路鄧葡萄球菌單寧酶基因的克隆、表達、純化與改造
2018-05-14 15:33:32劉鰩胡雪麗鐘秋梅等
中國測試 2018年9期
劉鰩 胡雪麗 鐘秋梅等


摘要:為提高路鄧葡萄球菌(Staphylococcus lugdunensis)單寧酶(Sl-tan)的活性,該文利用化學(xué)合成方法獲得Sl-tan基因,將該基因連接到重組表達質(zhì)粒pET43.1-A,再轉(zhuǎn)化到大腸桿菌感受態(tài)細(xì)胞BL21-DE3中進行表達,通過親和層析柱純化,以沒食子酸甲酯為底物進行酶活性測定以及酶學(xué)性質(zhì)分析,并基于生物信息學(xué)分析,結(jié)合定點突變技術(shù)對Sl-tan進行人工改造。結(jié)果顯示,獲得的重組單寧酶產(chǎn)量明顯增高,最高可達42 mg/L發(fā)酵液;酶學(xué)性質(zhì)研究顯示該酶在pH 8.0,溫度40℃時獲得的活性最高(40 U/mg);Ala460突變?yōu)镻r0460后的Sl-tan活性可提高82.5%。
關(guān)鍵詞:路鄧葡萄球菌;單寧酶;三明治結(jié)構(gòu);定點突變;沒食子酸甲酯
中圖分類號:TQ925 文獻標(biāo)志碼:A 文章編號:1674-5124(2018)09-0063-06
0引言
單寧是一種水溶性多酚化合物,在植物界中廣泛存在。單寧中含有豐富的碳源,但由于其含有大量的芳香環(huán)類結(jié)構(gòu),易于螯合金屬離子,并與蛋白聚合形成不可溶性沉淀,導(dǎo)致單寧難以被降解利用。一些微生物能夠表達單寧酶,將單寧水解為五倍子酸與葡萄糖,為微生物生長提供碳源以及能源物質(zhì)。單寧酶是已知的唯一能夠降解單寧的生物酶類,因此被廣泛應(yīng)用于食品行業(yè)、制藥行業(yè)以及動物飼料的生產(chǎn)中。
目前,單寧酶的生產(chǎn)主要通過產(chǎn)單寧酶菌株的液態(tài)深層發(fā)酵以及固體發(fā)酵兩種方法,耗時長、產(chǎn)量低、成本高、難以純化,且生產(chǎn)的單寧酶通常以粗酶或者菌體形式應(yīng)用,不利于單寧酶的應(yīng)用。隨著基因工程技術(shù)的發(fā)展,將單寧酶基因克隆,構(gòu)建表達質(zhì)粒,利用表達宿主進行表達的方法也已經(jīng)取得初步成效。Iwamoto等,首次將乳酸桿菌單寧酶(Lp-tan)基因通過基因擴增構(gòu)建Lp-tan重組表達質(zhì)粒,并在大腸桿菌DH5α中成功表達,但是純化后酶的產(chǎn)量與活性較低。在此基礎(chǔ)上,Wu等通過LIC-PCR的方式構(gòu)建了Lp-tan的重組表達質(zhì)粒,并優(yōu)化表達用宿主細(xì)胞,最終在大腸桿菌BL21-DE3中實現(xiàn)了Lp-tan的高產(chǎn)量表達,并保持了較高的酶活性。目前為止,已經(jīng)從米曲霉(Aspergillus oryzae)、乳酸桿菌(Lactobacillus plantarum)以及鏈霉菌(Streptomyces sviceus)中克隆了單寧酶基因,通過異源重組表達的方法生產(chǎn)單寧酶,取得了較好的結(jié)果。
