小麥分蘗成穗數相關分子標記在重組自交系(RIL)群體中的有效性驗證及實用性評價
2018-03-05 05:39:33胡洋山湯穎子晏本菊任正隆任天恒
麥類作物學報 2018年1期
胡洋山,湯穎子,李 治,晏本菊,任正隆,任天恒
(1.四川農業大學生命科學學院,四川雅安 625014; 2.四川農業大學農學院,四川成都 611130)
分蘗成穗是小麥(TriticumaestivumL.)重要的生物學特性,單位面積穗數作為產量三要素之一受到育種家的廣泛關注[1]。而單位面積最大分蘗數直接影響單位面積穗數,與其具有極顯著相關性,同樣對小麥的最終產量有較大的影響[2]。四川省屬于我國西南麥區,是典型的雨養農業區,較多的陰天和較少的光照會導致小麥分蘗成穗力弱,單位面積穗數減少,進而造成產量下降[3-4]。本課題組選育的多穗型系列小麥新品種川農12、川農17和川農18在西南麥區種植比例日益增大,這些新品種具有較強的分蘗成穗力,增產潛力高,為打破中國西南小麥生態區“生態穗容量”的限制提供了新的思路[5]。
小麥單位面積最大分蘗數和單位面積穗數是非常復雜的數量性狀,受多基因控制且易受到環境影響。目前,有三個控制小麥分蘗的基因( tin1、 tin2、 tin3)分別被定位在1A、2A和3A染色體上[6-9]。Xu等[10]利用高分蘗和矮桿突變體構建的RIL群體,在2D染色體上檢測到一個控制分蘗的主效QTL。Shah等[11]在3A染色體上檢測到一個控制成穗的QTL,且該QTL同時控制株高和穗粒數,為一因多效QTL。迄今,國內外研究者使用不同的小麥群體,將控制小麥分蘗成穗的QTL定位在1A、1B、2A、2B、2D、3A、3B、3D、4D、5A、5D、6D、7A和7B等多個染色體上,并開發出較多的分子標記[12-18]。但是這些與基因和QTL位點連鎖的分子標記大多具有品種特異性,只在特定的環境或遺傳背景下才有效,且多數并未實際應用到育種進程中。因此,這些已報道的分子標記在西南麥區的有效性和實用性有待進一步驗證。目前,分子標記的有效性和實用性越來越受到育種家的關注。馬 麗等[19]通過對7個已報道的小麥穗發芽抗性相關分子標記與表型的相關性研究,從中篩選出3個實用性好的分子標記。張兆萍等[20]選用4個小麥穗發芽抗性相關分子標記研究其與表型的相關性,篩選出2個可用于鑒定穗發芽抗性的分子標記。王金萍等[21]通過分析159份玉米自交系的分子標記驗證結果和莖腐病田間表型的符合度,篩選出4個可用于抗莖腐病檢測的分子標記。但是與小麥重要產量因素分蘗成穗相關的分子標記驗證尚未見報道。本研究利用6個已發表的分子標記對以多穗型小麥品種川農18和新品系T1208構建的包含371個株系的重組自交系群體進行PCR擴增,驗證其在該群體中的有效性,并評估其實用性,以期為西南麥區小麥優質育種提供參考。
1 材料與方法
1.1 小麥材料及田間種植
供試材料為小麥品種川農18與新品系T1208構建的重組自交系(F11及F12),包含371個株系,各株系農藝性狀表型基本穩定,適合進一步研究。川農18的單位面積(每平方米,下同)最大分蘗數和穗數分別為450.33和341.67,T1208的單位面積最大分蘗數和穗數分別為404.67和277.00,以上數據為兩年平均值。該群體于2014-2015年度和2015-2016年度種植在四川農業大學小麥育種基地(四川省邛崍市,30°25′N,103°28′E)。采用隨機區組設計,小區行長2 m,每行20株,行距0.25 m,每個株系種植4行,3次重復。栽培管理同大田標準化管理方法。
1.2 分蘗成穗數的調查
播種85 d后調查最大分蘗數,播種157 d后調查穗數[22]。從每個小區第2行和第3行的固定位置選取20株調查最大分蘗數和穗數,3次重復的平均值作為每個株系最大分蘗數和穗數的表型值,然后計算單位面積(m2)最大分蘗數和穗數(觀測值為每0.5 m2的最大分蘗數和穗數,乘以2得出單位面積最大分蘗數和穗數)。
1.3 基因組DNA的提取及PCR擴增
