高寒草甸土壤貯水量對封育措施的響應
2018-04-11 07:12:25賀慧丹祝景彬未亞西李紅琴楊永勝李英年
水土保持研究 2018年3期
賀慧丹, 祝景彬, 未亞西, 李紅琴, 楊永勝, 李英年
(1.中國科學院 西北高原生物研究所, 西寧 810001; 2.中國科學院大學, 北京 100049;3.中國科學院 高原生物適應與進化重點實驗室, 西寧 810001)
高寒矮嵩草(Kobresiahumilis)草甸廣布于青藏高原及其周圍山地,約占青藏高原可利用草場的25%[1]。由于高寒草甸植物具有“高蛋白、高脂肪、高熱值、低纖維”三高一低的特點,牧草適口性好,是畜牧業發展的物質基礎。近年來,隨著氣候變暖、人口急劇增長及家畜數量不斷增加,高寒草甸地區草地逐步退化、水土流失加劇,導致該區草地生產力、土壤固碳持水能力不斷下降[2-3]。正因如此,國務院劃分不同生態功能區進行生態恢復與治理工作。2005年開始的《青海三江源自然保護區生態保護和建設總體規劃》(生態治理工程)一期工程,經過幾年的實施,三江源生態系統退化趨勢得到初步遏制,重點生態建設工程區生態狀況逐漸好轉,但生態建設任務的長期性、艱巨性凸顯[4]。
三江源生態治理工程中,封育禁牧是有效恢復和重建植被的辦法之一。隨封育時間延長,植被在正向演替的恢復過程中,其固碳持水能力、土壤水貯存量以及物種多樣性均可發生改變[5-7]。然而,對于生態治理工程實施后生態環境的變化,以往的研究多集中在土壤有機碳、多樣性、生物量[8]等方面。也有一些學者,如徐翠等[9]提出退化使土壤孔隙度等物理性質發生改變,從而影響土壤水分涵養能力。周印東等[10]認為自然植被的正向演替對表層土壤有機質含量有明顯的促進作用,表層土壤田間持水量、容重、總孔隙度等與土壤持水性能相關的指標都與有機質含量呈顯著或極顯著相關。但較少涉及土壤貯水量、持水能力及與環境因素之間關系的研究。本研究以三江源瑪沁縣高寒草甸為研究區域,針對土壤貯水量對封育措施響應的問題,選擇封育和自然放牧地為試驗區,在定期測定土壤濕度、持水量、容重、孔隙度等指標的基礎上,分析植被恢復過程中封育措施下土壤容重、持水能力、貯水量時空變化特征及環境影響機制,以期為生態治理工程實施下高寒草甸生態系統物質循環和能量流動的研究提供科學依據。
1 材料與方法
1.1 研究區概況
本研究在青海省果洛州瑪沁縣進行,瑪沁地區位于青海省東南部,是國家級“三江源”重點生態保護區。瑪沁縣年平均氣溫-0.5℃,氣溫低,日溫差大。年平均降水量514 mm,多集中在6—9月份,屬高原大陸性半濕潤氣候,冬半年干燥少雨,夏半年涼爽濕潤。植被為高寒草甸類型,優勢種有矮嵩草、垂穗披堿草(Elymusnutans)、青藏苔草(CarexsiderostictaHance.)、異針茅(StipaalienaKeng)、高原早熟禾(PoaalpigenaLindm.)。伴生種有鵝絨委陵菜(Potentillaanserina)、異葉米口袋(GueldenstaedtiadiversifoliaMaxim.)、麻花艽(GentianastramineaMaxim.)、美麗風毛菊(Saussureasuperba)、黃花棘豆(Oxytropisochrocephala)、細葉亞菊(Ajaniatenuifolia)、矮火絨草(Leontopodiumnanum)、高原毛茛(Ranunculaceae)等[11]。植被群落結構簡單、分異不明顯。土壤為高山草甸土,土壤發育年輕,有機質含量高,粗骨性強。
