸掉封閉液,不洗,滴加足夠量稀釋好的一抗,4℃過夜孵育,PBST浸洗細胞3次,每次5 min;吸水紙吸干細胞上多余液體后滴加稀釋好的熒光二抗,37℃孵育1 h,PBST浸洗細胞3次,每次5 min。復染核:滴加1 μg/mL的DAPI避光孵育5 min, PBST 5 min×3次洗去多余DAPI,吸水紙吸去細胞上殘余液體,然后在熒光顯微鏡下觀察采集圖像。
1.2.4 實時熒光定量PCR檢測
提取細胞的總RNA,反轉錄成cDNA,以綿羊GAPDH基因(F: ACGGGAAGCTCACTGGCATGG R: GCCAGCCCCAGCATCGAAG)為靶標,對Oct4(F: TACACTGTACTCTTCGGTCCCATT R: AGCATCATTGAACTTCACCTTCCC)、Sox2(F:TACGGTAGGAGCTTTGCAGAAAGT R: TGCACGTTTGCAACTGTCCTAAAT)、Nanog(F:CAAGTATTTCAGTTCCCAGCAGCA R: TGCACGTTTGCAACTGTCCTAAAT)基因使用LightCycler? 480實時熒光定量PCR儀(Roche公司)進行熒光定量PCR檢測,反應條件:95℃預變性5 min,95℃變性15 s,60℃退火30 s,72℃延伸1 min,反應進行45個循環,65℃再延伸1 min并在這一過程中進行熔解曲線分析。基因的相對表達量采用2-△△Ct法計算,使用GraphPad Prism 5進行數據統計處理分析。
1.2.5 轉錄組測序
分別收集誘導0、10、20和30 d的細胞,由北京華諾生物科技有限公司提供測序服務,使用Illumina HiseqTM2000平臺測序。
1.3 數據處理
測序所得數據稱之為raw reads,將其測序接頭序列末端質量低于20 bp的堿基、含N比率大于5%和質量較低的reads去除,去除后的數據稱為clean reads。對clean reads使用Trinity軟件進行De novo組裝得到Unigene,然后使用Tgicl軟件對這些序列進行同源轉錄本聚類,得到最終的Unigene。之后將Unigene序列與蛋白數據庫NR、Swiss-Prot、KEGG和COG作blastx比對(evalue<0.000 01),獲得相應的注釋信息。針對Unigene表達量的計算,使用FPKM法(Fragments Per kb per Million fragments)。以FDR≤ 0.001且倍數差異在2倍以上的基因定義為差異表達基因,將所得到的差異基因GO數據庫和KEGG數據庫富集進行注釋與統計分析。
2 結果與分析
2.1 形態學鑒定
該細胞排列緊密,呈圓形或不規則集落狀生長,形似鳥巢,在集落邊緣可見部分分化細胞。堿性磷酸酶染色(AP)呈強陽性。圖1
