熱激因子AtHsfA1a突變對熱脅迫中擬南芥抗壞血酸過氧化物酶的影響
2018-02-28 03:18:35郭麗紅曾潔媛
西南農業學報 2018年1期
關鍵詞:研究
郭麗紅, 李 念, 徐 亞, 曾潔媛
(1.昆明學院云南省高校特色生物資源開發與利用重點實驗室,云南 昆明 650214;2.昆明學院農學院,云南 昆明 650214)
【研究意義】在自然界中各種農作物遭受各種逆境脅迫時表達一系列防御基因,產生相應的耐逆境生理、生化途徑,從而獲得耐逆境能力。擬南芥熱激因子AtHsfA1a是與耐逆境有關的主要轉錄調控因子,抗壞血酸過氧化物酶(APX,EC1.11.1.7)是在逆境中參與抗逆性形成的重要抗氧化酶,進一步鑒定熱激因子AtHsfA1a與APX基因的關系,對于揭示熱激因子AtHsfA1a生理功能的作用機制具有重要的意義。【前人研究進展】植物逆境信號轉導的研究發現,轉錄因子在調控防御基因表達中起重要作用。植物熱激因子(heat shock transcription factors,HSFs)是真核生物熱激反應的主要轉錄調控因子[ 1-2 ]。正常生長條件下,HSF是以非活性單體的形式存在,逆境脅迫誘導HSF形成有活性的三體,而與靶基因啟動子區的熱激元件(heat shock element, HSE)特異性結合,以調控靶基因的轉錄表達,從而控制生理功能[3-5 ]。在擬南芥中存在21個熱激因子的同源體,組成熱激因子家族[ 4]。雖然目前對絕大多數同源體的功能還不了解,但研究顯示其中熱激因子AtHsfA1a是與耐逆境有關的主要轉錄因子[ 5-7 ]。研究也發現各種逆境脅迫均可誘發細胞內活性氧濃度的增加而導致氧化脅迫,最終導致植物死亡[ 8-10 ]。然而植物細胞中的抗氧化酶系統可以把細胞內產生的具有很強氧化活性的活性氧直接或間接地清除,保障了細胞內各種生命代謝活動的正常進行[ 11-13 ]。目前關于熱激因子與抗氧化酶關系的研究主要集中在抗壞血酸過氧化物酶(APX,EC1.11.1.7)上,Storozhenko等研究發現熱脅迫可以誘導Apx1基因表達[14 ],Panchuk,et al.于2002年研究發現擬南芥過量表達HSF3可以使熱脅迫下的抗氧化酶Apx活性高于野生型[ 15 ],且出現新的同工酶,Apx的mRNA水平也高于野生型。研究還發現Apx的啟動子包括熱激因子HSF的結合位點[15 ]。然而,AtHsfA1a 對APX的影響研究不多,并且在逆境中,由于逆境信號的級聯反應,很難區分HSFs直接與靶DNA結合以調控抗APX基因的表達,還是由處理逆境引起細胞產生的其它反應而導致APX基因的表達。【本研究切入點】本研究以T-DNA 插入AtHsfA1a基因突變擬南芥及野生型植株為材料,比較熱脅迫下不同基因型植株的抗壞血酸過氧化物酶(APX)活性的變化,從生理水平揭示細胞內AtHsfA1a突變對APX活性的影響;為了研究AtHsfA1a 對APX影響的分子機制,采用RT-PCR分析熱脅迫不同基因型植株APX基因的表達水平,采用染色質免疫沉淀技術和凝膠阻滯電泳研究AtHsfA1a與APX基因啟動子區的體內外結合情況。【擬解決的關鍵問題】從生理生化和分子水平鑒定AtHsfA1a與APX的關系,為揭示熱激因子AtHsfA1a在逆境中的作用機制和生理功能范圍提供理論依據。
1 材料與方法
1.1 植物材料
擬南芥哥侖比亞種(Arabidopsisthaliana,ecotype Columbia),包括野生型(簡稱WT)和T-DNA 插入AtHsfA1a基因突變擬南芥(SALK-068042, 購于美國生物資源中心ABRC, 簡稱MT )。
1.2 方法
1.2.1 材料栽培及高溫處理 種子用0.1 %的HgCl2和70 %的乙醇進行滅菌處理后,接種于1/2 MS固體培養基[15]中,置于25 ℃光照下萌發。待種子長出4片真葉,把小苗移至土壤中,用保鮮膜遮蓋,在生長室培養3 d后去膜,恒溫培養。取生長4周的小苗進行高溫處理:將洗凈后的擬南芥植株置于SIB Buffer(0.5 mmol/L K2HPO4,0.5 mmol/L KH2PO4,1 % 蔗糖,pH 6.0) 中,42 ℃高溫處理10 min。對照實驗的處理溫度為25 ℃。