本文通過化學(xué)合成獲得了路鄧葡萄球菌(Staphylococcus lugdunensis)的單寧酶基因Sl-tan,構(gòu)建原核表達質(zhì)粒,使其在大腸桿菌宿主中進行表達,并通過與Lp-tan的氨基酸序列比對分析,結(jié)合已經(jīng)報道的Lp-tan的結(jié)構(gòu)(PDB序列號:4JOC)、Lp-tan與底物沒食子酸乙酯的結(jié)構(gòu)(PDB序列號:4JOK)、Lp-tan與產(chǎn)物五倍子酸的結(jié)構(gòu)(PDB序列號:4JOH)對Sl-tan進行定點改造。改造后獲得的重組Sl-tan活性較改造之前提高了82.5%。
1材料與儀器
1.1材料
大腸桿菌DH5a、BL21-DE3菌株以及質(zhì)粒pET-43.1-A均為實驗室保存;限制性內(nèi)切酶BamHI和XhoI、T4 DNA連接酶購買自Thermo-Fisher;DNA聚合酶購買自TaKaRa;DNA膠回收試劑盒購買自QIAGEN;定點突變試劑盒購買自北京全式金;商品化米曲霉單寧酶(Wako,Japan);沒食子酸甲酯、單寧酸、異丙基硫代半乳糖苷(IPTG)購買自Sigma;咪唑購買自科龍(成都);10KD濃縮管購買自Millipore。
1.2儀器
超凈工作臺(蘇凈集團安泰公司SW-CZ-1F);蛋白純化儀(蘇州利穗);全自動高壓蒸汽消毒器(上海三申醫(yī)療器械有限公司YX280A);搖床(上海智誠);10mLHisTrapHP親和層析柱(博格隆);超聲破碎儀(寧波新芝);高速冷凍離心機(貝克曼,美國)。
2方法
2.1目的基因獲取
在NCBI數(shù)據(jù)庫中查找Sl-tan基因(GenBank:KU882097.1),通過化學(xué)合成的方法獲得完整的基因序列(擎科生物,成都)。
2.2重組載體的構(gòu)建
本文中利用的重組表達載體為改造后的pET43.1-A(僅保留N端His-tag,并在His-tag后引人煙草花葉病毒酶(TEV)的酶切位點,去除了原先質(zhì)粒上的S-tag以及NusA-tag),以合成的Sl-tan基因為模板,設(shè)計引物序列進行PCR擴增,上游引物序列為CGGATCC-ATGAAA
GACTTTCATATC-ACTCT,下游引物序列為CCTCGAG-CTATTTTTT-ATTAATACTTTCTACC(斜體為BamHI和XhoI的酶切位點及保護堿基),擴增條件為95℃30 s,52℃30 s,72℃2min,35個循環(huán)。擴增后的PCR產(chǎn)物與pET43.1-A質(zhì)粒同時用限制性內(nèi)切酶BamHI和XhoI進行雙酶切,酶切后用膠回收試劑盒純化。將純化后的雙酶切質(zhì)粒與PCR產(chǎn)物按照1:3的比例混合后,用T4 DNA連接酶在室溫下連接1h,轉(zhuǎn)化DH5a,涂含有卡那霉素的LB平板,37℃培養(yǎng)過夜后,挑取單克隆提質(zhì)粒,并送公司(擎科生物,成都)測序。
2.3誘導(dǎo)表達與純化
將測序正確的重組質(zhì)粒轉(zhuǎn)化表達用大腸桿菌感受態(tài)細(xì)胞BL21-DE3,涂含有卡那霉素的LB平板,37℃培養(yǎng)過夜后挑取單克隆接種于含有卡那霉素的LB液體培養(yǎng)基中,于37℃,220 r/min培養(yǎng)至OD600值為0.6左右時,加入終濃度為0.5 mmol/L的IPTG繼續(xù)誘導(dǎo)培養(yǎng)4 h,6000 r/min離心30 min,棄上清,收集菌體。
