大蒜精油和發酵劑對熏馬腸中生物胺及微生物分布的影響
2018-05-25 00:52:40李彬彬張雅晴趙利利盧士玲
食品科學 2018年9期
李彬彬,張雅晴,趙利利,毛 順,盧士玲*
熏馬腸是新疆哈薩克民族特色的傳統肉類制品,是一種營養豐富、蛋白質含量極高、脂肪含量少但多不飽和脂肪酸所占比例高的發酵肉制品。熏馬腸作為優質獨特的功能性食品,具有很好的銷售前景,但由于其在加工成熟及貯藏過程中產生的生物胺累積,造成了一定的食品安全隱患[1]。
食物中存在生物胺的累積現象,主要可能由于十分充足的前體物質的存在,還有許多含有氨基酸脫羧酶基因的細菌以及能夠促進其生長并且對脫羧酶活性有促進作用的環境因素造成[2]。在所有胺類化合物中,組胺和酪胺被確認毒理作用最強。當它們作用于機體時,會對人類的精神狀態、血管活性造成影響,甚至導致嚴重的毒性反應[3]。此外生物胺具有良好的熱穩定性,食品加工和制備過程中的高溫對其破壞力有限,目前只能通過預防和監測的方式來有效控制生物胺在食品加工中的形成從而延長食品鏈[4]。
據目前大量研究結果來看,發酵劑對發酵香腸中生物胺的產生具有良好的控制作用[5],如乳酸菌和凝固酶陰性葡萄球菌[6]。乳酸菌在原料肉中數量極低(3~4(lg(CFU/g))),但它能夠在很短時間內提高乳酸菌的數量,使其在發酵初期成為熏馬腸中的優勢菌株(8(lg(CFU/g))),葡萄球菌能夠在發酵過程中分解原料形成風味化合物[7]。乳酸菌分解產生大量乳酸或有機酸導致pH值在短時間內快速降低,抑制了腐敗微生物的生長,從而減少生物胺的積累[8]。添加香辛料和天然植物提取物對原料肉進行腌制[9],也是目前控制生物胺產生的重要途徑,它能夠使熏馬腸中的初始微生物的數量和種類得到有效控制。
微生物宏基因組分類測序是根據對被檢樣品中微環境的基因序列實施檢測,能夠高效準確地辨別其中微生態物種的類別以及相對豐度。宏基因組學目前已被廣泛應用到發酵制品的發酵加工過程中,對微生物多樣性和動態進行研究。新疆熏馬腸發酵成熟時期涉及多菌種共同作用,分化發酵肉中微生物種類以及在各時期的變化規律和機理,對于深入研究其發酵機理和構建發酵時期的監測手段和控制途徑具有舉足輕重的意義。本實驗將利用宏基因組學的方法對熏馬腸發酵成熟時期的細菌群落的構成和多樣性進行詳細監測,旨在為發酵劑和大蒜精油對熏馬腸發酵成熟過程中優勢菌群的發掘和腐敗微生物的抑制提供一定的理論依據。
1 材料與方法
1.1 菌株、材料與試劑
屎腸球菌(Enterococcus faecium)KT923202、糞腸球菌(E. faecalis)KT923193均為本實驗室從熏馬腸中分離篩選。
馬肉購自石河子牧民家庭;輔料(天然動物腸衣、食鹽、煙熏液)均為市售。
改良后的甘露醇高鹽瓊脂培養基(manitol salt agar,MSA)、結晶紫中性紅膽鹽瓊脂培養基(violet red bile agar,VRBA)、MRS培養基 北京奧博星生物試劑公司;發酵劑 科漢森菌種有限公司;大蒜精油仲景大廚房股份有限公司。
1.2 儀器與設備
甲醇、乙腈(均為色譜純) 天津福晨化學試劑廠;生物胺標準品(色胺、苯乙胺、腐胺、尸胺、組胺、酪胺、亞精胺、精胺、丹磺酰氯(dansyl chloride,DNS-Cl))美國Sigma公司。
T25型勻漿機 德國IKA公司;BagMixer 400VW型均質器 法國Interscience公司;9020-0094 KBF 240型恒溫恒濕培養箱 德國Binder公司;高速冷凍離心機 美國貝克曼公司。
1.3 方法
1.3.1 新疆熏馬腸的制作
配方:瘦肉80%(質量分數,下同)、肥肉20%、食鹽2.5%、白糖2%、亞硝酸鈉0.02%、胡椒粉0.1%、花椒粉0.15%、姜粉0.2%、味精0.1%、五香粉0.1%、八角0.1%、煙熏液1%、大蒜精油0.025%、發酵劑0.025%。
工藝流程:原料肉處理(紫外燈照射30 min)→修整、切丁(切小拇指上指尖大小)→配料→腌制(4 ℃,1 d)→接種→灌腸→發酵(18 ℃,相對濕度90%~95%,2 d)→成熟(12 ℃,相對濕度70%~75%,25 d)→成品。
