不同放牧強度對錫林郭勒典型草原生產力及碳儲量的影響
2018-05-05 08:53:35杜寶紅高翠萍哈達朝魯
水土保持研究 2018年1期
關鍵詞:植物
杜寶紅, 高翠萍, 哈達朝魯
(1.錫林郭勒職業學院, 內蒙古 錫林郭勒盟 026000; 2 內蒙古農業大學, 呼和浩特 010011)
草原作為世界上分布最廣的重要的陸地植被類型之一,在地球表面占據著一定的自然地帶,分布面積廣泛[1-2]。全球草原面積約5.0×109hm2,約占全球陸地總面積的40%,中國的草原面積為3.55億hm2,約占國土面積的41.7%,是世界草原總面積的6%~8%,居世界第二,主要類型為溫帶草原、高寒草原和荒漠草原等[3-4]。內蒙古草原不僅是國家重要的畜牧業生產基地,同時也是重要的綠色生態屏障,在減少沙塵暴和惡劣天氣的發生方面發揮著作用,也是研究生態系統對人類干擾和全球氣候變化響應機制的典型區域之一和國際地圈—生物圈計劃(IGBP)陸地樣帶—中國東北陸地生態系統樣帶(NECT)的重要組成部分[5-7],但過度放牧和不合理利用,草原發生了不同程度的退化,甚至發生了大面積的沙化、鹽堿化,探討最大限度地優化草原具有重要的意義。
在全球氣候變化成為國際上備受矚目的問題的同時,我國科學家也開始研究管理模式對土壤碳庫和碳循環的影響[8-9]。草原生態系統是陸地碳循環及碳固持的一個重要組成部分,隨著全球變暖等環境問題日益突出,生態系統的碳循環成為人們研究的熱點[10-12]。我國草原生態系統是陸地生態系統的主體類型,貯存的碳總量約為266.3 Pg,占陸地生態系統的12.7%,其中90%貯存在土壤中。受氣候變化及長期不合理的人為活動干擾,我國90%的可利用草原處于不同程度的退化狀態,其中20%以上是由于過度放牧造成的,碳正常循環受到影響[10-12]。管理農事對碳循環及其生態過程的效應研究受到了土壤,環境,全球變化的影響,其中放牧導致的土地退化會使草原生態系統功能發生衰退,影響植被生產力及土壤有機質的分解和積累速率,進而影響土壤碳儲量和生態系統的碳循環,而適度利用可減少草原碳排放,增加土壤碳貯存,故針對不同草原生態系統類型,科學安排利用方式和強度對草原可持續利用尤為重要,這已成為草原管理研究領域的重要命題之一[13-14]。因此,開展放牧草原植被—土壤碳儲量的研究,不僅可以揭示放牧強度對草原植被—土壤碳儲量變化的影響,而且可以闡明放牧草原碳儲量各組分間的比例及其轉化關系,進而從維護碳匯的角度制定可持續利用的放牧制度[15]。典型草原是內蒙古草原的重要組成部分,屬于草原區向荒漠區過渡的旱生化草原生態系統,群落結構較單一化,種群密度和群落結構容易發生改變[5-6]。已有研究表明,在不同的放牧強度下或連續過度放牧下,典型草原草原植物種類組成改變,地表植被的生物量、生物多樣性和豐富度下降土壤容重增加,孔隙度減小,土壤質量退化[13-14]。土地與植被的退化必然會引起草原有機碳含量的變化,那么,典型草原不同放牧強度下土壤、植物的有機碳含量是否存在顯著差異以及如何分配,二者的相互關系如何?加強這些問題的研究有助于揭示放牧對典型草原碳匯的影響機制。有鑒于此,本研究以羊草典型草原為研究對象,研究不同放牧強度下植被—土壤系統有機碳儲量,旨在對比不同放牧強度典型草原有機碳含量與儲量的變化及其差異性,為科學制定草原保護、利用策略和提高草原生態生產功能的適應性供科學依據。
1 材料與方法
1.1 研究區概況