1.2 研究材料與方法
1.2.1試驗設計試驗區設在瑪沁縣西北5 km處的果洛州氣象局生態監測點100°12′E,34°28′N,海拔3 761 m的高寒草甸植被類型。封育樣地系生態監測點于2003年封育,僅在春季的2—4月進行適度放牧,時長3個月。未封育樣地為當地牧民自然放牧區,每年10月—翌年5月放牧,時長8個月。放牧強度均約1.3只羊單位/hm2。
觀測區設在封育與未封育樣地,在每個樣地設計10 m×20 m的觀測區,以中央點及10 m×20 m的角點附近為觀測樣點,即每個試驗區共5個點(重復)。
1.2.2監測指標及方法土壤濕度及貯水量:供試分析的土壤水分監測于2013年5—9月每月8日,18日,28日在設計的上述5個重復點右側4 m×4 m的范圍內隨機取樣,每次分別用土鉆收集0—10,10—20,20—30,30—40 cm層次土樣裝入鋁盒,及時稱取鮮重,帶回實驗室烘干至恒重后稱干重,并計算其土壤質量含水率(占干土重的百分比)。土壤貯水量是指自然狀況下一定土層厚度的土壤能夠容納的實際含水量,以土層深度的mm水量表示。有[12]:
W=F×H×SD×/10
(1)
式中:W為土壤貯水量(mm);F為土壤容重(g/cm3);H為土壤厚度(cm);SD為實測的土壤質量含水率;10代表把水層定為mm水量的轉換系數。
植被生物量:于2013年8月底在設計的上述5個重復點左側1 m處進行了50 cm×50 cm樣方的生物量等植被群落特征調查。觀測時先收集枯落物裝袋,其次用剪刀齊地面分種剪下綠色植物裝袋,并收集碎屑物,再用內徑8 cm的根鉆分0—10,10—20,20—40 cm層次收集土柱(每個樣方3個重復,并同層次混合),土柱現場搗碎混合后過篩,將過篩后的土壤和帶根系的土壤分別分層裝袋。帶根系的土壤帶回實驗室后分揀石粒、過篩、清洗等過程挑揀出包括多年死根在內的生物量裝袋。在室內將地上生物量(綠體、枯落物)、碎屑、地下根系生物量等一并置入烘箱,在65℃烘溫條件下烘干至恒重稱重。
土壤容重、持水量:上述監測生物量的樣方附近挖掘剖面,用內徑為5 cm,容積為100 cm3的環刀分0—10,10—20,20—40 cm層次收集環刀土壤,取樣后環刀迅速封蓋并稱量鮮重(m1),用透明膠帶密封后帶回實驗室,室內清除膠帶及污垢,揭去上底蓋,僅留帶網眼的下底蓋放入平底容器中,注入水直至高度恰好在環刀上沿,在水中充分浸透12 h后,蓋上底蓋水平取出稱重(m2);之后,把僅帶有網眼底蓋的環刀放在鋪有干沙的平底盤中2 h,蓋上底蓋稱重(m3);接著,把環刀放在平地盤上2晝夜,再稱重(m4);最后在實驗室烘箱烘干24 h(至恒重)后用電子天平(0.001 g)稱重(m5)。計算前后差值并取平均值,測定得到土壤容重(g/cm3)、土壤持水量、毛管孔隙度(%)等。