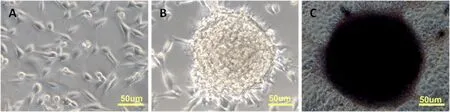
圖1 A:綿羊成纖維細胞;B:綿羊iPSC克隆;C:綿羊iPSC克隆堿性磷酸酶染色
Fig.1 A: sheep fibroblast cells; B: OiPSC colony ; C: AP staining
2.2 免疫熒光檢測
對誘導培養的綿羊iPS細胞進行細胞免疫熒光檢測,結果顯示克隆多能性蛋白SOX2、OCT4和表面標記SSEA-1呈陽性。圖2
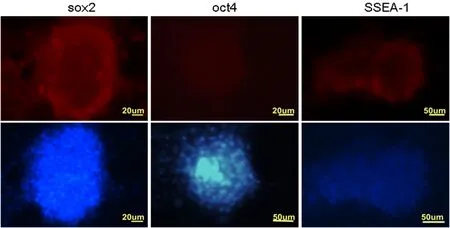
圖2 綿羊iPS細胞蛋白免疫熒光檢測
Fig.2 The immune fluorescence identification of OiPSC
2.3 實時熒光定量RT-PCR
隨機選取4個綿羊iPS細胞,以綿羊成纖維細胞為陰性對照,實時熒光定量PCR檢測OCT4、SOX2、Nanog的表達水平。結果顯示:綿羊iPS細胞中多能性基因的表達水平極顯著高于成纖維細胞(P< 0.01)。圖3
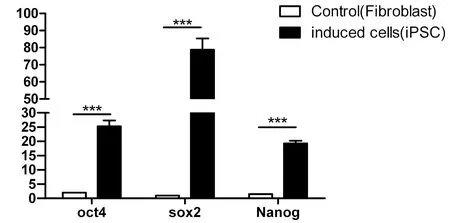
圖3 OiPSCs的Oct4、Sox2、Nanog基因的qPCR檢測
Fig.3 The quantitative PCR analysis of Oct4, Sox2 and Nanog in OiPSCs
2.4 轉錄組測序
2.4.1 不同誘導時期細胞RNA-Seq轉錄組數據拼接
統計誘導0、10、20和30 d Unigenes數量,分別獲得55 795、47 416、48 069和48 927條Unigenes序列。
2.4.2 不同誘導時期差異基因的表達
不同誘導時期差異基因的表達水平(圖4A-B)。以誘導0 d的細胞作為對照,誘導30 d的細胞共獲得9 465個差異表達基因,其中上調基因6 987個,下調基因2 478個。利用Blast2GO軟件,按照參與的細胞組分(Cellular Component)、分子功能(Molecular Function)和生物過程(Biological Process)(圖4C)對30和0 d的差異基因進行分類注釋。經比對共有5 374個差異基因對比到GO數據上。在參與的生物學過程中,細胞過程(cellular process)所占比例最多,占12.6%,其次為單體過程(single-organism process)和代謝過程(metabolic process),分別占10.5%和10.4%;在細胞成分(cell component)分類中,細胞組分(cell part)所占比例最多,占18.7%,其次是細胞(cell)和細胞器(organelle),分別占18.6%和15.9%;在分子功能(molecular function)分類中,分子綁定(binding)所占比例最高,占49.3%,其次是催化活性(catalytic activity)和酶調節活性(enzyme regulator activity),分別占25.6%和4.5%。圖4
