鞘內注射TRESK過表達腺病毒抑制JNK活化和神經元凋亡減輕神經病理性疼痛
2018-02-28 02:06:32熊艷峰林文靜林森黃振興黃騰王漢兵楊承祥仲吉英周俊
實用醫學雜志 2018年1期
熊艷峰 林文靜 林森 黃振興 黃騰 王漢兵 楊承祥 仲吉英 周俊
佛山市第一人民醫院麻醉科(廣東佛山 528000)
神經病理性疼痛(neuropathic pain,NP)是由神經系統原發性損害和功能障礙所激發或引起的一種慢性疼痛,發病機制尚未完全闡明。離子通道改變形成異常放電的是NP痛覺異常主要原因,雙孔鉀離子通道TRESK(TWIK?related spinal cord potassium channels)在背根神經節(dorsal root gan?glion,DRG)中大量表達,TRESK下調可能參與多種慢性疼痛的病理生理過程[1-4],本課題組的前期研究發現上調大鼠DRG的TRESK表達可明顯減輕NP[5],但具體機制尚不清楚。最近研究表明,神經元的凋亡與NP密切相關[6],而c?Jun 氨基末端激酶(Jun N?terminal kinase,JNK)是MAPK通路的重要信號通路,在細胞周期、凋亡等多種生理病理過程中發揮重要作用,已被證明與NP密切相關[7]。本研究通過鞘內注射TRESK基因重組腺病毒載體:pAd/CMV/V5?DEST?TRESK[8],上調NP大鼠DRG的TRESK表達,觀察其對NP大鼠疼痛行為學以及DRG的JNK磷酸化和神經元凋亡的影響,進一步闡明TRESK介導的神經元凋亡參與NP的機制,為NP提供新的治療靶點。
1 材料與方法
1.1 動物選擇及分組 雄性SD大鼠72只,體質量200~250 g,由廣東省佛山市第一人民醫院實驗動物中心提供。采用隨機數字表法,將其分為6組(n=12):空白對照組(C組)、假手術組(S組)、神經病理性疼痛組(NP組)、TRESK過表達腺病毒組(T組)、陰性腺病毒組(V組)和生理鹽水組(NS組)。NP組、T組、NS組和V組采用坐骨神經分支選擇性損傷(SNI)大鼠模型。T組、V組和NS組于造模后分別在鞘內注射TRESK基因重組腺病毒(pAd/CMV/V5?DEST?TRESK)、陰性腺病毒和等容量生理鹽水。
1.2 SNI模型的建立 NP組、T組、NS組和V組大鼠均取左側行SNI模型[9];S組手術操作與NP組相同,但不結扎神經。各組大鼠術后腹腔注射青霉素20萬U預防感染。
1.3 鞘內注射 在大鼠的L4,5椎間隙進行鞘內注射:T組大鼠鞘內注射TRESK基因重組腺病毒pAd/CMV/V5?DEST?TRESK(109IU/mL),V 組大鼠鞘內注射陰性腺病毒,劑量均為25 μL;NS組大鼠鞘內注射生理鹽水。
1.4 機械性縮足反射閾值(MWT)和熱縮足潛伏期(TWL)測定 各組大鼠分別于術前1 d(BL)和術后1、3、7和14 d(D1、D3、D7、D14)分別測定左后肢MWT和TWL。用Von?Frey纖維絲(Stoelting公司,美國)測定MWT,測量5次,每次間隔10 min,取均值,該值即為機械痛閾值[10]。BME?40A型熱痛刺激儀(中國醫學科學院生物工程研究所)以熱輻射法測定TWL,測量5次,每次間隔10 min[11]。
1.5 Western blot檢測大鼠DRG的TRESK、p?JNK、JNK和Caspase3蛋白表達 于造模后7 d(D7)痛閾測定結束后,每組斷頭處死6只大鼠,分別取L4~5節段背根神經節組織;造模后14 d痛閾測定結束,每組斷頭處死剩余6只大鼠,分別取L4~5節段脊髓組織。組織均于-80℃保存,Western blot分別檢測大鼠DRG的TRESK、Caspase3、p?JNK和JNK蛋白表達。