每個株系隨機選取10粒種子進行發芽、栽培,于三葉期提取基因組DNA[23]。PCR反應體系為25 μL,包含2 μL(50 ng· μL-1)基因組DNA、2.5 μL 10× buffer(由天根生化科技有限公司生產)、0.5 μL 10×dNTPs(由上海博彩生物科技有限公司生產)、1 μL上下游引物(表1,由擎科梓熙生物技術有限公司合成)、0.2 μLTaq酶(5 U·μL-1,由天根生化科技有限公司生產)和17.8 μL ddH2O。PCR反應程序為:94 ℃預變性4 min;94 ℃變性30 s,50~65 ℃復性30 s,72 ℃延伸30 s,35次循環;72 ℃延伸7 min。PCR產物用8%非變性聚丙烯酰胺凝膠電泳檢測,在恒定功率55 W下分離約45 min后銀染[24],最后照相并統計帶型。

表1 本研究所用的分蘗成穗相關分子標記Table 1 Molecular markers related to tiller number and spike number in the present study
1.4 數據統計分析
數據統計分析采用軟件SPSS 22.0和Microsoft Excel 2016進行。等位基因與分蘗成穗相關性分析參照馬 麗等[19]和張海萍等[25]的方法,即對等位基因進行賦值,將在某位點含有該等位基因的株系賦值為“1”,不含該等位基因的株系賦值為“0”,利用Pearson相關分析模型評價每個等位基因與單位面積最大分蘗數和穗數的相關性,進而驗證分子標記的有效性。單位面積最大分蘗數和穗數均取兩年平均值。
2 結果與分析
2.1 單位面積最大分蘗數和穗數及其相關性
通過田間調查和計算,371份供試材料的單位面積最大分蘗數為254.00~700.67,變異系數為22.28%;單位面積穗數為197.67~391.67,變異系數為14.12%。將單位面積最大分蘗數小于350定為低分蘗,350~500定為中分蘗,大于500定為高分蘗。371個株系中,低分蘗組有86個株系,中分蘗組有202個株系,高分蘗組有83個株系。將單位面積穗數小于250定為低成穗,250~300定為中成穗,大于300定為高成穗。371個株系中,低成穗組有54個株系,中成穗組有191個株系,高成穗組有126個株系。相關性分析結果表明,單位面積最大分蘗數與穗數極顯著正相關,相關系數為0.792(P<0.001)。
2.2 分蘗成穗相關分子標記的有效性驗證結果
2.2.1 分子標記Xgwm264的有效性
分子標記Xgwm264在371份供試材料中共擴增出四種類型的片段,其中,A型含有大小為180 bp和250 bp的2個條帶;B型含有大小為120 bp和175 bp的2個條帶;C型含有大小為150 bp左右的2個條帶;D型含有大小為120 bp、180 bp和250 bp的3個條帶(圖1)。擴增出A型條帶的材料共31份,占總數的8.36%;擴增出B型條帶的材料共185份,占總數的49.87%;擴增出C型條帶的材料共143份,占總數的38.54%;擴增出D型條帶的材料共12份,占總數的3.23%。B和C分別為親本T1208和川農18的帶型。從與分蘗、穗數的相關性來看(表2),擴增出A型條帶的材料與分蘗、穗數無顯著相關性;B型條帶與分蘗數和穗數均呈極顯著負相關;C型條帶與分蘗數和穗數均呈極顯著正相關;D型條帶與分蘗數呈極顯著正相關,但與穗數無顯著相關性。說明能擴增出B型條帶的材料單位面積最大分蘗數和穗數較低,能擴增出C型條帶的材料單位面積最大分蘗數和穗數較高,該分子標記可用于供試材料分蘗數和穗數的篩選鑒定。
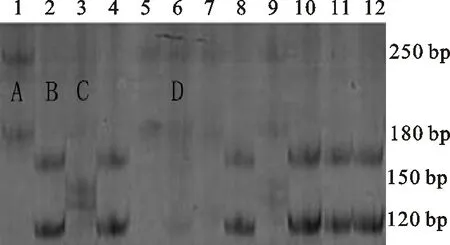
1~12為RIL群體的株系編號;A、B、C、D代表不同類型的擴增條帶,其中,B和C分別為親本T1208和川農18的帶型。
1-12 represent tested lines of RILs;A,B,C and D represent the different types of the amplified bands,among which,B and C were the same as that amplified from the parents T1208 and Chuannong 18,respectively.