本試驗依托內蒙古錫林郭勒草原生態系統國家野外科學觀測研究站(內蒙古站,43°38′N,116°42′E,海拔1 224 m)進行。位于內蒙古錫林郭勒盟錫林河流域中段,即中國科學院草原生態系統定位站1979年圍封針茅草原附近,該站位于內蒙古錫林河流域,氣候屬溫帶半干旱大陸性季風氣候,冬季寒冷干燥,夏季受季風影響,較為溫暖和濕潤。1970—2010年平均氣溫為0.4℃,其中最冷月(1月)平均氣溫為-21.4℃,最熱月(7月)平均氣溫為19.0℃。年降水量為337 mm,無霜期約為100 d,6-8月降水量占全年降水量的70%,降水量年際變異較大。地帶性土壤類型為暗栗鈣土,土層厚度可達1 m以上,地帶性植被為大針茅草原。試驗樣地位于羊草(Leymuschinensis)樣地北側,2004年通過國際合作項目由中德雙方共同建設,整個試驗區占地面積約300 hm2,群落類型為羊草+大針茅草原。試驗處理前,群落物種分布比較均勻,常見植物有45種,其中以廣旱生根莖禾草羊草占優勢,其次為大針茅、阿爾泰草、冰草、糙隱子草等多年生叢生禾草,這些禾草的地上生物量占群落總生物量的70%以上。常見種和稀有種均以雜類草為主,包括:星毛委陵菜、野韭等[3-5]。
1.2 試驗設計
從2011年8月下旬開始,每年進行放牧處理,試驗用綿羊統一選擇健康的2歲母羊,放牧處理于每年的6月上旬開始,9月中旬結束。采用連續放牧方式,晚上羊群在樣地不歸牧,不進行補飼。在羊草草原選擇4個代表類型,其中對照樣地為1979年圍欄封育樣地(無放牧,NG),另外3個樣地為放牧退化樣地,選取以自由放牧方式利用的草原為研究對象,以牧戶為起點向外輻射狀草原形成由重到輕的一個放牧退化梯度,根據草原植物種類組成、地上生物量、群落高度、群落蓋度等指標,劃分為對照(無放牧,NG)、輕度放牧(Light grazing,LG)、中度放牧(Moderate grazing,MG)和重度放牧(Heavy grazing,HG),不同重復小區內的4個處理完全隨機排列。設定不同的載畜率值分別為0(對照),0.91(輕度放牧),1.82(中度放牧)和2.71(重度放牧)羊單位/hm2/半年[3-5]。
利用隨機樣方法,2016年8月在NG,LG,MG和HG樣地分別設置1 m×1 m樣方10個,記錄和測定植物種類組成、蓋度和高度;篩選其中5個樣方采用齊地面刈割牧草的方法測定地上生物量鮮重,帶回實驗室65℃烘干至恒重;地下生物量與地上生物量在同一樣方內測定,采用剖面法,進行10 cm×10 cm×10 cm土塊切割,每10 cm為一層進行取樣,取到60 cm深度,將取得的樣品帶回實驗室進行篩根、洗根、烘干、稱重。土壤樣品亦在上述樣方處利用土鉆取樣,取樣深度為0—10,10—20,20—30,30—40,40—50,50—60 cm,每樣方內取3次后按層混合,帶回實驗室風干、過篩,測試相關指標,并在每個樣方挖取土壤坡面,環刀法測定土壤容重。取回的根樣用水洗法將其漂出,裝入信封。將根樣與草樣一起烘干(65℃)稱重,得到地上部分、地下部分生物量的值(g/m2)。同時記錄群落地形、地貌特征和土壤質地,GPS定位樣地經緯度和海拔。將根樣與草樣分別粉碎并過80目的篩,然后用元素分析儀測出含碳量(%)。
土壤—植物有機碳測定采用重鉻酸鉀-外加熱法,土壤碳儲量計算公式如下[11,16]:
式中:P為土壤碳儲量(g/m2);di為有機碳含量(%);Hi為土層高度(cm);bi為土壤容重(g/cm3);n為土層深度(cm)。
多樣性指數主要用來反映植物的豐富程度,是植物豐富度和均勻程度的綜合反映,多樣性指數越高,植被的物種多樣性越豐富。均勻度是植物分布均勻程度的反映,在一個植被中,各個種的相對密度越均勻,即各物種的個體數很接近,均勻度值越高。豐富度是植被物種多樣性豐富程度的反映,當個體數量一定時,物種數越多,計算公式如下[17]:
Patrick豐富度指數(S)
Shannon-Wiener多樣性指數(H):
H=-∑PilnPi
Gleason優勢度指數(D):
D=1-∑(Pi)2
Pielou均勻度指數(JP):
JP=-∑PilnPi/lnS
式中:S代表樣方內物種數目;Pi為樣方內種的相對重要值;相對重要值(Pi)=(相對覆蓋度+相對高度+相對多度)/3。此外,為了比較不同類型植被多樣性指數間的關系,方差分析和多重比較用來檢驗它們之間的差異顯著性。
Excel 2007和SPSS 18.0進行數據統計和方差檢驗,以平均值±標準誤差表示(Mean±SE),單因素方差分析(One-way ANOVA)。
2 結果與分析
2.1 放牧強度對物種特征的影響
放牧是天然草原群落受干擾的主要方式之一。在牧壓梯度上,植物群落及其主要植物種群的數量特征發生明顯的變化,主要表現為植物群落密度MG>LG>NG>HG,過牧區最低(表1)。優勢植物羊草的種群密度在中牧區最大,輕牧區次之,重牧區最低,而其相對密度從輕牧到重牧的梯度上逐漸降低。這些變化是由于不同強度的放牧牲畜踐踏,羊草種群大株叢破碎,由一個母株分離出若干個獨立小株叢的結果。羊草種群密度表現為MG>LG>NG>HG。表明適度的放牧有利于羊草根莖的生長發育,但超過一定的閾值后,如在過牧區羊草種群也會迅速消退。大針茅的種群密度MG>LG>NG>HG,表現出一定的耐牧性。禾本科草的種群密度從輕牧區到重牧區逐漸降低。綜上所述,天然草原在不同強度的放牧干擾下,植物生態適應對策是多樣的,這種多樣性是由植物自身生物生態學特性決定的。天然草原群落正是由于多樣的植物生態適應對策,對放牧干擾產生明顯的緩沖作用,表現為一定范圍內的草原群落抗干擾能力和系統穩定性。
2.2 放牧強度對物種多樣性的影響
多樣性指數是植被和生態系統恢復過程最重要的特征之一,是群落生物組成結構的重要指標,不僅能夠反映植被群落組織化水平,而且通過結構與功能的關系可間接反映群落功能的穩定性和發育階段。由圖1可知,隨著放牧強度的增加,典型草原植被覆蓋度、豐富度指數、均勻度指數、多樣性指數、優勢度指數均降低,其中LG,MG和HG均與NG達到顯著差異水平(p<0.05),并且均表現出NG>LG>MG>HG變化趨勢;輕度放牧豐富度指數、均勻度指數和多樣性指數與中度放牧差異不顯著(p>0.05)。HG具有顯著較低的群落均勻度水平,MG樣地的均勻度水平最高,顯著高于3個不同管理方式不同利用程度的羊草群落(p<0.05)(圖1)。這反映了較低水平的利用程度使群落內物種重要值的差異逐漸變大,優勢種的優勢程度被增強,群落的均勻程度降低,而HG降低了物種創造低競爭壓力的生存機會,從而群落內各種的重要值和群落地位差異并未充分顯露和穩定,使得群落的均勻度呈現較高水平。在輕牧區,群落有較低的植物均勻度和多樣性,隨著放牧強度增加,群落中的優勢植物逐漸被一些耐牧、適牧植物所替代,使中牧區的群落具有較高的植物多樣性和均勻度,如果繼續增加放牧強度,群落的均勻度和多樣性又降低,直至群落結構遭到致命的破壞。

表1 放牧強度對物種特征的影響

注:不同小寫字母表示差異顯著(p<0.05)。下同。
圖1放牧強度對物種多樣性的影響
2.3 放牧強度對地上生物量及碳儲量的影響