1.6 DRG凋亡細胞檢測 TUNEL標記法檢測DRG凋亡神經元細胞。造模后14 d(D14)痛閾測定結束,每組各取6只大鼠,10%水合氯醛麻醉下剖胸暴露心臟,經心尖部插管灌流固定,依次灌注生理鹽水、4%多聚甲醛,取L4~5DRG組織置于30%多聚甲醛中固定24 h,常規石蠟包埋,橫斷5 μm連續切片。TUNEL染色后二甲苯透明中性樹脂封片,光鏡下觀察并TUNEL陽性細胞(細胞核棕褐色染色)。每只大鼠取5張DRG切片,用奧林巴斯倒置顯微鏡(Olympus C7070wz,日本)觀察,應用圖像分析儀分析并計算細胞凋亡率(%)=凋亡細胞數/總細胞數×100%。
1.7 統計學方法 用SPSS 10.0統計軟件進行統計分析。計量資料以表示,組間比較采用方差分析。P<0.05為差異有統計學意義。
2 結果
2.1 MWT與TWL比較 各組各時點的TWL組間比較差異無統計學意義(P>0.05);NP組、T組、V組、NS組D1、D3、D7、D14時的MWT明顯低于C組、S組(P<0.05);與NP組、V組、NS組相比,T組D1、D3、D7、D14時的MWT的明顯升高(P< 0.05);T組、V組、NS組D1、D3、D7、D14時的MWT組間比較差異無統計學意義(P>0.05,表1)。
2.2 各組大鼠DRG的TRESK蛋白表達比較 與C組、S組相比,NP組、V組和NS組L4~5術側DRG的TRESK蛋白表達明顯降低(P<0.05);TRESK組L4~5術側DRG的TRESK蛋白表達明顯高于NP組、V組、NS組(P<0.05);T組、V組、NS組的TRESK蛋白表達組間比較差異無統計學意義(P>0.05,表2)。
表1 各組大鼠不同時點PWT和TWL的比較Tab.1 Comparation of PWT and TWL in each group rats at different time points ±s

表1 各組大鼠不同時點PWT和TWL的比較Tab.1 Comparation of PWT and TWL in each group rats at different time points ±s
注:與C、S組比較,aP <0.05;與NP、V、NS組比較,bP <0.05
MWT(g)TWL(s)組別C組S組NP組T組V組NS組BL 13.3±1.3 13.4±1.8 13.6±1.6 13.7±1.2 13.8±1.4 13.0±1.7 D1 13.4±1.0 13.9±1.9 8.5±1.1a 10.7±1.3ab 8.7±1.3a 8.4±1.0a D3 13.2±1.5 13.5±1.2 6.2±1.0a 8.8±1.2ab 6.0±0.8a 6.1±1.0a D7 13.2±1.0 13.5±1.3 5.1±0.9a 8.0±1.6ab 5.0±0.8a 5.2±1.0a D14 13.6±1.3 13.3±0.8 4.9±1.1a 8.5±1.3ab 4.8±0.8a 5.0±1.1a BL 17.7±2.0 17.6±1.6 17.5±2.1 18.0±2.1 17.9±2.1 17.3±2.0 D1 17.9±2.0 17.7±2.0 17.8±1.8 17.5±1.6 17.8±1.7 17.4±2.0 D3 17.3±1.6 17.5±2.2 17.3±2.0 17.8±1.3 17.3±1.2 17.6±1.6 D7 17.5±2.0 17.1±1.6 17.6±2.1 18.2±2.3 17.9±2.0 17.4±1.3 D14 17.7±2.2 17.0±1.8 17.2±1.8 17.3±2.2 17.7±2.4 17.0±1.8