將收集的菌體用平衡緩沖液(20 mmol/L Tris-HCl,100 mmol/LNaCl,20 mmol/L咪唑,pH 8.0)重懸,超聲破碎15 min,離心后收集上清并過5μm孔徑濾膜,收集上清并用蛋白純化儀過10 mL HisTrapHP親和層析柱,用洗脫緩沖液(20 mmol/L Tris-HCl+100 mmol/L NaCl+300 mmol/L咪唑,pH8.0)梯度洗脫。
2.4 TEV酶切去除N末端組氨酸標(biāo)簽
載體pET43.1-A自身帶有18個氨基酸的組氨酸標(biāo)簽,在組氨酸標(biāo)簽后帶有煙草花葉病毒酶(TEV)的酶切位點。為了去除組氨酸標(biāo)簽對Sl-tan活性的影響,用TEV酶切去除組氨酸標(biāo)簽。將親和層析后的Sl-tan,用透析緩沖液(20 mmol/L Tris-HCl+100 mmol/L NaCl,pH 8.0)透析,去除咪唑,然后與TEV按照100:1的比例混合,4℃條件酶切過夜。經(jīng)酶切后的Sl-tan(加入20mmol/L咪唑)再次過組氨酸親和層析柱純化,收集穿透峰樣品。
2.5酶活性的測定
酶活性單位U的定義:在溫度為40℃,pH為8.0的條件下,每分鐘內(nèi)水解底物生成1μmol產(chǎn)物五倍子酸所需要的酶量,定義為一個酶活性單位U。
單寧酶可以水解沒食子酸甲酯以及單寧酸,釋放出五倍子酸。通過繞單寧和五倍子酸的特異反應(yīng),可以測定單寧酶的活性。以25 mmol/L沒食子酸甲酯作為底物,單寧酶可以水解沒食子酸甲酯(MG),生成五倍子酸,五倍子酸(GA)可以與繞單寧反應(yīng),用NaOH終止反應(yīng),測520 nm的吸光度,顯色強度與五倍子酸的量成正比。在700μL的反應(yīng)緩沖液(20 mmol/L Tris-HCl+100 mmol/L NaCl,pH 8.0)中加入0.1μg的單寧酶與40μL摩爾濃度為25 mmol/L的沒食子酸甲酯在40℃條件下溫育5 min,然后加入150μL質(zhì)量濃度為0.667%的繞單寧溶液再次在40℃下溫育5mm,隨后加入100μL摩爾濃度為500 mmol/L的NaOH溶液反應(yīng)5 min終止反應(yīng),用分光光度計檢測酶解液在520 nm下的吸光度值。
標(biāo)準(zhǔn)曲線制備:以0.125~1 mmol/L的五倍子酸系列標(biāo)準(zhǔn)溶液與繞單寧反應(yīng),測520nm的吸光度,以五倍子酸的濃度為橫坐標(biāo)對縱坐標(biāo)吸光度值作出標(biāo)準(zhǔn)曲線。
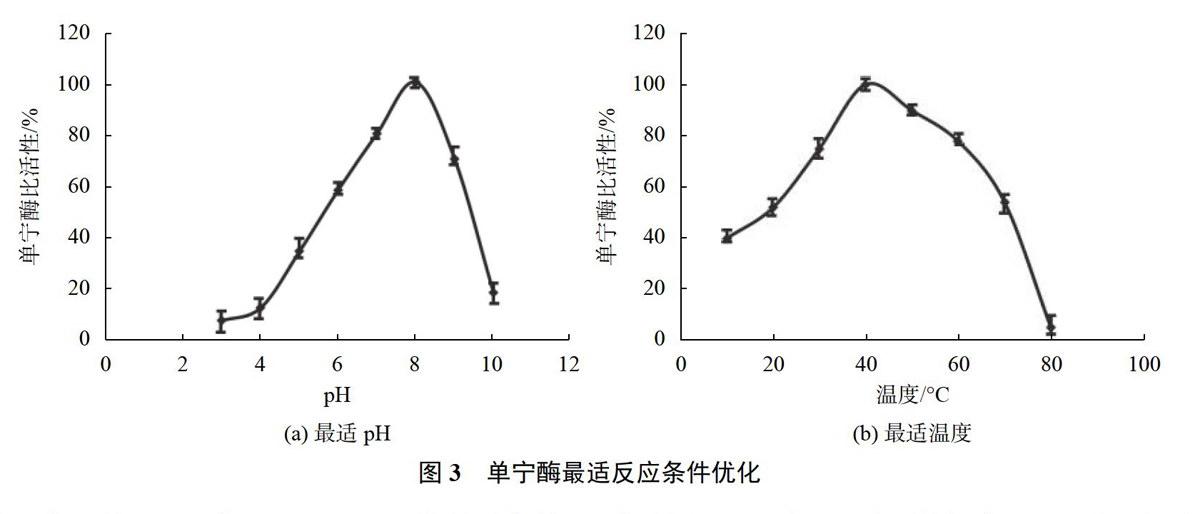
2.6溫度與pH對酶活性的影響
為了研究酶的最適反應(yīng)溫度與pH,將純化的單寧酶在10~80℃條件下以沒食子酸甲酯為底物,在pH值為8.0的條件測定酶的活性,以測得的最高活性值為100%作圖。在溫度為37℃時,配制pH值為3.0~10.0的緩沖液,以沒食子酸甲酯為底物檢測酶的活性,以測得最高活性數(shù)值為100%作圖。
2.7單寧酶的序列分析及定點突變