1.2.2 抗壞血酸過氧化物酶活性的測定 取葉片0.5 g于預冷的研缽中,加1 mL預冷的提取液(50 mmol/L Tris-HCl緩沖液,pH 值 7.0,含20% 甘油,5 mmol/L MgCl2,1 mmol/L EDTA,1 mmol/L ASA,1 mmol/L DTT,1 mmol/L GSH)冰浴研磨成勻漿,于4 ℃下15 000 r/min離心30 min,取上清液備用。在25 ℃水浴中預熱30 min反應混合液 (50 mmol/LTris-HCl 緩沖液,pH 7.0,內含0.1 mmol/L EDTA)2.890 mL。加入10 mmol/L H2O230 μl,酶液50 μl,最后加入30 mmol/L ASA 50 μl以啟動反應,終體積為3 mL。每隔10 s讀出OD290的減少值來計算酶活性。
1.2.3 抗氧化酶基因的mRNA的測定 總RNA提取采用QIAGEN RNA提取試劑盒,用 1.5 %的凝膠電泳(120V)15 min,在凝膠成像儀下檢測總RNA。反轉錄合成cDNA按照下列配方:RNA/mRNA(8 μl)、Trans criptTmRT/RI Enzyme Mix(1 μl)、Anehored Oligo(dT)18(1 μl)、2×TS Reaction Mix(10 μl),RNase-free Water to 20 μl,輕輕混勻后,在水浴鍋中42 ℃孵育30 min后,在85 ℃的水浴鍋中加熱5 min失活Trans ScriptTmRT。反轉錄產物作為PCR的模板,以APX啟動子片段為引物(APX-F: 5’-CGGCGTTATTATCGTCAG-3’,APX-R:5’-AAGCAGGAGTGAGCTACAGA-3’)進行PCR擴增, 以Actin-F: 5'-TTGTCACACACAAGTGCATCAT-3'; Actin-R: 5'- AAGCTGGGGTTTTATGAATGG-3'為對照。程序如下: 94 ℃ 5 min,94 ℃ 30 s,62 ℃ 30 s,72 ℃ 30 s,72 ℃10 min,35個循環,擴增后的產物用1.5 %瓊脂糖凝膠電泳檢測。
1.2.4 染色質免疫沉淀技術(CHIP)分析AtHsfA1a與APX基因啟動子區的結合 按照Guo LH (2008)等方法[ 5 ],將上述逆境處理后的小苗,在室溫下浸入crossing buffer 中(0.4 mmol/L sucrose, 10 mmol/L Tris-HCl(pH 8.0), 1 mmol/L EDTA, 1 mmol/L PMSF, 1 % formaldehyde),在真空下培養15 min后 加入終濃度為100 mmol/L甘氨酸在真空下培養5 min以終止交聯反應。小苗用無菌去離子水沖洗后,按照Guo,et al.的方法,利用AtHsfA1a專一性抗體,進行免疫沉淀,所獲得的沉淀DNA,用APX基因啟動子區的引物進行PCR擴增,擴增后的產物用1.5 %瓊脂糖凝膠電泳檢測。
1.2.5 凝膠阻滯電泳(EMSA)分析AtHsfA1a與APX基因啟動子區的結合 凝膠阻滯電泳(EMSA)按照Guo LH (2008)等方法[ 5 ],40 ng AtHsfA1a 與 40 fmol 生物素標記的HSE-探針在25 ℃ 共培養 20 min后用凝膠阻滯電泳分析。在競爭實驗中, 加入獲得的APX基因啟動子區片段。以標準牛血清蛋白(BSA)作為負對照,用于AtHsfA1a結合片段的特異性研究。
2 結果與分析
2.1 熱激因子AtHsfA1a突變對擬南芥幼苗在熱脅迫中APX酶活性的影響
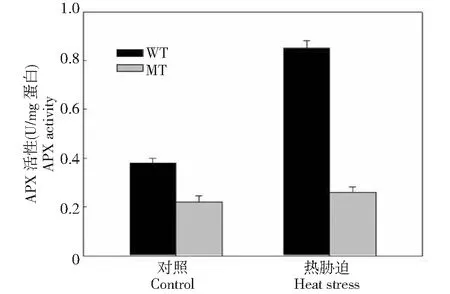
WT:野生型;MT:T-DNA插入突變型WT: Wild-type plants; MT: T-DNA inserted mutant of Arabidopsis AtHsfA1a圖1 不同基因型擬南芥植株在熱脅迫下APX的活性變化Fig.1 Activity of APX in different genotypes of Arabidopsis thaliana plants under heat stress