表2 各組大鼠背根神經節組織TRESK蛋白表達的比較(n=6)Tab.2 Comparation of TRESK expression in DRG of each group rats ±s

表2 各組大鼠背根神經節組織TRESK蛋白表達的比較(n=6)Tab.2 Comparation of TRESK expression in DRG of each group rats ±s
注:與C、S組比較,aP <0.05;與NP、V、NS組比較,bP <0.05
指標C組S組NP組T組V組NS組TRESK蛋白1.34±0.11 1.41±0.13 0.41±0.08a 1.05±0.21b 0.37±0.15a 0.44±0.16a
表3 各組大鼠DRG的JNK磷酸化水平比較(n=6)Tab.3 Comparation of JNK phosphorylation in DRG of each group rats ±s

表3 各組大鼠DRG的JNK磷酸化水平比較(n=6)Tab.3 Comparation of JNK phosphorylation in DRG of each group rats ±s
注:與C、S組比較,aP < 0.05;與NP、V、NS組比較,bP <0.05
指標C組S組NP組T組V組NS組p?JNK/JNK 0.13±0.04 0.15±0.05 1.21±0.13a 0.76±0.08ab 1.32±0.15a 1.26±0.12a
表4 各組大鼠DRG神經元凋亡比較(n=6)Tab.4 Comparation of neuronal apoptosis in DRG of each group rats ±s
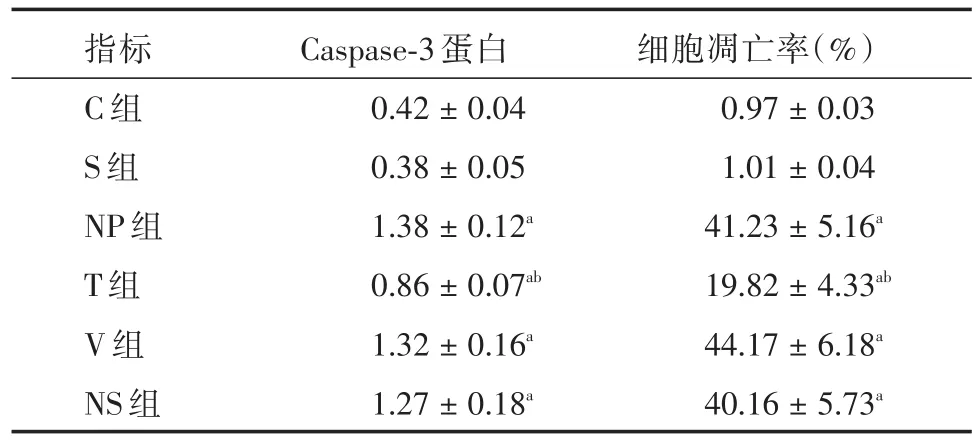
表4 各組大鼠DRG神經元凋亡比較(n=6)Tab.4 Comparation of neuronal apoptosis in DRG of each group rats ±s
注:與C、S組比較,aP < 0.05;與NP、V、NS組比較,bP <0.05
指標C組S組NP組T組V組NS組Caspase?3蛋白0.42±0.04 0.38±0.05 1.38±0.12a 0.86±0.07ab 1.32±0.16a 1.27±0.18a細胞凋亡率(%)0.97±0.03 1.01±0.04 41.23±5.16a 19.82±4.33ab 44.17±6.18a 40.16±5.73a
2.3 各組大鼠DRG的JNK磷酸化比較 與C組、S組相比,NP組、T組、V組和NS組L4~5DRG的JNK磷酸化水平明顯增高(P<0.05);與NP組、V組、NS組相比,T組L4~5DRG的JNK磷酸化水平明顯降低(P< 0.05);T組、V組、NS組的L4~5DRG的JNK磷酸化水平組間比較差異無統計學意義(P>0.05),見表3。