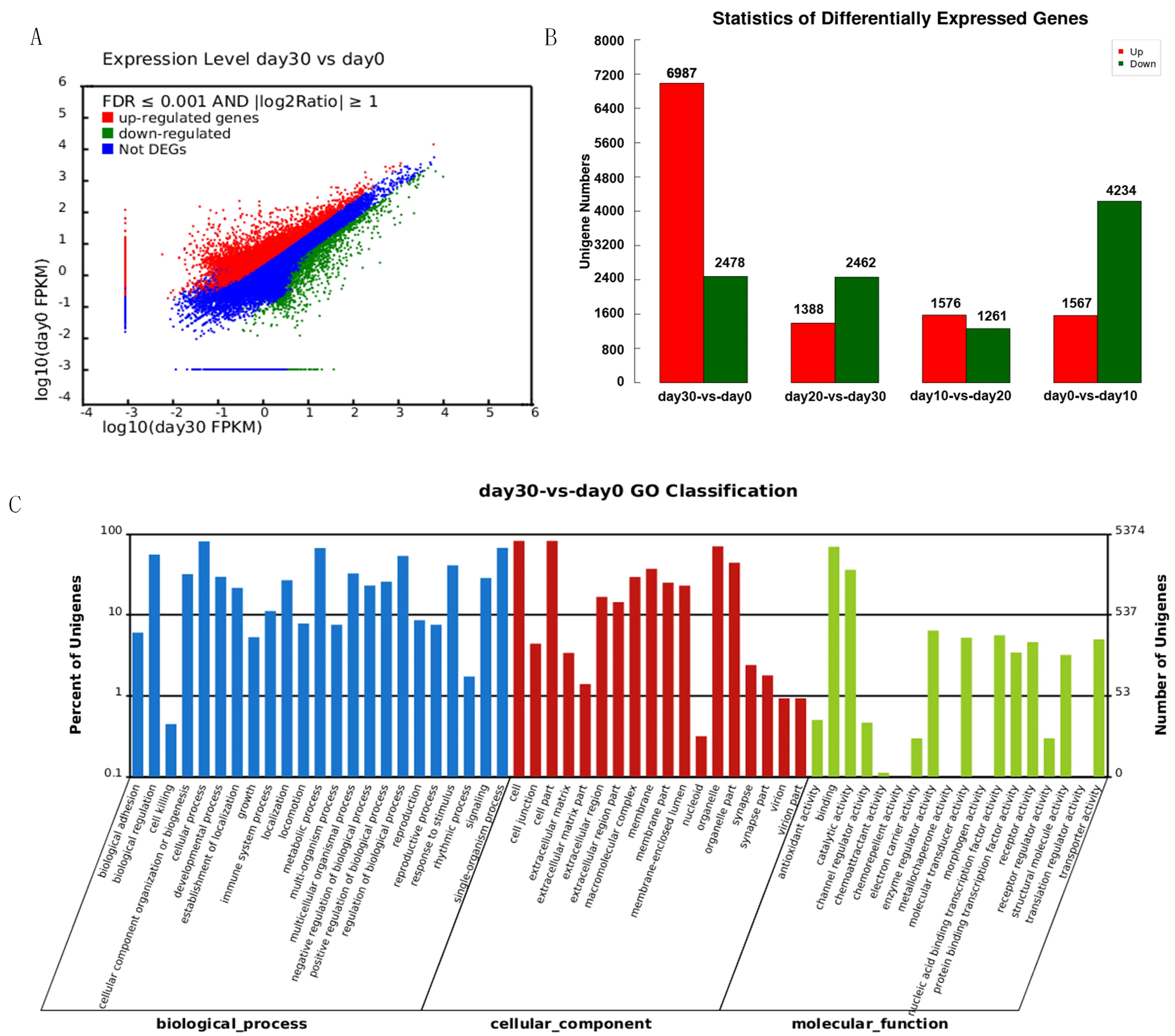
圖4 30和0 d細胞中基因差異表達
Fig.4 Different UniGene expression profiles between 30 cell and 0 d cell
2.4.3 部分干性相關基因誘導0和30 d表達量差異性比較
利用Cufflinks軟件,計算每個Unigene在各個樣本中的表達量FPKM值,FPKM法能消除基因長度和測序量差異對計算基因表達的影響,計算得到的基因差異表達量可直接用于比較不同樣本間的基因表達差異。l-Myc、Sox2、Oct4在綿羊成纖維細胞重編程過程中基因表達量均呈上調趨勢,其中,l-myc和Sox2基因表達量差異性顯著,Oct4基因差異不顯著。圖5
2.4.4 誘導30和0 d細胞差異表達基因的KEGG富集
將已注釋的差異表達基因與KEGG數據庫進行比對,共有5 773個基因比對到KEGG數據庫,共涉及到259條通路,其中富集通路最多的是剪接體通路,其次是卵母細胞成熟分裂通路和P53信號通路,說明這三條通路在綿羊多潛能干細胞誘導過程中最活躍。表1

圖5 RNA-Seq分析綿羊成纖維細胞重編程的基因表達
Fig.5 Gene analyses in reprogramming of Ovis fibroblasts using RNA-seq表1 差異表達基因KEGG代謝途徑分類
Table 1 KEGG classification of differentially expressed genes
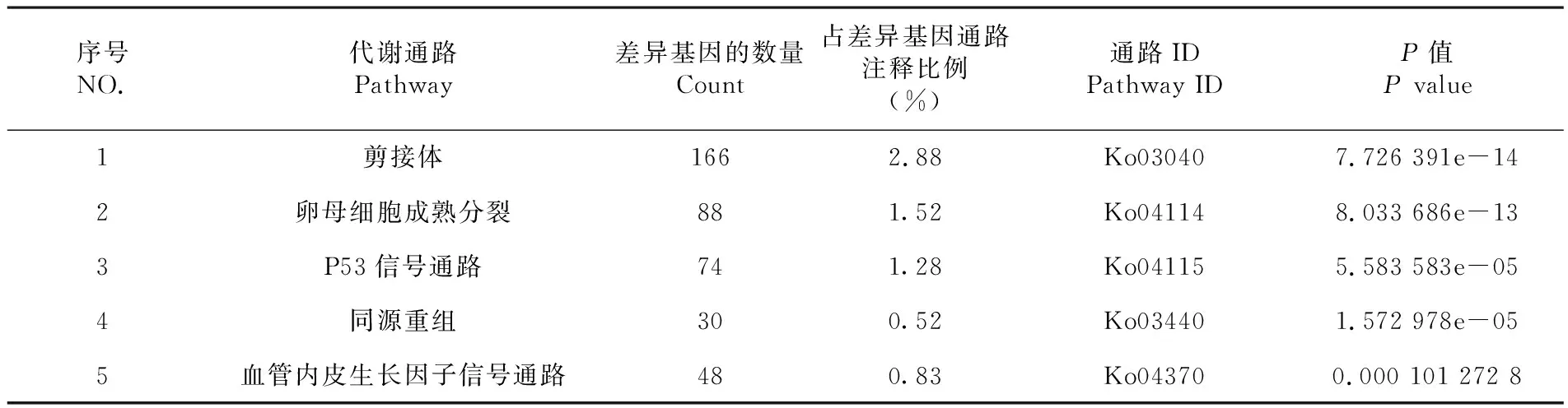
序號NO.代謝通路Pathway差異基因的數量Count占差異基因通路注釋比例(%)通路IDPathway IDP值P value1剪接體1662.88Ko030407.726 391e-142卵母細胞成熟分裂881.52Ko041148.033 686e-133P53信號通路741.28Ko041155.583 583e-054同源重組300.52Ko034401.572 978e-055血管內皮生長因子信號通路480.83Ko043700.000 101 272 8
3 討 論