WT:野生型;MT:T-DNA插入突變型 WT: Wild-type plants;MT: T-DNA inserted mutant of Arabidopsis AtHsfA1a 圖2 不同基因型擬南芥植株在熱脅迫下APX的表達情況Fig.2 Expression of APX in different genotypes of Arabidopsis thaliana plants under heat stress
為了從生理水平揭示細胞內AtHsfA1a 對APX活性的影響,測定熱脅迫下AtHsfA1a基因突變擬南芥和野生型植株中APX酶活性變化,結果如圖1所示,在對照環境中(25 ℃)下,2種基因型植株的APX的活性差異不大,但在熱脅迫42 ℃下處理10 min后,野生型植株中的活性升高較大,但AtHsfA1a基因突變擬南芥活性的變化沒有野生型的明顯,說明擬南芥AtHsfA1a突變對熱脅迫中APX活性有影響。
2.2 熱激因子AtHsfA1a突變對擬南芥幼苗在熱脅迫中APX表達的影響
為了研究AtHsfA1a 對APX影響的分子機制,采用RT-PCR法比較熱脅迫下不同基因型植株中APX的表達量,以肌動蛋白(Actin)為內參,從圖2可知,野生型的APX基因42 ℃下的表達量高于25 ℃的表達量,且野生型的APX的表達量在2種溫度下均比AtHsfA1a基因突變植株的高,說明在熱脅迫下AtHsfA1a突變能夠影響抗氧化酶APX基因的表達。
2.3 CHIP體內鑒定AtHsfA1a與APX的基因啟動子區的結合狀況
為了研究AtHsfA1a在體內的表達水平是如何影響抗氧化酶APX基因的表達,應用染色質免疫沉淀技術(Chip)體內篩選AtHsfA1a的靶DNA,以AtHsfA1a的靶DNA為模板,以APX基因啟動子區域設計引物進行Promotor-specific-PCR擴增,體內鑒定AtHsfA1a與抗氧化酶的基因啟動子區的結合狀況,其結果如圖3。
從圖3可以知道,在熱脅迫下,以抗氧化酶APX基因啟動子區的片段為引物,野生型植株有擴增產物,而AtHsfA1a基因突變擬南芥植株無擴增條帶,說明AtHsfA1a在體內能與抗氧化酶APX的基因啟動子區的結合,影響抗氧化酶APX的表達。
2.4 EMSA體外鑒定AtHsfA1a與APX的基因啟動子區的結合狀況
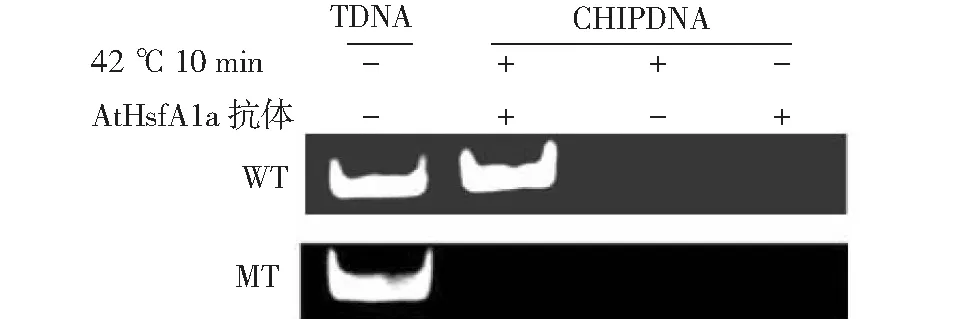
T DNA:總DNA;CHIP DNA:染色質免疫沉淀DNA;WT:野生型,MT:T-DNA插入突變型T DNA:Total DNA;IP DNA:Chromatin immunoprecipitation DNA; WT: Wild-type plants, MT: T-DNA inserted mutant of Arabidopsis AtHsfA1a圖3 CHIP體內驗證AtHsfA1a與抗氧化酶APX的基因啟動子區的體內結合狀況Fig.3 The binding of AtHsfA1a to the promoter regions of APX investigated by using chromatin immunoprecipitation

HSE* :生物素標記的標準的熱激元件;BSA:標準牛血清蛋白HSE* :Biotin labeled perfect heat shock element;BSA:Standard bovine serum albumin圖4 EMSA體外驗證AtHsfA1a與抗氧化酶APX的基因啟動子區片段的結合狀況Fig.4 The binding of AtHsfA1a to the promoter regions of APX investigated by using electrophoretic mobility shift assay
為了研究AtHsfA1a對抗氧化酶APX基因的表達影響是直接的還是間接的,設計含有熱激元件(HSE)的保守片段(GAATTC)進行人工合成,生物素標記合成的DNA片段。選擇染色質免疫沉淀技術(Chip)體內篩選出的能被APX啟動子區引物擴增片段為競爭性片段,標準牛血清蛋白(BSA)用于AtHsfA1a結合片段的特異性進行研究。采用電泳遷移率變動分析(EMSA) 體外鑒定AtHsfA1a與APX的啟動子區域的特異性片段的結合情況,結果如圖4。從圖4可知,在EMSA中染色質免疫沉淀技術(Chip)體內篩選出APX1基因啟動子區片段能特異性地與熱激元件競爭結合AtHsfA1a,體外驗證了AtHsfA1a直接與APX的基因啟動子區的結合,進一步說明了AtHsfA1a對APX的影響是直接的。