降水量:采用果洛州氣象局地面氣象觀測資料。
土壤有機質和有機碳密度:過篩后收集的土壤樣品帶回室內自然陰干,利用重鉻酸鉀容量法—外加熱法測定土壤有機質的含量。有機碳密度(SOC,Kg C/m2)采用以下公式[13]:
(2)
式中:Di,Bi,OMi和S分別為土層厚度(cm)、土壤容重(g/cm3)、土壤有機碳含量(%)和對應面積(cm2);i代表土壤的分層數(i=1,2,3)。
1.2.3數據分析采用Excel 2013進行數據的基本處理和作圖,SPSS 19.0軟件的One-Way ANOVA統計分析指標的平均值、標準差及在p=0.05水平上各處理間的差異顯著性。
2 結果與分析
2.1 土壤容重與土壤持水能力
圖1給出了瑪沁高寒草甸封育與未封育措施下土壤容重變化。由圖1可知,封育與未封育土壤容重均隨著深度增加而增加,最大可達1.27~1.39 g/cm3。
封育使不同層次的土壤容重均有所下降,與未封育區相比,0—40 cm整層土壤平均容重降低了11%,其中0—10 cm和10—20 cm差異達顯著性差異(p<0.05),而20—40 cm無顯著差異。
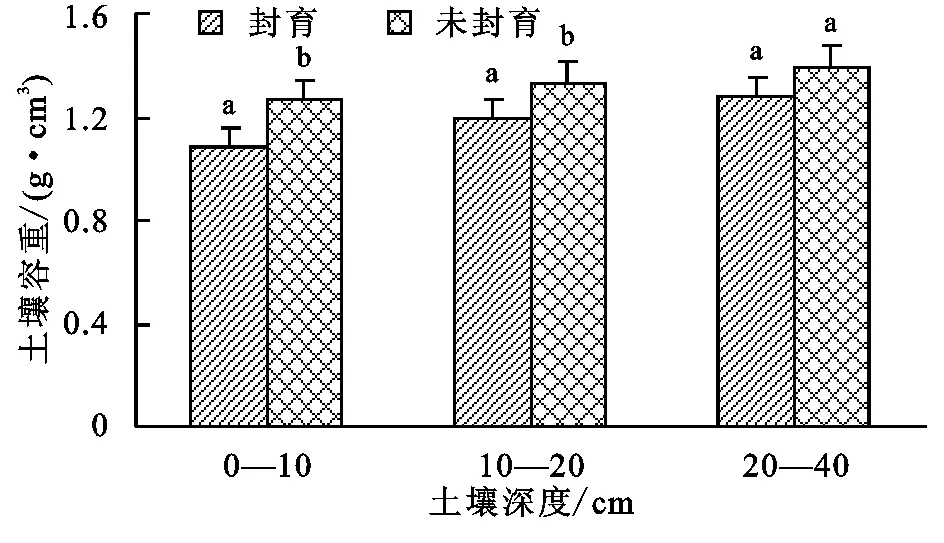
注:圖中字母不同表示兩者達到0.05顯著水平,字母相同表示兩者未達到顯著水平。
圖1封育和未封育土壤容重與土層深度的關系變化
封育11 a后土壤不同層次的最大持水力、毛管持水力、最小持水量均有所增加(表1),0—40 cm整層分別達201.96,177.37,156.61 mm,比未封育區分別高11.34,15.05,15.14 mm,分別增加了6%,9%和11%。封育11 a來,高寒草甸土壤飽和持水量、毛管持水量和田間持水量分別按1.03,1.37,1.38 mm/a的速率增加,封育11 a后毛管持水量和田間持水量增加速率較高。盡管封育樣地與未封育樣地持水能力差異均未達到顯著水平(p>0.05),但封育后3個指標均高于未封育樣地,說明封育措施能提高土壤的持水能力。
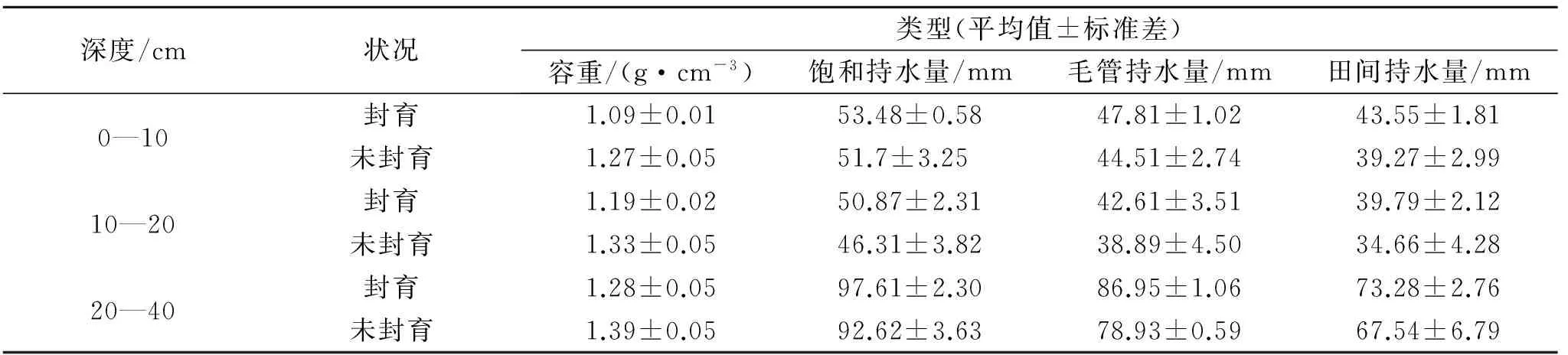
表1 土壤容重及土壤持水量
2.2 植物生長期土壤貯水量季節動態及與封育與非封育環境因素的關系
圖2給出了2013年果洛瑪沁高寒草甸封育與未封育措施下0—40 cm土壤層次貯水量以及降水量在植物生長期的季節動態。圖2看到,封育與未封育措施下0—40 cm土壤層次貯水量動態變化基本一致,封育區0—40 cm層次土壤貯水量波動程度大于未封育樣地且量值較高。兩者貯水量差距在5月底6月初、8月底以后較為明顯。在植物生長期的5—9月,封育與未封育措施下5月初到7月中旬土壤貯水量保持較高的水平,7月下旬以后,土壤貯水量下降明顯,進入9月略有升高。土壤貯水量的這種變化不僅與降雨具有極大的相關性,而且與植物生長對土壤水的利用消耗有關。