由于植物光合產物的生產和累積能力,以及牲畜的采食方式和喜食程度的差異,不同放牧強度下植物群落地上生物量的差異較大,導致地上植物有機碳儲量差異顯著(圖2)。由圖可知,隨著放牧強度增加,地上生物量和凋落物量呈遞減趨勢,NG地上植被和凋落物碳儲量最高,其大小依次為NG>LG>MG>HG,放牧顯著降低了群落的地上植物有機碳儲量(p<0.05),除LG與MG之間差異不顯著外,其余放牧梯度間差異顯著(p<0.05)。以NG作對照,LG,MG,HG平均群落地上生物量分別下降15.63%,25.98%和46.12%。地表凋落物也表現出同樣的規律,與群落地上生物量相比下降的更為明顯。放牧干擾降低了植物群落的生產力,直接影響植物對土壤水分和營養元素的吸收,使有機物質生產和地表凋落物累積的減少,歸還土壤的有機質降低,回歸土壤養分和保持水分的能力下降,給土壤的理化性狀造成影響,從而導致土壤貧瘠化和干旱化,進而導致群落結構的簡單化和生態功能的下降。
2.4 放牧強度對根系碳儲量的影響
草原生物量的80%以上集中在地下,地下生物量是土壤有機碳庫的最主要輸入源,在草原生態系統碳循環中起著關鍵作用。分析放牧強度對根系碳儲量的影響,結果表明,典型草原植物地下部分(根系)有機碳儲量明顯高于地上部分有機碳儲量(表2),隨著土層深度的增加,根系有機碳儲量呈現減少的趨勢,均以0—10 cm到10—20 cm下降的量最多。從地下0—60 cm根系有機碳儲量來看,大小序列仍為NG>LG>MG>HG,NG和LG差異不明顯,但顯著高于MG和HG(p<0.05)。0—10 cm根系有機碳儲量占到0—60 cm根系總有機碳儲量的50%以上,0—20 cm根系有機碳儲量占到0—60 cm根系總有機碳儲量的20%以上,因此根系總有機碳儲量在不同放牧強度間的變化主要受0—20 cm根系有機碳儲量的影響,信息量可達70%以上,0—20 cm根系有機碳儲量仍以NG最高,HG最低。

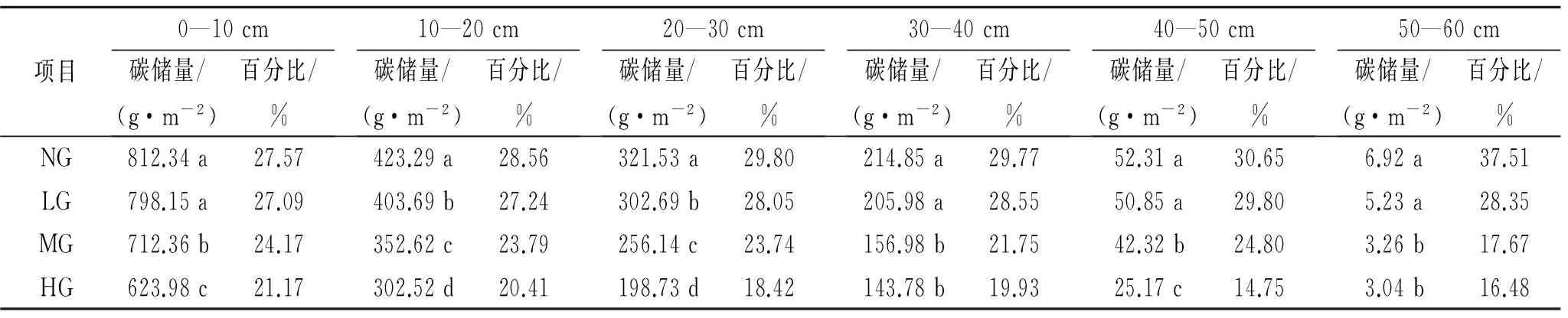
圖2 放牧強度對地上生物量及碳儲量的影響
2.5 放牧強度對土壤有機碳儲量的影響
土壤是草原生態系統中碳的主要存儲庫,占系統的80%以上。放牧利用在明顯影響植被生物量的同時,也會對土壤的碳儲量產生影響,致使草原土壤碳儲量產生差異。本研究比較了不同放牧強度下草原的土壤碳儲量(表3),結果顯示,土壤有機碳含量隨放牧強度的增加呈先增加后減少趨勢,并在MG和LG樣地達到最大。不同放牧強度只對0—40 cm土層有機碳含量影響差異顯著,在40 cm以下土層,其影響不顯著(p>0.05)。土壤有機碳含量均隨土層深度的增加呈降低趨勢,其中40—50 cm,50—60 cm土層有機碳含量及儲量差異不顯著(p>0.05)。由表3還可知,表層0—10 cm土壤有機碳儲量最高,達到2.19~2.89 kg/m2,已遠遠大于植被有機碳儲量,0—10 cm的淺層土壤中,NG和HG土壤碳儲量差異不明顯(p<0.05),MG和LG表現出較強的儲碳能力,10—20 cm呈現出顯著降低的特征;從0—20 cm的土壤總碳儲量來看,該深度范圍內的碳儲量占到0—60 cm碳儲量的30%~40%,碳儲量大小序列為MG>LG>NG>HG,MG和LG的土壤儲碳優勢逐漸表現出來,MG表現出強勁的固碳能力,其碳儲量顯著高于其他,而在這一深度范圍,HG碳儲量亦較低;從0—60 cm土層深度來看,碳儲量表現出隨著退化程度的加重而增加的趨勢,NG土壤碳儲量顯著下降,不同退化程度之間均存在顯著性差異(p<0.05),其中MG的固碳能力最強,其余依次為LG,NG和HG,HG相對于NG土壤有機碳儲量顯著減少,降幅為6.23%。由此可以看出,適度放牧有助于土壤有機碳的積累。
2.6 放牧強度對生態系統有機碳儲量的影響