3 討 論
植物對高溫抵抗能力的變化與細胞生理生化的一系列適應性變化有關。熱脅迫損傷細胞膜系統并導致氧化脅迫,抗氧化酶系統的增強與提高植物的抗逆性密切相關[ 8-10 ]。前期研究均表明抗氧化酶APX在逆境中可以通過活性升高或表達量的增加而提高抗逆性。擬南芥HSF是一個大的蛋白質家族,研究發現,擬南芥HSF除調控熱激蛋白表達外,也可調控一些非熱激蛋白的表達,例如擬南芥HSF3可以在熱脅迫下可以提高抗氧化酶Apx活性,同時Apx的mRNA水平也提高[15],研究還發現Apx的啟動子包括熱激因子HSF的結合位點[14],說明擬南芥HSF家族與APX密切相關。然而,擬南芥AtHsfA1a 與APX的關系研究不多。研究者前期利用熱激因子AtHsfA1a基因沉默植株研究發現AtHsfA1a對APX有影響。本研究采用T-DNA 插入AtHsfA1a基因突變擬南芥植株的目的是該植株由于T-DNA 插入導致AtHsfA1a基因的無法正常表達,研究結果表明T-DNA 插入AtHsfA1a基因突變植株的APX活性要比野生型植株中的活性弱,從生理水平說明體內AtHsfA1a與APX活性有關。關于AtHsfA1a是直接調控APX的活性,還是調控APX的表達需要進一步研究。為了從分子水平研究AtHsfA1a與APX表達的關系,本研究采用RT-PCR分析熱脅迫不同基因型植株APX基因的表達水平,結果表明AtHsfA1a基因突變植株中的APX的mRNA的相對量低于野生型,說明AtHsfA1a可以誘導APX的表達。雖然目前研究顯示逆境誘導熱激因子調控抗氧化酶APX基因的表達;然而很難區分HSFs直接與靶DNA結合以調控抗氧化酶基因的表達,還是由處理逆境引起細胞產生的其它反應而影響APX的表達。由于染色質免疫沉淀技術可以找出在生理條件下轉錄因子與靶DNA序列的結合位點,從而反映體內基因表達調控的真實情況,而凝膠阻滯電泳 (EMSA)又稱為電泳遷移率變動分析,它是一種在體外證實目的蛋白與DNA直接結合的方法[5]。因此采用了染色質免疫沉淀技術和凝膠阻滯電泳研究AtHsfA1a與APX基因啟動子區片段的結合情況,體內和體外鑒定APX是否是熱激因子直接調控的下游靶基因。結果表明,在熱脅迫下AtHsfA1a基因突變擬南芥植株中未篩選出APX基因啟動子區片段,而野生型篩選出APX基因啟動子區片段,且AtHsfA1a與APX基因啟動子區的結合是直接的,初步說明AtHsfA1a對APX表達的調控是直接的。
4 結 論
熱激因子AtHsfA1a突變對熱脅迫中擬南芥APX的影響是從轉錄調控水平直接影響APX的表達,這將為揭示熱激因子AtHsfA1a在逆境中的作用機制和生理功能范圍提供理論依據。
[1]Wunderlich M, Doll J, Busch W, et al. Heat shock factors: regulators of early and late functions in plant stress response[J].Plant Stress, 2007(1):16-22.
[2]Akerfelt M, Morimoto R I, Sistonen L. Heat shock factors: integrators of cell stress, development and lifespan[J].Molecular Cell Biology, 2010,11:545-555.
[3]von Koskull-D?ring P, Scharf K, Nover L. The diversity of plant heat stress transcription factors[J]. Trends Plant Sci, 2007, 12: 452-457.
[4]Nover L. Arabidopsis and the heat stress transcription factor world: how many heat stress transcription factors do we need?[J]. Cell Stress Chaperones, 2001(6):177-189.