從季節動態來看,5月到6月初,受低溫環境影響,植物生長緩慢,葉面積較小,蒸騰作用不強,土壤蒸發也受到溫度影響較弱,底層土壤(一般在40 cm層次以下)仍維持凍結狀態,凍結層在融化過程中不斷產生融凍水來補給土壤上層,同時凍結層的依然存在阻止了水分下滲,加之自然降水產生,促使了土壤貯水量增加。6月中旬到7月下旬,降水明顯增多,但植被生長達到一定高度,葉面積處在增加階段,有降水補給但也有較高的蒸散,二者共同作用下土壤貯水量也較高。其中,7月底封育區降水量具有最高值,但屬于雨熱同期,蒸發強烈,溫度的升高和降雨的增加,植被快速生長,植被生長旺盛消耗大,這也是這一時期降水量大,但貯水量最高值卻小于5月底的原因之一。也說明水分蒸散是高寒草甸地區降水損失的重要因素,蒸散降低相應的土壤貯水量就提高。2013年8月與多年平均降水量相比是偏少較多的一年,整個8月降水量僅為46.8 mm,是多年平均降水量的9.1%,降水偏少明顯,加之植被蒸散在相對較高的溫度環境下蒸散加大,導致8月土壤水散失嚴重,貯水量下降明顯。進入9月后,隨溫度下降,日最低氣溫常處在0℃以下,植被生長減緩,葉面積處在下降階段,植物進入枯黃期,地表面形成一定的枯落物,植物對其下面的地表產生最大的郁閉度,植被蒸騰受植物基本停止生長影響下降明顯,土壤蒸發受較高的郁閉度影響而降低,土壤水分散失減少,加之溫度低,土壤表面日消夜凍,使土壤水分容易積聚,緩慢提高了土壤貯水量。形成了土壤貯水量5月到6月初的融凍集聚期、6月中旬至9月初的波動消耗期和9月以后的蓄積期。
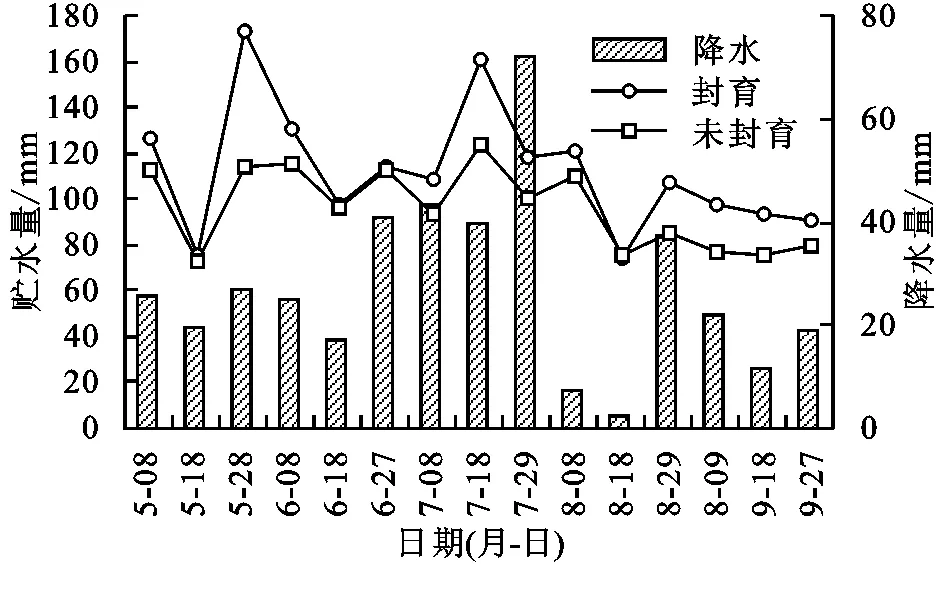
圖2封育和未封育樣地土壤貯水量及時段降水量的季節動態變化
統計發現,5—9月封育與未封育措施下0—40 cm層次土壤平均貯水量在73.38~173.23 mm范圍波動,平均分別為112.58,96.26 mm。封育11 a后,土壤貯水量比未封育試驗區高16.32 mm,平均按1.25 mm/a的速率增加。所表現的貯水量變化說明封育措施能有效提高土壤貯水量,利于土壤水源涵養。
分析生長期5—9月0—40 cm層次土壤平均貯水量與環境要素關系發現(表2,圖3),5—9月平均土壤貯水量與土壤容重具有極顯著負相關(圖3A),與地下生物量、地上生物量和有機碳密度分別呈極顯著正相關(圖3B)、顯著正相關(圖3C)和無相關關系(圖3D)。與土壤容重呈極顯著負相關,說明隨著土壤容重增加,土壤結實,孔隙度降低,大部分水分在地表被蒸發,土壤水分滲水率降低,土壤貯水率下降[14]。與地下生物量呈極顯著正相關反映了植被雖然因植被生長好的狀況下蒸騰提高,但較高的根系量是水分貯存的重要物質條件[15],同時,高寒草甸區的植被大多根系位于0—20 cm,尤其根系死后被分解留有的空隙,增加了土壤顆粒之間的孔隙度,水分下滲較多,貯水量高。地上生物量大,雖然對地面的覆蓋可以減少土壤蒸發,但植被本身蒸騰作用較強,耗水明顯,進而降低了土壤貯水量,由于地上生物量高低與植被覆蓋度關系較大,綠體植物覆蓋度增加可減少土壤水的蒸發,也可減少降水對土壤水的補給,但可增加植被的蒸騰,故實際上土壤貯水量與地上生物量的關系顯得復雜了[16]。草地生態系統中有機碳主要儲存于土壤中,其中土壤有機碳密度的大小取決于土壤容重、土壤有機質含量及土層厚度,容重增加,有機碳密度降低。封育樣地未被啃食,枯落物較多,地面蒸發降低,而分解能使土壤有機質增加,提高有機碳密度,改善土壤結構,有利于土壤持水,貯水量增加。