圖1 分子標記Xgwm264擴增出的片段類型
*和**分別代表在0.05和0.01水平上顯著相關。
* and ** indicate significant correlation at 0.05 and 0.01 levels,respectively.
2.2.2 分子標記Xbarc232的有效性
用分子標記Xbarc232在371份供試材料中共擴增出兩種類型的片段,其中,A型不含有大小為175 bp的條帶;B型含有大小為175 bp的條帶(圖2)。擴增出A型條帶的材料共201份,占總數的54.18%;擴增出B型條帶的材料共170份,占總數的45.82%。A和B分別為親本T1208和川農18的帶型。從與分蘗、穗數的相關性分析可以看出(表2),A型條帶和B型條帶均與分蘗數和穗數無顯著關系,說明該分子標記不適合用于分蘗成穗數的篩選鑒定。
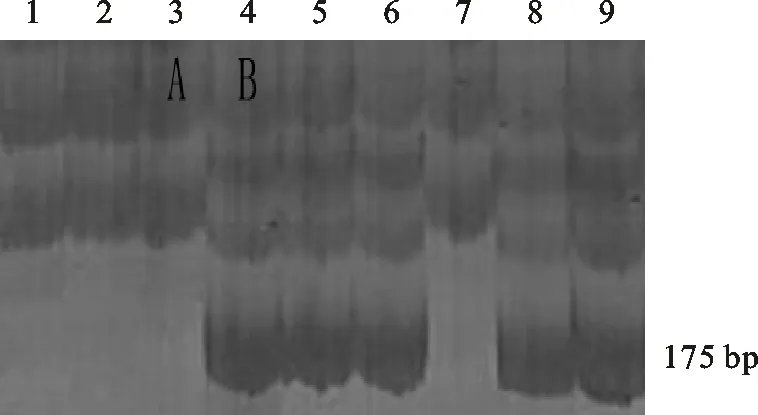
1~9為RIL群體的株系編號;A和B代表不同類型的擴增條帶,A和B也分別為親本T1208和川農18的帶型。
1-9 represent tested lines of RILs;A and B represent the different types of the amplified bands,which were the same as that amplified from the parents T1208 and Chuannong 18,respectively.