誘導性多能干細胞是經體細胞重編程形成的,但重編程的分子調控機制至今尚未完全清楚,尤其是在大動物的iPSCs的研究略有滯后。在綿羊iPSCs誘導技術方面最為成熟的是iPS細胞誘導分化成三胚層,并且能檢測到iPSCs的標記。Li等[14]成功將慢病毒介導的Oct4、Sox2、Klf4、c-Myc四因子轉入綿羊成纖維細胞,培養14 d后出現類似人胚胎干細胞樣形態的克隆,經堿性磷酸酶染色、畸胎瘤三胚層組織學鑒定均證明是干細胞。Bao等[5]將四環素誘導慢病毒系統介導的Oct4、Sox2、Klf4、c-Myc四個轉錄因子,同時補充添加Nanog、Lin28、SV40 large T和人端粒酶反轉錄酶hTERT轉染至胎羊成纖維細胞,用Knockout血清替代品(Knockout serum replacement, KSR)代替胎牛血清培養細胞,11~20 d出現克隆,形態類似小鼠胚胎干細胞,經堿性磷酸酶染色,Oct4、 Sox2、 Nanog、SSEA-1、Tra1-60以及Tra-1-81這些標記物染色都為陽性。另有研究表明,盡管Oct4、Sox2、Klf4、c-Myc這四個限定因子具有高度的保守性,但iPSCs的形態、增殖能力和相應的分子機制存在種屬差異。譬如,人和小鼠的iPS細胞形態和自我更新的分子機制具有明顯的差異。與人和小鼠相比,綿羊iPSCs較難獲得[9]。一般認為,Oct3/4 和 Sox2 是誘導多能性所需的核心因子,而Klf4 和 c-Myc則能提高Oct3/4和Sox2的誘導效率[15]。Esteban等[16]認為,四因子誘導法中,去除c-Myc可獲得較好的iPS,建系成功率也顯著提高。Nakagawa等[17]研究表明,去除c-Myc誘導iPS細胞時,誘導時間較長且效率較低,但特異性強。這說明c-Myc的作用可能是促進細胞的增殖而不是細胞的重編程[18]。研究采用Oct4、Sox2、Klf4、l-Myc、lin28五因子誘導法,結果顯示,在其他條件一致的情況下,選擇質粒濃度質量比1∶1∶1時,更有利于形成AP+克隆[19]。常見的質粒轉染方法包含病毒載體法、脂質體法和電轉法等,鑒于電轉保護劑既能降低細胞死亡率,又能提高電轉效率,試驗采取電轉法并且使用KSR代替胎牛血清培養綿羊iPS細胞,細胞傳代明顯增強,可傳至第4代。
細胞表面標記能有效識別和區分多能干細胞,是鑒定多能干細胞重要的工具。SSEAs作為一種表面標記物被廣泛用于監測胚胎干細胞的多能性,它包括SSEA-1、SSEA-3、SSEA-4。 Dattena等[20]報道,綿羊胚胎干細胞表達SSEA-1、SSEA-3、SSEA-4,Li等[14]研究顯示,綿羊iPSCs只表達Oct4、Sox2、Nanog、SSEA-4,未表達SSEA-1和SSEA-3;而試驗顯示綿羊iPSCs只表達Sox2、Oct4、SSEA-1,部分轉染的基因表現沉默。