2.4 各組大鼠DRG神經元凋亡比較 與C組、S組相比,NP組、T組、V組和NS組L4~5DRG的細胞凋亡率和Caspase?3表達明顯增高(P< 0.05);與NP組、V組、NS組相比,T組L4~5DRG的細胞凋亡率和Caspase?3表達明顯降低(P< 0.05);T組、V組、NS組的L4~5DRG的細胞凋亡率和Caspase?3表達組間比較差異無統計學意義(P>0.05,表4)。
3 討論
SNI動物模型的特點是具有顯著異常機械痛敏,但無熱痛閾變化。該模型更接近NP的臨床特征,是一種較理想的NP模型。本研究制備SNI模型可重復性高,與前期研究行為學一致[5]:各組大鼠均在術后1 d即開始表現為機械痛敏,而并不表現熱痛超敏,符合SNI模型典型的行為學特征。
雙孔鉀離子通道亞型TRESK在大鼠DRG大量表達,主要功能是維持神經元細胞的靜息電位,在神經元細胞的興奮過程中起重要作用。本課題組前期研究構建的TRESK基因重組腺病毒載體pAd/CMV/V5?DEST?TRESK,已證明在離體和在體均具有可靠的轉染效率和上調TRESK目的基因的效果[11-12]。本試驗所有實施SNI手術的組別TRESK蛋白均下調,而給予pAd/CMV/V5?DEST?TRESK的TRESK大鼠背根神經節的TRESK的表達明顯上調。另外,與NP組比較,T組的MWT升高,再次驗證了上調SNI大鼠DRG的TRESK表達可減輕大鼠NP。
NP脊髓水平的神經元的凋亡、炎性反應,已被多個研究證實在NP中發揮重要作用,多型NP均伴有背根神經節和脊髓神經元的凋亡[13],其機制可能為:抑制性中間神經元的凋亡減少了其對傷害性信息傳遞的抑制作用,導致背根神經節向脊髓傳導的痛覺傷害性神經元興奮增高,從而產生痛覺過敏[14]。而JNK是介導MAPK磷酸化的重要信號通路,也介導了炎癥反應和凋亡的過程,因此,筆者推測JNK與NP之間有聯系。同時,已有學者認為脊髓水平的MAPK磷酸化通過調節炎性反應和神經元凋亡參與了慢性疼痛的機制[15]。本研究觀察了SNI大鼠TRESK下調對DRG凋亡神經元的影響,結果SNI大鼠伴隨著DRG的TRESK下調,凋亡細胞和凋亡相關蛋白Caspase?3明顯上調,而上調DRG的TRESK表達后,SNI大鼠的凋亡明顯減輕,說明神經元的凋亡介導了NPDRG的TRESK機制,也證實TRESK是下游通路,通過介導JNK信號來發揮作用。
細胞內鉀離子外流是細胞凋亡的重要特征之一。在許多病理條件下,胞內鉀離子濃度降低可以引起神經元凋亡,鉀通道阻斷劑或胞外高鉀可以阻斷神經元凋亡。鉀離子通道TRESK是DRG最主要的背景鉀離子通道,DRG的TRESK下調改變神經元興奮性,通過突觸傳遞等方式將疼痛信號傳遞到中樞,參與多型疼痛的發病機制。JNK在細胞周期、凋亡等多種生理病理過程中發揮重要作用,與NP密切相關[6],而TRESK參與的慢性疼痛機制也與JNK、神經元凋亡有相關性。結合本試驗中DRG的JNK磷酸化結果,筆者推測在NP發病機制中,DRG的TRESK通過JNK磷酸化引起神經元凋亡,從而介導了痛敏的發生。
綜上所述,TRESK介導大鼠的神經病理性痛機制可能與背根神經節JNK磷酸化和神經元凋亡有關。本實驗驗證了K+通路TRESK在NP中發揮了重要的作用,并且其介導了下游通路。在今后的臨床工作中,通過特異性離子通道抑制劑、信號下游通道的抑制劑或激動拮抗劑來達到治療NP的效果,也是今后的研究方向[16]。
[1]周俊,姚尚龍,楊承祥,等.神經病理性痛大鼠背根神經節TRESK mRNA表達的變化[J].中華麻醉學雜志,2011,31(2):183?185.