圖2分子標記Xbarc232擴增出的片段類型
Fig.2PCRfragmentsamplifiedwiththemolecularmarkerXbarc232
2.2.3 分子標記Xwmc169的有效性
用分子標記Xwmc169在371份供試材料中共擴增出三種類型的片段,其中,A型不含有大小為150 bp和170 bp的條帶;B型含有大小為150 bp的條帶;C型含有大小為150 bp和170 bp的條帶(圖3)。擴增出A型條帶的材料共66份,占總數的17.79%;擴增出B型條帶的材料共269份,占總數的72.51%;擴增出C型條帶的材料共36份,占總數的9.70%。A和B分別為親本T1208和川農18的帶型。從與分蘗、穗數的相關性分析可以看出(表2),A型條帶與分蘗數和穗數均呈極顯著正相關;B型條帶與分蘗數和穗數均呈極顯著負相關;C型條帶與分蘗數和穗數均無顯著相關。說明能擴增出A型條帶的材料單位面積最大分蘗數和穗數較高,能擴增出B型條帶的材料單位面積最大分蘗數和穗數較低,該分子標記可用于供試材料分蘗數和穗數的篩選鑒定。
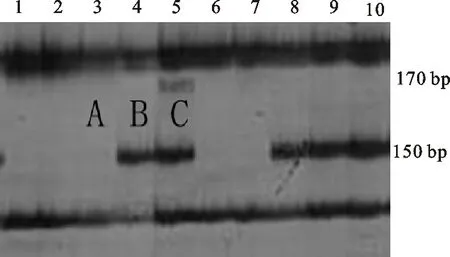
1~10為RIL群體的株系編號;A、B和C代表不同類型的擴增條帶,其中,A和B分別為親本T1208和川農18的帶型。
1-10 represent tested lines of RILs;A,B and C represent the different types of the amplified bands,among which,A and B were the same as that amplified from the parents T1208 and Chuannong 18,respectively.
圖3分子標記Xwmc169擴增出的片段類型
Fig.3PCRfragmentsamplifiedwiththemolecularmarkerXwmc169
2.2.4 分子標記Xwmc215的有效性
用分子標記Xwmc215在371份供試材料中共擴增出三種類型的片段,其中,A型含有大小為230 bp和375 bp的條帶;B型含有大小為235 bp的條帶,且不含有大小為375 bp和230 bp的條帶;C型含有大小為230 bp和235 bp的條帶(圖4)。擴增出A型條帶的材料共204份,占總數的54.99%;擴增出B型條帶的材料共160份,占總數的43.13%;擴增出C型條帶的材料共7份,占總數的1.88%。A和B分別為親本T1208和川農18的帶型。從與分蘗、穗數的相關性分析可以看出(表2),能擴增出A型條帶的材料與分蘗數呈極顯著正相關,但與穗數無顯著相關;B型條帶與分蘗數呈極顯著負相關,但與穗數無顯著相關;C型條帶與分蘗數和穗數均無顯著相關。因此該分子標記僅可用于供試材料分蘗數的鑒定篩選。

1~7為RIL群體的株系編號;A、B和C代表不同類型的擴增條帶,其中,A和B分別為親本T1208和川農18的帶型。
1-7 represent tested lines of RILs,A,B and C represent the different types of the amplified bands,among which,A and B were the same as that amplified from the parents T1208 and Chuannong 18,respectively.
圖4分子標記Xwmc215擴增出的片段類型
Fig.4PCRfragmentsamplifiedwiththemolecularmarkerXwmc215
2.2.5 分子標記Xgwm437的有效性