由圖3可知,不同放牧強度典型草原土壤有機碳儲量大小序列為MG>LG>NG>HG,植被有機碳儲量大小序列為NG>LG>MG>HG,生態系統有機碳儲量大小序列為MG>LG>NG>HG,與土壤有機碳儲量保持一致的變化規律,由此說明放牧對植被—土壤系統有機碳儲量的影響具有非同步性,輕度和中度放牧有助于生態系統有機碳儲量的積累,而重度放牧草原生態系統有機碳儲量顯著降低。隨著放牧強度的增加,地上植物碳儲量占生態系統有機碳儲量比例分別為17.85%,15.68%,13.01%,15.29%,土壤碳儲量所占比例分別為82.15%,84.32%,86.99%,84.71%。土壤碳貯量占總碳貯量的80%以上,是草原碳儲量的主體部分,影響著整個生態系統有機碳儲量的變化。各樣地各部分所占生態系統有機碳儲量比例均表現為土壤>根系>地上植物>凋落物。

表3 放牧強度對根系碳儲量的影響

圖3放牧強度對生態系統有機碳儲量的影響
2.7 土壤碳儲量與植被碳儲量及土層深度的關系
本研究分析了植被地上生物量、地下生物量(0—60 cm)與土壤有機碳儲量的相關性,以及不同放牧強度下,草原土壤不同深度的碳儲量與相應深度的植被地下生物量關系(表4)。植被地上碳儲量與0—20 cm土壤碳儲量存在著顯著的相關性(p<0.05),其相關系數隨著土層深度的增加而降低,以MG的相關系數絕對值最高,而20—40 cm以下,植被地上碳儲量與土壤碳儲量沒有顯著的相關性(p>0.05)。
上述研究表明,不同放牧強度下草原土壤有機碳的垂直分布呈現出了較好的一致性變化,即隨著土層深度的增加土壤有機碳儲量呈遞減趨勢。為了定量描述這種趨勢,本研究利用回歸分析方法,建立土層深度與土層有機碳儲量的回歸方程(表4)。對于不同放牧強度下,線性方程、對數方程與一元二次方程的效果都比較好,且對數方程和二次方程效果略優于線性方程。另外,研究還將4組數據綜合,建立通用方程,發現3類方程的效果也是可以接受的。總之,回歸分析結果表明土壤有機碳儲量與土層深度具有較好的相關關系。

表4 放牧強度對根系碳儲量的影響
注:*表示顯著相關(p<0.05),**表示極顯著相關(p<0.01)。

表5 土壤有機碳儲量與土層深度的回歸分析
注:y為土壤有機碳儲量,x為土層深度。
3 討論與結論
天然草原的生產從大尺度上來講,主要取決于當地的氣候條件、地形特征和土壤特性等環境因子,但對同一塊草原而言,這些因素的影響是恒定不變的,那么放牧是影響草原群落特征最主要的因子。放牧對草原的影響,首先表現在草原植物的變化,地上生物量是反映草原生態系統、放牧系統穩定性的重要指標,其大小可判斷草原狀況、生產潛力和載畜能力等,在放牧脅迫下,群落組分在放牧中有直接的損失,影響植物群落的生產力[18]。本研究表明,隨放牧強度的增加,群落地上生物量呈現出降低的變化趨勢,由此看來,隨著放牧強度的增強,家畜的頻繁啃食降低了牧草的生物量,影響其物質能量的積累,進而影響植物的生長發育和繁殖,造成群落中物種個體普遍減少。另外,家畜的踩踏也影響了草原的土壤特性,這也引起草原植物群落生產力的降低。這與段敏杰[19]、董全民[20]等在天然草原的研究結果一致。不同功能群植物地上生物量和占群落生物量的比例變化主要受放牧過程中放牧家畜選擇性采食的影響。放牧過程中,家畜優先選擇采食優良牧草(禾草和莎草),尤其是禾草,使禾草原上生物量減小。主要是因為禾草相對高度較高,處于草群上層,家畜優先采食,同時抑制了其生長發育。上層高大植物的減少,使草層下部能接受到陽光照射,光合作用增強,促進了物質能量的積累。且受到上層禾草郁蔽環境的影響減小,增強了自身的補償作用和放牧抵抗力,比例增加;雜類草和毒草是適口性差或不能食用的,同時上層高大草類的減少降低了對其的抑制作用,故隨放牧強度的增加,比例增加[21]。