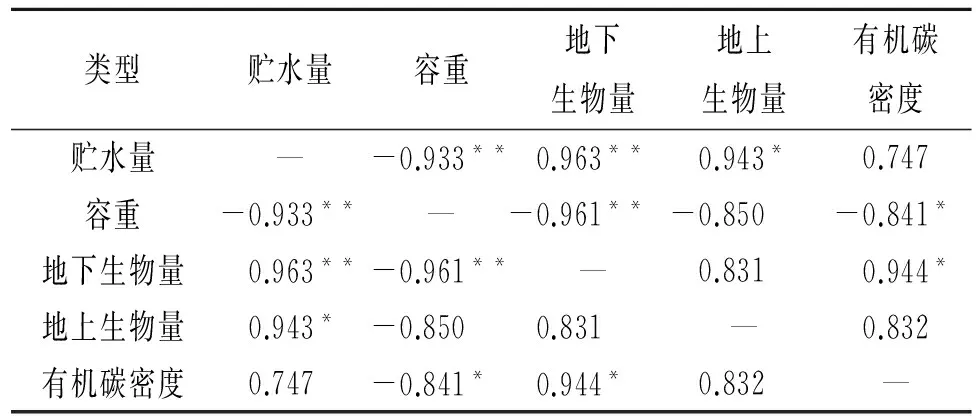
表2 土壤貯水量與容重、地下生物量、地上生物量以及有機碳密度的相關性檢驗
注:*表示顯著相關性檢驗p<0.05;**表示極顯著相關性檢驗p<0.01。
2.3 土壤貯水量的垂直分布
圖4給出了封育與自然放牧的未封育樣地0—40 cm層次土壤貯水量的垂直變化。可以看出,封育和未封育土壤貯水量在植物返青期(5月18日)和地上生物量達最大時期(8月18日)的上層0—10 cm和深層30—40 cm相反,而在強度生長期的7月18日和植物枯黃初期(9月18日),0—40 cm均表現為封育大于未封育。
圖4A看到,在植物生長初期的5月18日,封育和未封育措施下,土壤20—30 cm深度土層貯水量均高于上層和下層,這與上層地面裸露,土壤已融化,底層仍有強度不大的土壤凍結有關。植物生長初期地表蒸發和較小的植被蒸騰導致上層土壤水散失相對較高而導致土壤貯水量低,而下層土壤仍維持一定的凍結層,其土壤水由凍結水和非凍結水組成,極大多非凍結水已通過土壤層次的溫度梯度的作用補充到上一層次,在提高上層土壤貯水量的同時降低了下層土壤貯水量。
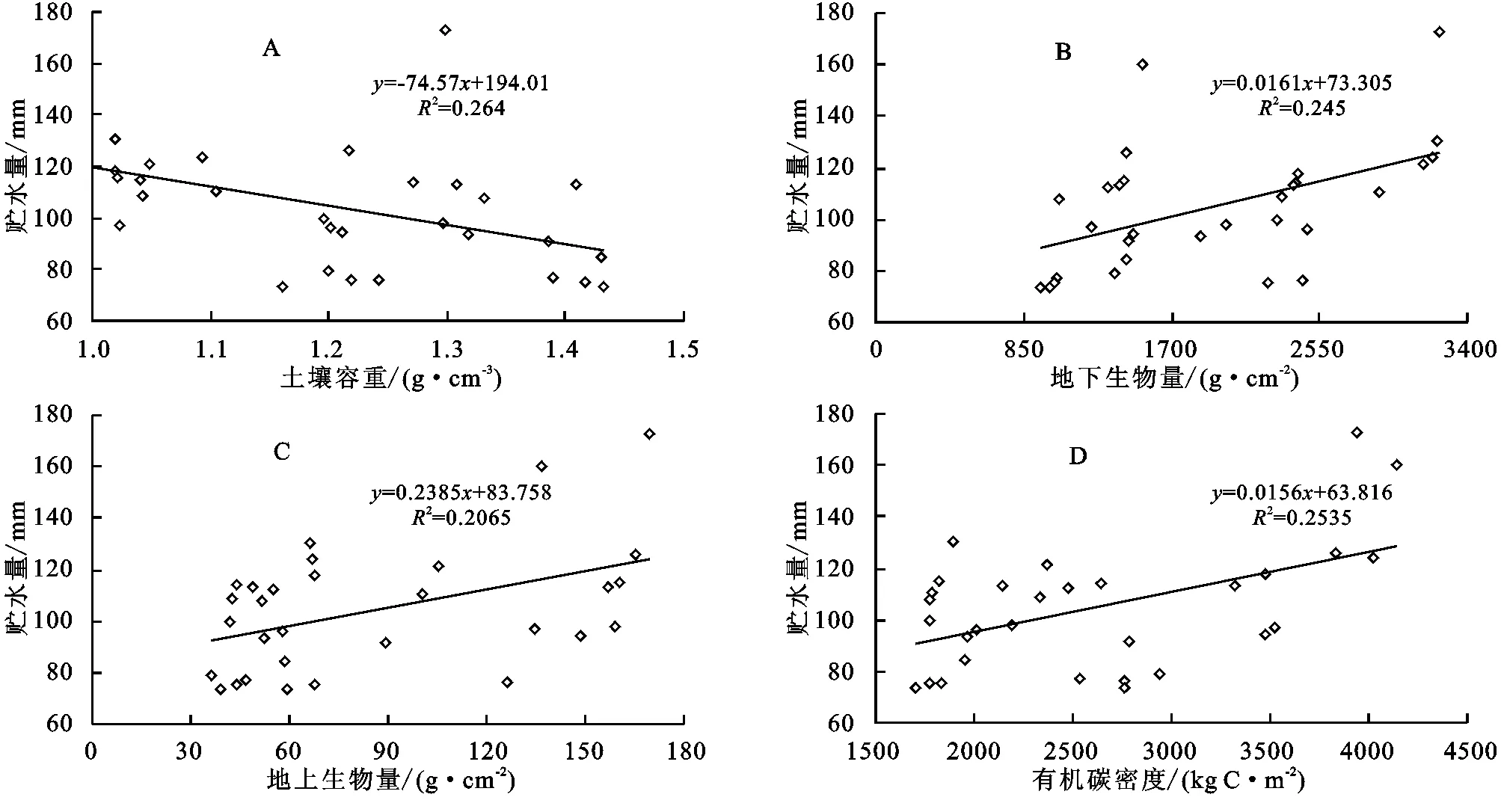
圖3 封育與未封育樣地5-9月平均土壤貯水量與土壤容重、地下生物量、地上生物量以及有機碳密度的關系
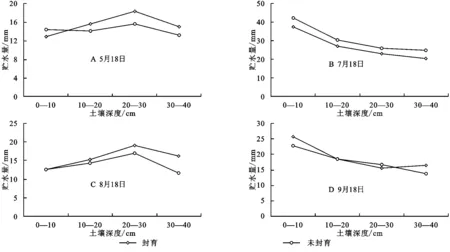
圖4 封育與未封育樣地土壤貯水量的垂直變化
植物進入強度生長期的7月18日(圖4B),土壤貯水量在垂直方向上表現為隨土層深度增加而降低。封育與未封育試驗區垂直變化一致,在任何層次均表現為封育區明顯高于未封育試驗區,且因前期較高降水的影響,垂直變化上均表現出土壤表層高而深層低。7月是年內雨量最為豐沛時期,約達155.5 mm,占植被生長期5—9月降水量409.1 mm的38%,7月9日到7月18日期間,階段降水量仍達40 mm。7月也是年內溫度最高時期,降水量的增加及良好的熱量條件下,植被加快生長,雖然植被蒸騰強,但不及降水的補給大,導致土壤上層土壤貯水量較高。由于植被根系大多分布在較淺,一定程度上阻止降水入滲土壤后的重力水下滲,使大部分水分集中于土壤上層,相對保持了深層土壤貯水量較低。
8月是植被地上生物量達最大時期,8月9日到8月18日(圖4C)期間降水最少,僅為2.3 mm,而降水量的多少是影響土壤水分的主要因素[17]。這一時期溫度較高,太陽輻射強,植被蒸騰和土壤蒸散強烈,豐富的地上生物量對降水有截流作用,使原本很少的降水很難滲入土壤,土壤水分散失量大,導致土壤上層貯水量達年內最低,而深層水由于重力作用得以保存,所以深層土壤貯水量高于上層土壤貯水量。