這可能與不同實驗室采用的誘導方案及誘導產生iPSCs重編程程度有關,也有可能是其他一些因素影響了細胞狀態,尚需進一步的試驗佐證。Chin等[21]通過比較不同代數人iPS細胞和人胚胎干細胞的基因表達圖譜,結果顯示,代數高的人iPS細胞(第54~61代)比代數低的人iPS細胞(第5~9代)更接近于胚胎干細胞。由此推測,持續培養的過程對完善iPS細胞重編程意義重大。從試驗實時熒光定量PCR檢測結果來看,綿羊iPS細胞中多能性基因Oct4,Sox2,Nanog的表達量均顯著增高,和RNA-Seq的結果相一致,初步證實了獲得的iPS細胞具有一定的多能性。
比較綿羊iPSCs和用于重編程的成纖維細胞之間基因的表達情況對于了解綿羊多能性干細胞誘導的分子過程是非常重要的。高通量轉錄組測序技術為揭示研究動、植物基因表達及調控發揮了重要作用,尤其在側重了解動物發育某一方面的基因表達,挖掘差異表達基因備受關注[22]。研究利用高通量測序技術構建了誘導0、10、20和30 d細胞的轉錄組文庫,獲得了大量基因信息。重點針對誘導30和0 d的陽性細胞測序結果分析發現,篩選到的差異表達基因顯著富集在卵母細胞成熟分裂、同源重組、P53信號通路等通路上。其中,顯著富集的P53信號通路參與細胞的生長調控過程。已有研究證實,P53基因的突變和刪除與一系列腫瘤相關聯;P53基因編碼的蛋白在細胞損傷的情況下可誘導激活下游的P21蛋白,觸發DNA修復和細胞凋亡,P53~P21蛋白通路能抑制干細胞的分化和增殖,因此,抑制P53基因的表達可提高重編程的效率[23]。研究表明,敲除P53蛋白的和ASF1A蛋白能夠提高綿羊腎臟細胞重編程的效率[24]。有趣的是,不同于成熟細胞中的低水平P53,這種蛋白在ESC和iPSCs中呈高豐度表達[25]。研究結果進一步證實了綿羊成纖維細胞在向著iPSC轉變。
4 結 論
利用電轉法將Oct4、Sox2、Klf4、l-Myc、Lin28五個轉錄因子導入至綿羊成纖維細胞中,經AP染色、細胞免疫熒光及熒光定量PCR檢測,細胞均呈陽性,且表達Oct4、Sox2、SSEA-1等多種ES細胞的多能性標記基因;轉錄組測序結果獲得了大量的綿羊候選功能基因,經生物信息學分析發現,差異表達基因主要涉及細胞組分、分子結合、細胞過程等諸多生理生化過程,同時參與多種信號通路,其中,P53信號通路顯著富集,且其在干細胞增殖方面起著非常重要的作用,獲得了綿羊誘導性多能干細胞。
參考文獻(References)
[1] Molofsky, A. V. , Pardal, R. , & Morrison, S. J. . (2004). Diverse mechanisms regulate stem cell self-renewal.CurrentOpinioninCellBiology, 16(6): 700-707.