用分子標記Xgwm437在371份供試材料中共擴增出五種類型的片段,其中,A型含有大小為130 bp和250 bp的條帶;B型含有大小為190 bp和250 bp的條帶;C型含有大小為210 bp和250 bp的條帶;D型含有大小為130 bp的條帶;E型含有大小為210 bp的條帶(圖5)。擴增出A型條帶的材料共102份,占總數的27.49%;擴增出B型條帶的材料共12份,占總數的3.23%;擴增出C型條帶的材料共225份,占總數的60.65%;擴增出D型條帶的材料共10份,占總數的2.70%;擴增出E型條帶的材料共22份,占總數的5.93%。A和C分別為親本T1208和川農18的帶型。從與分蘗、穗數的相關性分析可以看出(表2),A型條帶與分蘗數呈極顯著正相關,但與穗數無顯著相關;B、C、D型條帶與分蘗數和穗數均無顯著相關;E型條帶與分蘗數和穗數均呈極顯著負相關。說明能擴增出A型條帶的材料單位面積最大分蘗數較高,能擴增出E型條帶的材料單位面積最大分蘗數較低,該標記可用于供試材料分蘗數的篩選鑒定。僅有E型條帶與單位面積穗數具有相關性,因此該分子標記能否用于穗數的篩選鑒定還有待進一步驗證。
2.2.6 分子標記Xcfd23的有效性
用分子標記 Xcfd23在371份供試材料中共擴增出三種類型片段,其中,A型含有大小100 bp的條帶;B型含有大小125 bp的條帶;C型含有大小為100 bp和125 bp的條帶(圖6)。擴增出A型條帶的材料共136份,占總數的36.66%;擴增出B型條帶的材料共14份,占總數的3.77%;擴增出C型條帶的材料共221份,占總數的59.57%。A和C分別為親本T1208和川農18的帶型。從與分蘗、穗數的相關性來看(表2),擴增出A型條帶的材料與分蘗數呈極顯著負相關,但與穗數無顯著相關性;擴增出B型條帶的材料與分蘗數無顯著相關性,與穗數呈顯著正相關;擴增出C型條帶的材料與分蘗數呈極顯著正相關,但與穗數無顯著相關性。說明能擴增出A型條帶的材料分蘗數較低,能擴增出C型條帶的材料分蘗數較高,該標記可用于供試材料分蘗數的篩選鑒定。僅B型條帶與穗數具有相關性,因此該分子標記能否用于穗數的篩選鑒定還有待進一步驗證。
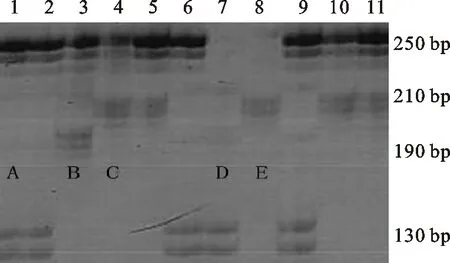
1~11為RIL群體株系編號;A、B、C、D和E代表不同類型的擴增條帶,其中,A和C分別為親本T1208和川農18的帶型。
1-11 represent tested lines of RILs,A,B,C,D and E indicated different types of amplified bands,among which,A and C were the same as that amplified from the parents T1208 and Chuannong 18,respectively.
圖5分子標記Xgwm437擴增出的片段類型
Fig.5PCRfragmentsamplifiedwiththemolecularmarkerXgwm437

1~10為RIL群體株系編號;A、B和C代表不同類型的擴增條帶,其中,A和C分別為親本T1208和川農18的帶型。
1-10 represent tested lines of RILs,A,B and C indicated different types of amplified bands,among which,A and C were the same as that amplified from the parents T1208 and Chuannong 18,respectively.
圖6分子標記Xcfd23擴增出的片段類型
Fig.6PCRfragmentsamplifiedwiththemolecularmarkerXcfd23
3 討 論
3.1 單位面積最大分蘗數和穗數的相關性及多效性基因