植物多樣性對植物群落穩定性的影響是通過不同功能群間的補償作用實現的,在牧壓梯度下,物種和植物功能群間的這種補償作用盡管有時是十分有限的,但對維持生態系統結構與功能過程的相對穩定性卻具有極其重要的作用[22]。本研究表明,隨著放牧強度的增加,物種豐富度和多樣性指數基本都呈現出單峰變化,即在中度放牧區多樣性最高,雖然均勻度指數的變化趨勢較復雜,但在中度放牧的均勻度指數高于重度放牧。這是因為通過適度的放牧干擾,家畜對植物的采食作用,抑制了優勢種的競爭能力,使弱勢物種的入侵和定居成為可能,一些下繁草的物種數量增加,同時家畜不喜食和不采食的雜類草和毒草類的數量也增加,增加了群落結構的復雜性,群落內物種的多樣性出現一定程度的增加。這在一定程度上支持了“中度干擾假說”[23]。即干擾對資源和環境異質性的作用表現為非線性,適中水平的干擾增加物種多樣性,而超過一定閾值的干擾又會降低物種多樣性。重度放牧下群落物種豐富度、多樣性指數和均勻度指數都表現為最低,當放牧壓力過大時,群落內的可食性牧草因家畜啃食過度而失去再生能力,以致在群落中消失,加之不具可食性的毒雜草在群落中所占的比例增加,而且由于家畜的過度采食,降低了有機質向土壤中的輸入,土壤營養過度消耗,降低了植物的競爭能力,使草原植物群落中的物種多樣性下降。
植被是土壤有機碳的主要輸入來源,在放牧退化演替過程中,植被演替的短期效應較為明顯,而土壤作為一個相對穩定的系統,其變化較為緩慢,尤其是土壤有機碳的積累是長期結果,這就致使土壤有機碳儲量與植被狀況的關系變得復雜而不確定[24]。不同干擾強度主要是通過家畜采食改變地上部分,降低植物地上生物量(圖2);過度放牧,使植物葉面積降低,草原初級生產固定碳素的能力下降,光合作用向地下的能量輸入減弱,導致植物地下根系的生產力和現存量降低;草原生物量降低使凋落物的積累和輸入降低,減少了碳素由植物凋落物向土壤的輸入,降低土壤有機碳含量,使土壤碳儲量降低,而適度放牧有利于草原有機碳的蓄積,因為它可以減少凋落物的積累,促進其分解并加速碳向土壤的輸送[24]。不同放牧強度草原土壤有機碳儲量大小序列為MG>LG>NG>HG,輕度和中度放牧提高了生態系統有機碳水平。導致這一變化的原因可能歸因于不同放牧強度下根冠比、凋落物分解及地上生物量的差異;一方面,放牧強度的加大導致植物地下生物量增加,使得植物根冠比率增加,增加了有機碳向地下的分配量[25];另一方面,動物的踐踏使凋落物破碎并與土壤充分接觸,促進凋落物的分解,有助于碳和養分元素轉移到土壤中而重度放牧促使草原植被葉面積和儲藏物質嚴重減少,凈初級生產力降低,物質的輸入小于輸出,導致生態系統碳儲量降低。Frank等[26]指出放牧會抑制土壤侵蝕的發生,從而保護了土壤有機碳含量。食草動物排泄物的歸還也使土壤表層速效養分增加,土壤的礦化作用加強。此外,放牧使植物的組成發生變化,導致了低的產草量,增加了碳向地下的分配量。放牧還使凋落物積累量減少,這是由于動物的踐踏使凋落物破碎并與土壤充分接觸,有助于凋落物的分解,也有助于碳和養分元素轉移到土壤中。
在生態系統碳儲量中,有機碳儲量從大到小依次為:土壤有機碳儲量、根系有機碳儲量、地上植物有機碳儲量和凋落物有機碳儲量,由此可以看出,不同放牧強度羊草生態系統有機碳儲量中土壤有機碳儲量占居主導地位,這一結果與對典型草原和草甸草原生態系統碳儲量研究所得到的結果相似[3-5]。不同放牧強度草原0—10 cm土壤有機碳儲量均為最高,這一方面是由于地上植物的枯萎、凋落物沉積在土壤表層,促進了表層土壤有機碳的輸入,另一方面歸因于草原地下生物量大部分集中于表層土壤中,隨著深度增加,數量急劇降低[25,27]。