[2] Odorico, J. S. , Kaufman, D. S. , & Thomson, J. A. (2001). Multilineage differentiation from human embryonic stem cell lines.StemCells, 19(3): 193-204.
[3] Takahashi, K., Tanabe, K., Ohnuki, M., Ichisaka, T., Tomoda, K. & Yamanaka, S. (2007). Induction of pluripotent stem cells from adult human fibroblasts by defined factors.Cell, 131(5): 861-872,
[4] Yu, J. , Vodyanik, M. A. , Smuga-Otto, K. , Antosiewicz-Bourget, J. , Frane, J. L. , & Tian, S. , et al. (2008). Induced pluripotent stem cell lines derived from human somatic cells.Obstetrical&GynecologicalSurvey, 63(3): 154-155.
[5] Zhao, X. Y. , Li, W. , Lv, Z. , Liu, L. , Tong, M. , & Hai, T. , et al. (2010). Ips cells produce viable mice through tetraploid complementation.ChinaBasicScience,461(7260): 86-90.
[6] Liao, J. , Cui, C. , Chen, S. , Ren, J. , Chen, J. , & Gao, Y. , et al. (2009). Generation of induced pluripotent stem cell lines from adult rat cells.CellStemCell, 4(1):11-15.
[7] Liu H; Zhu F; Yong J; Zhang P; Hou P; Li H; Jiang W; Cai J; Liu M; Cui K; Qu X; Xiang T; Lu D; Chi X; Gao G; Ji W; Ding M; Deng H. (2008). Generation of induced pluripotent stem cells from adult rhesus monkey fibroblasts.CellStemCell, 3(6): 587-590.
[8] Wu, Z. , Chen, J. , Ren, J. , Bao, L. , Liao, J. , & Cui, C. , et al. (2009). Generation of pig induced pluripotent stem cells with a drug-inducible system.JournalofMolecularCellBiology, 1(1): 46-54.
[9] Bao, L. , He, L. , Chen, J. , Wu, Z. , Liao, J. , & Rao, L. , et al. (2011). Reprogramming of ovine adult fibroblasts to pluripotency via drug-inducible expression of defined factors.CellResearch, 21(4): 600-608.
[10] Takahashi, K., Okita, K., Nakagawa, M., & Yamanaka, S. (2006). Induction of pluripotent stem cells from fibroblast cultures.NatureProtocols, 51(15): 3,081-3,089.
[11] Liu, J. , Balehosur, D. , Murray, B. , Kelly, J. M. , Sumer, H. , & Verma, P. J. . (2012). Generation and characterization of reprogrammed sheep induced pluripotent stem cells.Theriogenology, 77(2): 338-346.
[12] Sartori, C. , Didomenico, A. I. , Thomson, A. J. , Milne, E. , & Whitelaw, C. B. A. . (2012). Ovine-induced pluripotent stem cells can contribute to chimeric lambs.CellularReprogramming, 14(1): 8-19.
[13]王峰,劉燦,潘傳英.大家畜誘導多潛能干細胞(iPSCs)研究進展[J]. 農業生物技術學報,2014, 22(10): 1 286-1 297.