高產是育種家們追求的主要目標,小麥單位面積穗數作為“產量三要素”之一,是非常重要的農藝性狀,而單位面積最大分蘗數又直接決定了單位面積穗數。本研究中,單位面積最大分蘗數與單位面積穗數的相關性達到極顯著水平(P<0.001),這也與前人的研究結果相一致[15]。前人研究表明,小麥穗數的廣義遺傳力高達0.88,分蘗數的廣義遺傳力也達到0.63[2,26]。這說明通過育種手段,從品種本身提高分蘗數和穗數進而提高產量是非常有效的方法。單位面積最大分蘗數與單位面積穗數是一對具有顯著相關性的性狀,受到多效性基因的控制,Li等[15]報道了一個位于6D染色體同時控制分蘗數和穗數的一因多效QTL。多效性是基因較為常見的屬性,在遺傳、進化、衰老、發育和疾病等多個方面都有出現[26]。在育種進程中,具有多效性的基因往往更加有利于穩定化選擇[27]。與多效性基因或QTL所連鎖的分子標記具有更高的實際應用價值。本研究選取了6個與單位面積最大分蘗數或單位面積穗數相關的分子標記,在本課題組所構建的RIL群體中進行有效性驗證,該群體來源于親本多穗型小麥品種川農18和1BL/1RS新品系T1208,川農18具有較強的分蘗成穗力,其單位面積最大分蘗數和單位面積穗數均明顯高于T1208。選用某一性狀差異較大的兩親本所構建的群體,更有利于對該性狀進行QTL定位,同樣也更有利于分子標記的有效性驗證[28]。
3.2 分子標記有效性驗證的意義
小麥大量性狀的QTL定位已見報道,但已報道的小麥QTL位點大多僅在一個或兩個環境中被檢測出,與QTL位點緊密連鎖的分子標記則缺乏通用性。楊 燕等[29-30]在95份中國地方小麥品種和歷史小麥品種中檢測分子標記Xgwm155,發現其與穗發芽抗性相關,但在67份紅粒春小麥中檢測分子標記Xgwm155卻與穗發芽抗性不相關。張兆萍等[20]在41份黃淮南片區試品系和309份國內外品種中檢測分子標記Xgwm155,發現其與穗發芽抗性不相關。說明與基因和QTL位點相關的分子標記大多具有品種特異性,只在特定的環境或遺傳背景下才有效,且多數分子標記并未實際應用到育種進程中。因此,若想將QTL定位結果和分子標記與實際育種相結合,那么對分子標記在某一特定環境及遺傳群體中的有效性驗證具有非常重要的意義。
3.3 分蘗與穗數相關分子標記的實用性
目前,小麥分子標記大多集中在遺傳作圖和QTL定位等基礎環節,有關分蘗、穗數相關分子標記的有效性研究尚未見報道。經本研究相關分析表明,分子標記Xgwm264擴增出的B、C、D型條帶,分子標記Xwmc169擴增出的A、B型條帶,分子標記Xwmc215擴增出的A、B型條帶,分子標記Xgwm437擴增出的A、E型條帶以及分子標記Xcfd23擴增出的A、C型條帶,均與單位面積最大分蘗數存在極顯著相關性,其中,分子標記Xgwm264和Xwmc169所擴增出的條帶,同時與單位面積穗數存在極顯著相關性,且相關性非常一致。例如,分子標記Xwmc169擴增出的A型條帶,同時與單位面積最大分蘗數和單位面積穗數呈極顯著正相關,擴增出的B型條帶則同時與其呈極顯著負相關。說明這兩個分子標記可能與多效性基因連鎖,可同時用于篩選分蘗數和穗數,具有較高的應用價值。其中,分子標記Xgwm264的等位基因與單位面積最大分蘗數和單位面積穗數的相關系數更高,用于西南麥區育種的準確性更高。分子標記Xwmc215、Xgwm437和Xcfd23擴增出的條帶僅可用于分蘗數的篩選,具有一定的應用價值。分子標記Xbarc232的等位基因與單位面積最大分蘗數和單位面積穗數相關性不顯著,這與湯穎子[28]的研究結果不一致,其研究表明分子標記Xbarc232與單株最高分蘗數和成穗數呈顯著正相關,相關系數分別為0.402和0.299。究其原因可能是研究方法的不同,本研究著重剖析每一種等位基因類型與單位面積最大分蘗數和單位面積穗數的相關性,再通過其相關性確定該分子標記的實用性;同時,由于數量性狀明顯受到環境的影響,因田間數據來源于不同年份,導致不一致的結果。因此,分子標記Xbarc232是否適用于分蘗、成穗數的篩選有待進一步驗證。
4 結 論
分子標記Xgwm264和Xwmc169與由多穗型小麥品種川農18和1BL/1RS易位新品系T1208構建的RIL群體的單位面積最大分蘗數和單位面積穗數具有極顯著相關性,將其用于分蘗成穗數的篩選,可得到具有較高分蘗成穗力的小麥品種(系),為西南麥區高產育種提供理論依據和參考信息。
[1]王紹中,趙 虹,王西成,等.小麥超高產品種篩選的品種初報[J].作物學報,1998,24(6):871.