隨著放牧強度的增加,地上植物、凋落物、根系有機碳儲量所占生態系統有機碳儲量比例呈現降低趨勢,而土壤有機碳儲量所占比例呈遞增趨勢,0—40 cm土壤碳儲量占到系統總量的80%以上,說明草原碳儲量主要儲存在土壤當中,如李怡[27]指出,針茅草原地上植被碳儲量僅占總碳儲量的2%~5%,其余大部分碳儲存在土壤中,土壤表現出了碳匯的特征,如果大范圍進行科學保護與適當利用,其碳固持能力,也即草原的碳匯功能將得到充分的發揮,其固碳潛力巨大。另外,很多研究對土壤碳垂直分布做出了定性判斷,認為草原生態系統土壤有機碳隨著深度的增加而減少,土壤有機碳的垂直分布格局受凋落物量、淋溶作用、植物根系分布及活動特征和微生物活動等多種因素的影響,在典型草原地區隨著土層深度的增加,土壤溫度、含水量和質地條件變差,有機質來源減少,因此出現了有機碳隨土層深度的增加而表現出逐漸降低的趨勢[23,27]。本研究發現,土壤有機碳儲量與深度之間存在著顯著的相關性,而且這一關系可以用線性、對數和二次函數來定量描述。通過本研究認為,面臨當前草原退化嚴重的現狀,實施圍封禁牧具有重要意義,但是,在退化草原的恢復過程中要加強對圍封不干擾生態系統的監測,同時,要科學認識放牧利用對維持草原生態系統健康的重要性及特殊意義,適時采取相應措施,發揮適度放牧利用對退化草原碳積累和恢復進程的優化調控作用。
致謝:特別感謝中科院內蒙古錫林郭勒草原生態系統國家野外科學觀測研究站所有工作人員和研究人員在本研究中提供的幫助和支持。
參考文獻:
[1] Dick M, Silva M A D, Dewes H. Life cycle assessment of beef cattle production in two typical grassland systems of southern Brazil[J]. Journal of Cleaner Production, 2015,96:426-434.
[2] Zhu X, Yu G, Wang Q, et al. Seasonal dynamics of water use efficiency of typical forest and grassland ecosystems in China[J]. Journal of Forest Research, 2014,19(1):70-76.
[3] Shi Y, Wang Z Q, Zhang X Y, et al. Effects of nitrogen and phosphorus addition on soil microbial community composition in temperate typical grassland in Inner Mongolia[J]. Acta Ecologica Sinica, 2014,34:4943-4949.
[4] Han D, Wu P, Zhang Q, et al. Feature extraction and image recognition of typical grassland forage based on color moment[J]. Transactions of the Chinese Society of Agricultural Engineering, 2016,32(23):168-175.
[5] 柴曦,梁存柱,梁茂偉,等.內蒙古草甸草原與典型草原地下生物量與生產力季節動態及其碳庫潛力[J].生態學報,2014,34(19):5530-5540.
[6] 王晶,王珊珊,喬鮮果,等.氮素添加對內蒙古退化草原生產力的短期影響[J].植物生態學報,2016,40(10):980-990.