WANG Feng, LIU Can, PAN Chuang-ying. (2014). Progress of induced pluripotent stem cells (iPSCs) of big domestic animals [J].JournalofAgriculturalBiotechnology, 22(10):1,286-1,297. (in Chinese)
[14] Li, Y. , Cang, M. , Lee, A. S. , Zhang, K. , & Liu, D. . (2011). Reprogramming of sheep fibroblasts into pluripotency under a drug-inducible expression of mouse-derived defined factors.PLOSONE, 6(1): e15947:1-8.
[15]陳倩,史慶華. iPS細胞的遺傳安全性[J]. 遺傳,2012, 34(3): 260-268.
CHEN Qian, SHI Qing-hua. (2012). The genetic safety of induced pluripotent stem (ips) cells[J].Hereditas, 34(3): 260-268. (in Chinese)
[16] Esteban, M. A. , Wang, T. , Qin, B. M. , Yang, J. Y. , Qin, D. J. , & Cai, J. L. , et al. (2010). Vitamin c enhances the generation of mouse and human induced pluripotent stem cells.CellStemCell, 6(1): 71-79.
[17] Nakagawa, M. , Koyanagi, M. , Tanabe, K. , Takahashi, K. , Ichisaka, T. , & Aoi, T. , et al. (2008). Generation of induced pluripotent stem cells without myc from mouse and human fibroblasts.NatureBiotechnology, 26(1): 101-106.
[18] Pera, M. F. , & Hasegawa, K. . (2008). Simpler and safer cell reprogramming.NatureBiotechnology, 26(1): 59-60.
[19]王聰慧,趙帥,張譯元,等.電穿孔轉染制備綿羊誘導多潛能干細胞條件的優化[J].西北農業學報,2015, 24(9): 9-15.
WANG Cong-hui, ZHAO Shuai, ZHANG Yi-yuan, et al. (2015). Optimization of inducing pluripotent stem cells by 0; electroporation in sheep [J].ActaAgriculturaeBoreali-occidentalissinica, 24(9): 9-15. (in Chinese)
[20] Dattena, M. , Chessa, B. , Lacerenza, D. , Accardo, C. , & Cappai, P. . (2006). Isolation, culture, and characterization of embryonic cell lines from vitrified sheep blastocysts.MolecularReproductionandDevelopment, 73(1): 31-39.
[21] Chin, M. H. , Mason, M. J. , Xie, W. , Volinia, S. , Singer, M. , & Peterson, C. , et al. (2009). Induced pluripotent stem cells and embryonic stem cells are distinguished by gene expression signatures.CellStemCell, 5(1):111-123.
[22]朱志明,陳紅萍,林如龍,等.山麻鴨開產期和產蛋高峰期卵巢組織轉錄組分析[J].中國農業科學,2016, 49(5): 998-1 007.
ZHU Zhi-ming, CHEN Hong-ping, LIN Ru-long, et al. (2016). Transcriptome analysis of ovary tissue in early laying period and egg laying peak period of Shanma ducks [J].ScientiaAgriculturaSinica, 49(5): 998-1,007. (in Chinese)
[23] Bao, X. , Wu, H. , Zhu, X. , Guo, X. , Hutchins, A. P. , & Luo, Z. , et al. (2015). The p53-induced lincrna-p21 derails somatic cell reprogramming by sustaining h3k9me3 and cpg methylation at pluripotency gene promoters.CellResearch, 25(1): 80-92.
[24] Shi H, Fu Q, Li G, Ren Y, Hu S, Ni W, Guo F, Shi M, Meng L, Zhang H, Qiao J, Guo Z, Chen C. (2015). Roles of p53 and asf1a in the reprogramming of sheep kidney cells to pluripotent cells.CellularReprogramming, 17(6): 441-452.
[25] Lee, D. F. , Su, J. , Ang, Y. S. , Carvajalvergara, X. , Muleronavarro, S. , & Pereira, C. F. , et al. (2012). Regulation of embryonic and induced pluripotency by aurora kinase-p53 signaling.CellStemCell, 11(2): 179-194.