WANG S Z,ZHAO H,WANG X C,etal.The preliminary research report on screening of new wheat varieties with super-high-yield [J].ActaAgronomicaSinica,1998,24(6):871.
[2]WANG Z Q,LIU Y X,SHI H R,etal.Identification and validation of novel low tiller number QTL in common wheat [J].TheoreticalandAppliedGenetics,2016,129:607.
[3]任正隆.雨養農業區的小麥育種[M].北京:科學出版社,2011:534.
REN Z L.Rain-Fed agricultural region of wheat breeding [M].Beijing:Science Press,2011:534.
[4]任正隆.中國南方小麥優質高效生產的若干問題[J].四川農業大學學報,2002,20(3):302.
REN Z L.Several limiting factors of wheat production in south area of China and the new approach of wheat breeding [J].JournalofSichuanAgriculturalUniversity,2002,20(3):302.
[5]郝艷玲,張紫晉,粟永英,等.西南麥區高產多穗型小麥單株分蘗特征研究[J].核農學報,2016,30(11):2255.
HAO Y L,ZHANG Z J,SU Y Y,etal.Morphological characteristics of tillers per plant in high-yield and multi-spike type wheat in southwest China [J].JournalofNuclearAgriculturalSciences,2016,30(11):2255.
[6]SPIELMEYER W,RICHARDS R A.Comparative mapping of wheat chromosome 1AS which contains the tiller inhibition gene(tin) with rice chromosome 5S [J].TheoreticalandAppliedGenetics,2004,109:1306.
[7]PENG Z S,YEN C,YANG J L.Genetic control of oligo-culms character in common wheat [J].WheatInformationService,1998,86:22.
[8]KURAPARTHY V,SHIPA S,DHALIWAL H S.Identification and mapping of a tiller inhibition gene( tin3) in wheat [J].TheoreticalandAppliedGenetics,2007,114:289.
[9]KURAPARTHY V,SOOD S,GILL B S.Genomic targeting and mapping of tiller inhibition gene( tin3) of wheat using ESTs and synteny with rice [J].Functional&IntegrativeGenomics,2008,8:36.
[10]XU T,BIAN N F,WEN M X,etal.Characterization of a common wheat(TriticumaestivumL.) high-tillering dwarf mutant [J].TheoreticalandAppliedGenetics,2017,130:489.
[11]SHAH M M,GILL K S,BAENZIGER P S,etal.Molecular mapping of loci for agronomic traits on Chromosome 3A of bread wheat [J].CropScience,1999,39:1730.
[12]KATO K,MIURA H,SAWADA S.Mapping QTLs controlling grain yield and its components on chromosome 5A of wheat [J].TheoreticalandAppliedGenetics,2000,101:1117.
[13]LI W L,NELSON J C,CHU C Y,etal.Chromosomal locations and genetic relationships of tiller and spike characters in wheat [J].Euphytica,2002,125:360.
[14]HUANG X Q,CSTER H,GANAL M W,etal.Advanced back-cross QTL analysis for the identification of quantitative trait loci alleles from wild relatives of wheat(TriticumaestivumL.) [J].TheoreticalandAppliedGenetics,2003,106:1386.
[15]LI Z K,PENG T,XIE Q G,etal.Mapping of QTL for tiller number at different stages of growth in wheat using double haploid and immortalized F2populations [J].JournalofGenetics,2010,89:411.
[16]NARASIMHAMOORTHY B,GILL B S,FRITZ A K,etal.Advanced backcross QTL analysis of a hard winter wheat×synthetic wheat population [J].TheoreticalandAppliedGenetics,2006,112:793.
[17]HUANG X Q,KEMPF H,GANAL M W,etal.Advanced back-cross QTL analysis in progenies derived from a cross between a German elite winter wheat variety and a synthetic wheat(TriticumaestivumL.) [J].TheoreticalandAppliedGenetics,2004,109:940.