[7] 于麗,趙建寧,王慧,等.養分添加對內蒙古貝加爾針茅草原植物多樣性與生產力的影響[J].生態學報,2015,35(24):8165-8173.
[8] Genet H, Mcguire A D, Barrett K, et al. Modeling the effects of fire severity and climate warming on active layer thickness and soil carbon storage of black spruce forests across the landscape in interior Alaska[J]. Environmental Research Letters, 2013,8(4):045016.
[9] Torn M S, Trumbore S E, Chadwick O A, et al. Mineral control of soil organic carbon storage and turnover[J]. Nature, 1997,389(6647):170-173.
[10] Poeplau C, Marstorp H, Thored K, et al. Effect of grassland cutting frequency on soil carbon storage:A case study on public lawns in three Swedish cities[J]. Soil, 2016, 2(2):1-23.
[11] Schwendenmann L, Pendall E. Effects of forest conversion into grassland on soil aggregate structure and carbon storage in Panama: Evidence from soil carbon fractionation and stable isotopes[J]. Plant and Soil, 2006,288(1):217-232.
[12] Reid J P, Adair E C, Hobbie S E, et al. Biodiversity, Nitrogen Deposition, and CO2Affect Grassland Soil Carbon Cycling but not Storage[J]. Ecosystems, 2012,15(4):580-590.
[13] Milchunas D G, Lauenroth W K, Chapman P L, et al. Effects of grazing, topography, and precipitation on the structure of a semiarid grassland[J]. Plant Ecology, 1989,80(1):11-23.
[14] Harrison S, Inouye B D, Safford H D. Ecological Heterogeneity in the Effects of Grazing and Fire on Grassland Diversity[J]. Conservation Biology, 2003,17(3):837-845.
[15] Bullock J M, Hill B C, Dale M P, et al. An experimental study of the effects of sheep grazing on vegetation change in a species-Poor grassland and the role of seedling recruitment into gaps[J]. Journal of Applied Ecology, 1994,31(3):493-507.
[16] Masiello C A, Chadwick O A, Southon J, et al. Weathering controls on mechanisms of carbon storage in grassland soils[J]. Global Biogeochemical Cycles, 2004,18(4):449-460.
[17] Bakker E S, Ritchie M E, Olff H, et al. Herbivore impact on grassland plant diversity depends on habitat productivity and herbivore size. [J]. Ecology Letters, 2006,9(7):780-789.
[18] Gustavsson E, Lennartsson T, Emanuelsson M. Land use more than 200 years ago explains current grassland plant diversity in a Swedish agricultural landscape[J]. Biological Conservation, 2007,138(1/2):47-59.
[19] 段敏杰.放牧干擾下藏北紫花針茅高寒草地生物量遙感監測[D].北京:中國農業科學院,2011.
[20] 董全民,趙新全,馬玉壽,等.放牧強度對高寒混播人工草地群落特征及地上現存量的影響[J].草地學報,2012,20(1):10-16.
[21] An Hui, Li Guoqi. Effects of grazing on plant biomass and soil nutrient in desert steppe[J]. Journal of Plant Nutrition & Fertilizer, 2013,19(3):705-712.
[22] May F, Grimm V, Jeltsch F. Reversed effects of grazing on plant diversity: The role of below-ground competition and size symmetry [J]. Oikos, 2009,118(12):1830-1843.
[23] 劉艷紅,趙惠勛.干擾與物種多樣性維持理論研究進展[J].北京林業大學學報,2000,22(4):101-105.
[24] Gang C, Zhang J, Li J. The Advances in the carbon source/sink researches of typical grassland ecosystem in china[J]. Procedia Environmental Sciences, 2011,10:1646-1653.
[25] Xiong D, Shi P, Sun Y, et al. Effects of grazing exclusion on plant productivity and soil carbon, nitrogen storage in alpine meadows in northern tibet, china[J]. Chinese Geographical Science, 2014,24(4):488-498.
[26] Frank D A, Mcnaughton S J, Tracy B F. The Ecology of the Earth′s Grazing Ecosystems[J]. BioScience, 1998,48(7):513-521.
[27] 李怡,韓國棟.放牧強度對內蒙古大針茅典型草原地下生物量及其垂直分布的影響[J].內蒙古農業大學學報:自然科學版,2011,32(2):89-92.
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