[2]ASI C,REVITAL S,EREZ S,et al.Pain?associated signals,acidosis and lysophosphatidic acid,modulate the neuronal K2P2.1 channel[J].Mol Cell Neurosci,2009,40(3):382?389.
[3]CZIRJAK G,ENYEDI P.Targeting of calcineurin to an NFAT?like docking site is required for the calcium?dependent activa?tion of the background K+channel,TRESK[J].J Biol Chem,2006,281(21):14677?14682.
[4]LAFRENIèRE R G,CADER M Z,POULIN J F,et al.A dom?inant?negative mutation in the TRESK potassium channel is linked to familial migraine with aura[J].Nat Med,2010,16(10):1157?1160.
[5]ZHOU J,YAO S L,YANG C X,et al.Intrathecal TRESK gene recombinant adenovirus attenuates Spared nerve injury?in?duced neuropathic pain in rats.International Journal of Molecu?lar Medicine[J].Neuro Report,2013,24(3):131?136.
[6]MENG C,LIANG X,LI Q,et al.Changes of GTP cyclohydro?lase I and neuronal apoptosis in rat spinal dorsal cord induced by sciatic nerve injury[J].Neurol Scie,2013,34(12):2145?2150.
[7]GAO Y J,ZHANG L,SAMAD O A,et al.JNK?induced MCP?1 production in spinal cord astrocytes contributes to central sen?sitization and neuropathic pain[J].J Neurosci,2009,29(13):4096?4108.
[8]周俊,姚尚龍,楊承祥,等.大鼠TRESK基因重組腺病毒載體的構建[J].中華麻醉學雜志,2011,31(3):296?298.
[9]DECOSTERD I,WOOLF C J.Spared nerve injury:an animal model of persistent peripheral neuropathic pain[J].Pain,2000,87(2):149?158.
[10]CHAPLAN S R,BACH F W,POGREL J W,et al.Quantita?tive assessment of tactile allodynia in the rat paw[J].J Neuro?sci Methods,1994,53(1):55?63.
[11]HARGREAVES K,DUBNER R,BROWN F,et al.A new and sensitive method for measuring thermal nociception in cutane?ous hyperalgesia[J].Pain,1998,32(1):77?88.
[12]ZHOU J,YAO S L,YANG C X,et al.TRESK gene recombi?nant adenovirus vector inhibits capsaicin?mediated substance P release from cultured rat dorsal root ganglion neurons[J].Mol Med Rep,2012,5(4):1049?1052.
[13]SEKIGUCHI M,SEKIGUCHI Y,KONNO S,et al.Compari?son of neuropathic pain and neuronal apoptosis following nerve root or spinal nerve compression[J].Eur Spine J,2009,18(12):1978?1985.
[14]MAO J,SUNG B,JI R R,et al.Neuronal apoptosis associated with morphine tolerance:evidence for an opioid?induced neuro?toxic mechanism[J].J Neurosci,2002,22(17):7650?7661.
[15]OBATA K,YAMANAKA H,DAI Y,et al.Differential activa?tion of extracellular signal?regulated protein kinase in primary afferent neurons regulates brain?derived neurotrophic factor ex?pression after peripheral inflammation and nerve injury[J].J Neurosci,2003,23(10):4117?4126.
[16]縱雪梅,曹磊,許鐵.SP600125?JNK特異性抑制劑對大鼠腦缺血再灌注神經元損傷的保護作用及其作用[J].實用醫學雜志,2011,27(14):2529?2532.