[18]楊 林,邵 慧,吳青霞,等.小麥分蘗數和單株穗數QTL定位及上位性分析[J].麥類作物學報,2013,33(5):879.
YANG L,SHAO H,WU Q X,etal.QTLs mapping and epistasis analysis for the number of tillers and spike number per plant in wheat [J].JournalofTriticeaeCrops,2013,33(5):879.
[19]馬 麗,李 治,任天恒,等.普通小麥穗發芽抗性相關分子標記在RIL群體中的驗證與評價[J].麥類作物學報,2014,34(4):440.
MA L,LI Z,REN T H,etal.Evaluation and validation of molecular marker associated with pre-harvest sprouting tolerance in a RIL population [J].JournalofTriticeaeCrops,2014,34(4):440.
[20]張兆萍,周麗敏,宋曉朋,等.小麥穗發芽抗性鑒定及相關分子標記的有效性驗證[J].麥類作物學報,2015,35(3):305.
ZHANG Z P,ZHOU L M,SONG X P,etal.Identification of PHS tolerance in wheat varieties and validation of molecular markers associated with PHS tolerance [J].JournalofTriticeaeCrops,2015,35(3):305.
[21]王金萍,劉永偉,孫果忠,等.抗莖腐病分子標記在159份玉米自交系中的驗證及實用性評價[J].植物遺傳資源學報,2017,18(4):760.
WANG J P,LIU Y W,SUN G Z,etal.Evaluation and validation of molecular markers associated with stalk rot resistance in 159 maize inbred lines [J].JournalofPlantGeneticResources,2017,18(4):760.
[22]ZADOKS J C,CHANG T T,KONZAK C F.A decimal code for the growth stages of cereals [J].WeedResearch,1974,14:417.
[23]MURRAY M G,THOMPSON W F.Rapid isolation of high molecular weight plant DNA [J].NucleicAcidsResearch,1980,8(19):4321.
[24]CRESTE S,NETO A T,FIGUEIRA A.Detection of single sequence repeat polymorphisms in denaturing polyacry-lamide sequencing gels by silver staining [J].PlantMolecularBiologyReporter,2001,19:299.
[25]張海萍,常 成,游光霞,等.中國小麥微核心種質及地方品種籽粒休眠特性的分子標記鑒定[J].作物學報,2010,36(10):1651.
ZHANG H P,CHANG C,YOU G X,etal.Identification of molecular markers associated with seed dormancy in mini core collections of Chinese wheat and landraces [J].ActaAgronomicaSinica,2010,36(10):1651.
[26]DENG S M,WU X R,WU Y Y,etal.Characterization and precise mapping of a QTL increasing spike number with pleiotropic effects in wheat [J].TheoreticalandAppliedGenetics,2011,122:287.
[27]HODGKIN J.Seven types of pleiotropy [J].InternationalJournalofDevelopmentalBiology,1998,42:505.
[28]湯穎子.協調型小麥分蘗成穗規律及控制基因的QTL定位[D].雅安:四川農業大學,2016:27.
TANG Y Z.Rules of ear bearing and QTL mapping for spike numbers per plant in coordination-type wheat [D].Ya’an:Sichuan Agricultural University,2016:27.
[29]楊 燕,趙獻林,張 勇,等.四個小麥抗穗發芽分子抗性標記有效性的驗證與評價[J].作物學報,2008,34(1):23.
YANG Y,ZHAO X L,ZHANG Y,etal.Evaluation and validation of four molecular markers associated with pre-harvest sprouting tolerance in Chinese wheats [J].ActaAgronomicaSinica,2008,34(1):23.
[30]楊 燕,張春利,陳新民,等.紅粒春小麥穗發芽抗性鑒定及相關分子標記的有效性驗證[J].麥類作物學報,2011,31(1):59.
YANG Y,ZHANG C L,CHEN X M,etal.Identification and validation of molecular markers for PHS tolerance in red-grained spring wheat [J].JournalofTriticeaeCrops,2011,31(1):59.