乳酸桿菌單寧酶具有較高的生物活性,且其三維結(jié)構(gòu)已經(jīng)被解析,并且根據(jù)其結(jié)構(gòu)闡明了單寧酶的水解機制。通過Expasy Align比對分析Lp-tan與Sl-tan單寧酶的差異,并結(jié)合已報到的乳酸桿菌單寧酶的結(jié)構(gòu)與水解機制,對差異位點進行定點突變,具體突變方法如下:
通過PCR的方法,以包含Sl-tan基因的質(zhì)粒為模板,按照定點突變試劑盒試劑盒要求,設(shè)計引物(上游引物:TTTAAA CGTAGCCAA-CAG-GAAA-ATGAAGT;下游引物:CTG-TTGGCTACGTTTTAA-ATCATC,斜體表示突變位點),進行PCR擴增(94℃30 s,60℃30 s,72℃8 min,20個循環(huán))。將獲得的PCR產(chǎn)物用DpnI酶處理,消化掉模板質(zhì)粒。將酶切后的PCR產(chǎn)物跑1.0%的瓊脂糖凝膠電泳,切膠后回收PCR產(chǎn)物,轉(zhuǎn)化大腸桿菌DH5α,涂平板后于37℃培養(yǎng)24 h后,挑取單克隆于LB培養(yǎng)基中,37℃,200 r/min,過夜培養(yǎng)。提質(zhì)粒,送測序,將測序結(jié)果正確的質(zhì)粒轉(zhuǎn)化大腸桿菌BL21-DE3感受態(tài)細(xì)胞,進行誘導(dǎo),表達、純化及活性測定,方法同前。
3結(jié)果
3.1目的基因PCR擴增及酶切鑒定結(jié)果
以合成的Sl-tan基因為模板,進行PCR,1.0%瓊脂糖凝膠電泳可見1800 bp左右的DNA片段,與預(yù)期一致(見圖1)。構(gòu)建好的重組質(zhì)粒進行測序,測序結(jié)果與NCBI數(shù)據(jù)庫登陸序列一致。
3.2蛋白表達與純化
用大腸桿菌BL21-DE3作為宿主表達蛋白并于20℃過夜誘導(dǎo)。收菌后,超聲破碎,將上清用蛋白純化儀進行純化。表達載體pET43.1-A,帶有N末端組氨酸標(biāo)簽,表達的目的蛋白前端含有MG-HHHHHHGTENLYFQGS氨基酸序列。破碎后上清過組氨酸親和層析柱,用高濃度咪唑梯度洗脫,在咪唑濃度為80~170 mmol/L之間出峰。酶切去除N末端組氨酸標(biāo)簽后,再次用鎳柱親和純化后得到純度95%以上的單寧酶。10%SDS-PAGE電泳驗證(見圖2),在分子量67 kD左右有明顯的單一目的條帶,與Sl-tan中單寧酶分子量大小相符,純化后,重組單寧酶的產(chǎn)量為42 mg/L菌液。
3.3溫度和pH值對酶活性的影響
在不同pH值和不同溫度條件下,測定純化后單寧酶的活性,如圖3(a)所示,在pH7~9時,酶可以保持相對較高的活性,在pH值8附近,酶的活性最高;如圖3(b)所示,在溫度30~60℃時,酶可以保持相對較高的活性,在40℃附近,酶的活性最高。
將純化后的Sl-tan在40℃,pH 8.0的最適條件下,以沒食子酸甲酯為底物進行活性測定,結(jié)果顯示帶有N端組氨酸標(biāo)簽的單寧酶活性為40 U/mg,去除標(biāo)簽后的單寧酶活性并未有明顯變化。
3.4 Sl-tan的改造