9月18日(圖4D)植被基本進入枯落期,變化規律與7月18日相似,封育區與未封育區土壤表層貯水量分別為27.2,25.8 mm,相互之間差異很小,但受8月15日前后稍大降水量的影響,土壤水分的滯后效應下0—10 cm土層貯水量有所提高,且封育地大于未封育區。30—40 cm層次土壤貯水量差異逐漸減小。這時期植被生長逐漸停止,植被蒸散作用減弱,地表由于被枯落物和腐殖質覆蓋蒸散作用也較弱,對降水的截流以及攔蓄使水分多集中于地表,這一時期地下生物量較高,能很好的涵養水分,故土壤淺層0—10 cm貯水量高于10—40 cm的深層貯水量。
3 討論與結論
3.1 土壤容重與持水能力
張社奇等[18]的研究表明土壤容重會隨著深度增加而增加,這與本研究的結論一致。土壤容重的大小主要受土壤孔隙度的影響,土壤疏松多孔,則容重越小,毛管吸水力越大,最大持水能力越大。通常來講,土壤的最大持水能力越強,貯水能力越強,對水源的涵養功能越高。植物也通過提高土壤有機質含量改變土壤的結構,使土壤容重降低,并且使毛管孔隙度增大,有利于土壤的持水性[19-20]。本研究中放牧遭受家畜踩踏嚴重,土壤板結,土壤孔隙度減小,容重增加。而土壤持水能力與容重關系密切[21],放牧使土壤容重增加,滲透阻力加大,土壤的保水和持水能力下降。禁牧避免了家畜的影響,使土質變得松軟,表層受到雨水、凍融、根系等作用的影響變得疏松,水分入滲能力加強,土壤顆粒之間疏松,容重降低,同時,封育區植被生長良好,植被高度、蓋度、地上和地下生物量均提高明顯,物種豐富度和多樣性也較高[22],土壤結構改善,也利于土壤容重的下降,水分更易下滲,提高土壤持水能力[23-24]。這表明封育不僅影響土壤容重提高植被生物生產力,同時,對生物固碳和土壤持水能力的提高有利。
3.2 土壤貯水量影響因素
土壤貯水量受土壤容重、土壤有機質、生物量等多種因素的影響,而土壤表層更易受到溫度等的影響。趙中秋等[25]認為不同土地利用類型土壤的自然含水量、持水性能、供水性能和滲透性能均有顯著差異。本研究表明封育區長期禁牧,地面枯落物和腐殖質較多,特別是植物生長初期的5月、6月,以及植物生長末期以后枯落物和腐殖質均較多,對地面覆蓋度大,郁蔽環境和大量的枯草堆積減少了蒸散,進而使土壤貯水量增加。未封育區由于自然放牧,受家畜啃食踐踏影響,土壤孔隙分布的空間格局發生變化,土壤總孔隙減少,滲透阻力加大[26],影響土壤重力水分的下滲,植被生物量和對地面的覆蓋度都較封育低[27],所產生的郁蔽環境較輕,植物的蒸騰和土壤蒸發增強,故封育區土壤貯水量高于未封育區。封育還提高土壤有機質含量,使土壤的團粒結構增加,穩定性團聚體增加,提高了土壤顆粒間的總孔度,利于提高貯水能力。這也與郭濼等[28]關于森林土壤蓄水能力受森林類型枯落物的厚度、孔隙度、有機質含量、溫度、土溫、容重等多方面因素的影響,不同森林類型條件下土壤蓄水能力差異明顯的結果相一致。說明不同利用方式下土壤水分特性與土壤的其他性質如土壤有機質含量、孔隙度、結構團聚體穩定性、容重等有直接或間接的密切關系。土壤的貯水能力在某種程度上還與降水量呈顯著相關,一個地區一定時段內降水量增加,貯水量也增加[29],但是所有變量的增加或減少都不是單一因素作用的結果。貯水量受降水量影響,又與植物生物量、有機物質和土壤質地有關[30]。同樣,不同時期封育與未封育措施下,土壤貯水量隨深度加深其變化規律不盡相同,相互差異各有不同。整體看,高寒草甸的封育措施對土壤上層的影響較深。