[5]Guo LH, Chen S N, Liu KH, et al. Isolation of heat shock factor HsfA1a-binding sites in vivo revealed variations of heat shock elements in Arabidopsis thaliana[J]. Plant Cell Physiol, 2008, 49:1306-1315.
[6]Liu Y, Zhang C, Chen J, et al. Arabidopsis heat shock factor HsfA1a directly senses heat stress, pH changes, and hydrogen peroxide via the engagement of redox state[J]. Plant Physiology and Biochemistry, 2013, 64: 92-98.
[7]Liu H, Liao H, Charng Y. The role of class A1 heat shock factors (HSFA1s) in response to heat and other stresses inArabidopsis[J]. Plant Cell Environ, 2011, 34:738-751.
[8]Imlay J A. Pathway of oxidative damage[J]. Annu Rev Microbiol, 2003,57:395-418.
[9]Mittler H. Reactive oxygen gene network of plants[J]. Trends in Plant Science, 2004(9): 490-498.
[10]Foyer C H, Noctor G. Oxidant and antioxidant signaling in plants: a reevaluation of the concept of oxidative stress in a physiological context[J]. Plant Cell Environ, 2005, 28:1056-1071.
[11]Busch W, Wunderlich M, Sch?ffl F. Identification of novel heat shock factor-dependent genes and biochemical pathways inArabidopsisthaliana[J]. The Plant Journal, 2005, 41:1-14.
[12]Davletova S, Rizhsky L, Liang H,et al. Cytosolic ascorbate peroxidase 1 is a central component of the reactive oxygen gene network of Arabidopsis[J]. Plant Cell, 2005, 17: 268-281.
[13]周 靜,曾學英,賀 維,等.低溫脅迫下藍花楹的耐寒生理機制分析[J].西南農業學報,2016,29:74-80.
[14]Storozhenko S. The heat-shock element is a functional component of the Arabidopsis APX.I gene promoter[J].Plant Physiol, 2005,118: 1005-1014.
[15]Panchuk I I, Volkov R A, Schoffl F. Heat stress and heat shock transcription factor-dependent expression and activity of ascorbate peroxidase in Arabidopsis[J]. Plant Physiol, 2002, 29: 838-853.
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