Sl-tan與Lp-tan的氨基酸序列比對結(jié)果顯示兩者只有21.9%的序列同源性,但在Sl-tan中具有與Lp-tan相同的單寧酶酶活性中心保守序列G-X-S-X-G-G(X代表任意氨基酸,見圖4)。在Lp-tan中Pr0356-底物-Ile206形成類似于三明治的結(jié)構(gòu),Pro356的苯環(huán)結(jié)構(gòu)能夠穩(wěn)定底物結(jié)合,但是在Sl-tan中,用于形成三明治結(jié)構(gòu)的Pro被Ala取代,從而破壞了三明治結(jié)構(gòu)。通過定點突變的方法在Sl-tan中重建三明治結(jié)構(gòu)(Ala460突變?yōu)镻r0460),活性測定結(jié)果顯示重建三明治結(jié)構(gòu)后的Sl-tan活陛為73 U/mg,與野生Sl-tan相比,酶活性提高82.5%(見表1)。
4討論
現(xiàn)階段,我國單寧酶的研究主要集中在產(chǎn)單寧酶的菌株篩選上,對產(chǎn)單寧酶的菌株進行誘變以獲取高產(chǎn)菌株。另外,對單寧酶的應(yīng)用也主要是以粗酶形式,或者直接以發(fā)酵后的菌體作為單寧酶使用,由于粗酶或者菌體直接作用可能存在細(xì)菌污染,限制了單寧酶的應(yīng)用。
前期研究中,本課題組通過基因重組表達的方式將乳酸桿菌中的單寧酶基因以及鏈霉菌中的單寧酶的基因進行克隆,并用大腸桿菌BL21-DE3進行表達,最終獲得高產(chǎn)量的單寧酶,并且保持了較高的酶活性。與傳統(tǒng)的單寧酶生產(chǎn)方式相比,更易于獲得高純度的單一的單寧酶,有利于單寧酶的工業(yè)化應(yīng)用,但是能夠用于重組表達的單寧酶仍然很少。
本文通過基因工程方法將Sl-tan基因進行克隆,并在大腸桿菌BL21-DE3中進行成功表達,獲得了高產(chǎn)量的Sl-tan(42 mg/L菌液)。對Sl-tan的活性研究顯示,其在pH 8.0,溫度40℃的條件下具有最高活性,但進一步的活性測定結(jié)果顯示其活性較低(40 U/mg)。在之前的研究中,我們報道了Lp-tan的晶體結(jié)構(gòu),并對單寧酶的水解機制進行了解析。通過對Sl-tan與Lp-tan的氨基酸序列比對發(fā)現(xiàn),兩種單寧酶的序列相似度只有21.9%,但是兩種單寧酶具有相同的活性中心序列,進一步比對發(fā)現(xiàn),在Lp-tan中形成的有利于底物結(jié)合的Pro356-底物-Ile206三明治的結(jié)構(gòu),在Sl-tan中被Ala460-底物-Ile271所替代(見圖4)。因此,本文利用定點突變技術(shù)在Sl-tan中將Ala460突變?yōu)镻ro460,重建三明治結(jié)構(gòu),活性測定結(jié)果顯示突變后的Sl-tan活性提高了82.5%,研究結(jié)果也進一步說明了在單寧酶中形成的三明治結(jié)構(gòu)有利于底物沒食子酸甲酯的結(jié)合。
5結(jié)束語
本研究通過化學(xué)合成的方法獲得了Sl-tan的基因,并構(gòu)建原核表達質(zhì)粒使其在大腸桿菌中表達,獲得了高產(chǎn)量、高純度的Sl-tan。對Sl-tan定點突變(Ala460突變?yōu)镻ro460)重建三明治結(jié)構(gòu)后,使Sl-tan的活性提高了82.5%,使其能夠更好地應(yīng)用于工業(yè)化生產(chǎn)實際。