3.3 影響土壤持水能力的其他因素
植被截流的因素不可忽略。候瓊等[31]指出,隨著覆蓋度的增加,植被截留量增多,頻繁而量小的降水過程增加了植被截留的幾率,使累積截流量占降水量的比例增大。本文表明,封育樣地長時間禁牧,植被逐漸得到恢復,地上和地下生物量都有提高,地面覆蓋度增加,對降水的截流和攔蓄作用明顯。土壤持水力還與植被根系有關系[32],因為植物的根系直接著生在土壤里,所以土壤水分含量的多少直接影響到植物根系的發育,反過來,植被根系也會影響土壤的持水能力。本文試驗樣地處于高寒地區,晝夜溫差大,氣候干旱,植被類型主要為矮嵩草等具有大量地下根系的高寒草甸,能保護土壤表層減少水力和風力及凍融作用的侵蝕作用,同時,植被根系的存在能增加土壤孔隙度,尤其死亡的根系經過分解,留下的孔隙使土壤變得更松軟,從而使表層容重降低。這也與隨著土層深度增加,土壤容重增加,飽和持水量、毛管持水量和毛管孔隙度降低的結果相一致。
3.4 結 論
(1) 經11 a封育后高寒草甸0—40 cm各土壤層次容重在1.08~1.27 g/cm3,比未封育的自然放牧區平均降低11%,并達顯著性差異(p<0.05),地表0—10 cm層次最為明顯。
(2) 與未封育相比,封育11 a后土壤飽和持水量、毛管持水量和田間持水量均有所提高,0—40 cm整層分別增加了6%,9%和11%,封育11 a來分別按1.03,1.37,1.38 mm/a的速率增加。
(3) 封育與未封育條件下土壤貯水量季節動態變化與降水量基本一致,受季節等不同因素影響可分為融凍集聚期、波動消耗期和蓄積期。貯水量在5月底和7月中下旬達到最大值,其中封育和未封育土壤平均貯水量分別為112.58 mm和96.26 mm,表現為封育>未封育。說明封育能促使土壤貯水量增加,利于土壤涵養水分,放牧使土壤貯水量減少。
(4) 土壤貯水量與土壤容重呈極顯著負相關,分別與地下生物量、地下生物量呈極顯著、顯著的正相關,與有機碳密度無相關。
(5) 土壤貯水量在垂直方向上隨著土層深度增加呈下降趨勢,受降水量影響較大。7月土壤貯水量達到最大值,尤以0—10 cm最為顯著,遠大于30—40 cm土壤層次。說明不同樣地類型土壤貯水量不僅在時間尺度上動態變化,在垂直方向上也存在變化,且在表層和深層封育大于未封育地區。
參考文獻:
[1]王文穎,王啟基,王剛.高寒草甸土地退化及其恢復重建對土壤碳氮含量的影響[J].生態環境,2006,15(2):362-366.
[2]李媛媛,董世魁,李小艷,等.圍欄封育對黃河源區退化高寒草地植被組成及生物量的影響[J].草地學報,2012,20(2):275-279.
[3]劉紀遠,徐新良,邵全琴.近30年來青海三江源地區草地退化的時空特征[J].地理學報,2008,63(4):364-376.
[4]邵全琴,樊江文.三江源區生態系統綜合監測與評估[M].北京:科學出版社,2012:447-563.
[5]何念鵬,韓興國,于貴瑞.長期封育對不同類型草地碳貯量及其固持速率的影響[J].生態學報,2011,31(15):4270-4276.
[6]Wu G L, Liu Z H, Zhang L, et al. Long-term fencing improved soil properties and soil organic carbon storage in an alpine swamp meadow of western China[J]. Plant & Soil, 2010,332(1/2):331-337.
[7]Bilotta G S, Brazier R E, Haygarth P M. The impacts of grazing animals on the quality of soils, vegetation, and surface waters in intensively managed grasslands[J]. Advances in Agronomy, 2007,94:237-280.
[8]范月君,侯向陽,石紅宵,等.封育與放牧對三江源區高寒草甸植物和土壤碳儲量的影響[J].草原與草坪,2012,32(5):41-46.
[9]徐翠,張林波,杜加強,等.三江源區高寒草甸退化對土壤水源涵養功能的影響[J].生態學報,2013,33(8):2388-2399.
[10]周印東,吳金水,趙世偉,等.子午嶺植被演替過程中土壤剖面有機質與持水性能變化[J].西北植物學報,2003,23(6):895-900.
[11]周華坤,趙新全,趙亮,等.高山草甸垂穗披堿草人工草地群落特征及穩定性研究[J].中國草地學報,2007,29(2):13-25.
[12]中國土壤學會農業化學專業委員會.土壤農業化學常規分析方法[M].北京:科學出版社,1984.
[13]劉曉琴,吳啟華,李紅琴,等.不同封育年限高寒草甸植被/土壤碳密度及凈生態系統CO2交換量的比較[J].冰川凍土,2013,35(4):848-856.
[14]李紅琴,喬小龍,張鎰鋰,等.封育對黃河源頭瑪多高寒草原水源涵養的影響[J].水土保持學報,2015,29(1):195-200.
[15]吳啟華,毛紹娟,劉曉琴,等.牧壓梯度下高寒雜草類草甸土壤持水能力及影響因素分析[J].冰川凍土,2014,36(3):590-598.
[16]王潔,賈文雄,趙珍,等.祁連山北坡草甸草原地上生物量與土壤理化性質的關系[J].水土保持研究,2017,24(1):36-43.
[17]劉安花,李英年,張法偉,等.高寒矮嵩草草甸植物生長季土壤水分動態變化規律[J].干旱區資源與環境,2008,22(10):125-130.
[18]張社奇,王國棟,時新玲,等.黃土高原油松人工林地土壤水分物理性質研究[J].干旱地區農業研究,2005,23(1):60-64.
[19]Wu G L, Li Z H, Zhang L, et al. Effects of artificial grassland establishment on soil nutrients and carbon properties in a black-soil-type degraded grassland[J]. Plant and Soil, 2010,333(1/2):469-479.
[20]徐翠,張林波,杜加強,等.三江源高寒草甸退化對土壤水源涵養功能的影響[J].生態學報,2013,33(8):2388-2399.
[21]Su X K, Wu Y, Dong S K, et al. Effects of grassland degradation and re-vegetation on carbon and nitrogen storage in the soils of the Headwater Area Nature Reserve on the Qinghai-Tibetan Plateau, China[J]. Journal of Mountain Science, 2015,12(3):582-591.
[22]孫宗玖,安沙舟,馬金昌.圍欄封育對草原植被及多樣性的影響[J].干旱區研究,2007,24(5):669-674.
[23]Eldridge D J, Wang L, Ruiz-Colmenero M. Shrub encroachment alters the spatial patterns of infiltration[J]. Ecohydrology, 2015,8(1):83-93.
[24]李建興,何炳輝,梅雪梅,等.紫色土區坡耕地不同種植模式對土壤滲透性的影響[J].應用生態學報,2013,24(3):725-731.
[25]趙中秋,蔡運龍,付梅臣,等.典型喀斯特地區土壤退化機理探討:不同土地利用類型土壤水分性能比較[J].生態環境,2008,17(1):393-395.
[26]Leung A K, Garg A, Coo J L, et al. Effects of the roots of Cynodon dactylon and Schefflera heptaphylla on water infiltration rate and soil hydraulic conductivity[J]. Hydrological Processes, 2015,29(15):3342-3354.
[27]何晴波,趙凌平,白欣,等.封育和放牧對典型草原地上植被的影響[J].水土保持研究,2017,24(4):247-251.
[28]郭濼,夏北成,倪國祥.不同森林類型的土壤持水能力及其環境效應研究[J].中山大學學報,2005,44(4):327-330.
[29]馬宗泰.三江源北部天然草地土壤水分動態變化規律[J].安徽農業科學,2009,37(8):3619-3620.
[30]白曉,張蘭慧,王一博,等.祁連山區不同土地覆被類型下土壤水分變異特征[J].水土保持研究,2017,24(2):17-25.
[31]候瓊,王英舜,楊澤龍,等.基于水分平衡原理的內蒙古典型草原土壤水動態模型研究[J].干旱地區農業研究,2011,29(5):197-203.
[32]Schenk H. Vertical vegetation structure below: Scaling from root to globe[J]. Progress in Botany, 2005,66(2):341-373.