分別在第0、3、7、14、21、28天取樣,每次取一根,每根香腸分裝成5 節,對微生物數量、水分含量、pH值測定,置于-20 ℃冷凍保藏,以備生物胺的測定。
1.3.2 實驗設計
將1.3.1節處理好的原料肉均勻地分為8 組,其中4 組接種濃度約為103CFU/mL的屎腸球菌和糞腸球菌菌液作為產胺菌接種劑,另外4 組不接種產胺菌。然后在模擬發酵條件下進行熏馬腸的發酵和成熟,并在發酵過程中的第0、3、7、14、21、28天分別取樣,進行實驗分析。
對原料馬肉分別做如下處理:第1組(OJ)添加大蒜精油和接種產胺菌;第2組(CK)空白對照組;第3組(S)只添加發酵劑;第4組(SO)添加發酵劑和大蒜精油;第5組(O)只添加大蒜精油;第6組(SOJ)添加發酵劑和大蒜精油及接種產胺菌;第7組(SJ)添加發酵劑和接種產胺菌;第8組(J)接種產胺菌。每組腐敗菌的接種量均約為103CFU/g。發酵劑為清酒乳桿菌和肉糖葡萄球菌,添加量為106CFU/g。
1.3.3 微生物計數
取10 g香腸,加入到90 mL無菌生理鹽水中,在室溫下200 r/min搖床培養30 min。然后根據微生物實驗操作書上的要求進行規范操作,吸取適當濃度菌液于對應固體培養基進行均勻涂布。乳酸菌用MRS培養基,在37 ℃條件下培養72 h;葡萄球菌/微球菌用MSA培養基,37 ℃培養48 h;腸細菌用VRBA培養基,30 ℃培養48 h。
1.3.4 樣品pH值測定
無菌條件下取樣品20 g于180 mL滅菌生理鹽水,裝入無菌拍打袋中。使用拍打機,強度調至4拍打20 min,直至樣品完全散開在無菌生理鹽水中,上清液用pH計進行測定。
1.3.5 樣品生物胺含量測定
參考文獻[10]的方法,取5 g熏馬腸樣品加入20 mL的0.4 mol/L的高氯酸,取上述樣品1 mL,加入200 μL 2 mol/L NaOH溶液使溶液呈堿性,再加入300 μL飽和NaHCO3溶液進行緩沖,然后再加入2 m L 10 mg/mL的DNS-Cl丙酮溶液,使之在黑暗條件40 ℃反應,45 min后加入100 μL的NH3·H2O終止反應,去除殘留的DNS-Cl溶液。用乙腈定容到5 mL,用0.22 μm濾膜過濾后裝入樣品瓶待測。
色譜條件:C18色譜柱(250 mm×4.6 mm,5 μm);流動相A為水,流動相B為乙腈;流速為0.9 mL/min;進樣量:20 μL;柱溫:30 ℃;檢測波長:254 nm。梯度洗脫程序為:0.1 min、35% A;5.0 min、25% A;44 min、0% A;55 min、35% A。1.3.6 宏基因組學測序
將8 組熏馬腸發酵成熟所有時期的共41 個樣品,無菌條件下取樣送至華大基因測序公司進行宏基因組學測序。對樣品純度進行檢測;檢測合格的樣品構建文庫:回收目的Amplicon片段,用T4 DNA Polymerase、Klenow DNA Polymerase和T4 PNK將打斷形成的黏性末端修復成平末端,再通過3’端加堿基“A”,使得DNA片段能與3’端帶有“T”堿基的特殊接頭連接。也可通過對具有測序接孔的雙Index融合引物進行設計并合成,以DNA為模板,對PCR引物進行融合。磁珠篩選目的Amplicon片段,最后,用合格的文庫進行cluster制備和測序。用下機得到的數據進行相應的生物信息分析。
下機數據經過過濾,濾除低質量的reads,剩余高質量的Clean data方可用于后期分析。通過reads之間的Overlap關系將reads拼接成Tags。在規定的一定相似度下將Tags集聚合成操作分類單元(operational taxonomic units,OTU),最后將OTU與數據庫比對,對OTU進行物種注釋。基于OTU和物種注釋結果進行樣品物種復雜度分析以及組間物種差異分析。
1.4 數據統計分析
所有數據從液相分析平臺導出,用Excel 2010建立數據庫,并用DPS 7.05版進行鄧肯氏多重比較分析。
2 結果與分析
2.1 pH值分析
pH值的下降是熏馬腸在發酵成熟過程中的一個顯著變化,新鮮馬肉的初始pH值為5.78。由表1可知,8 組熏馬腸在發酵初期(0~3 d),pH值迅速下降,接種發酵劑組熏馬腸的酸化程度要顯著高于對照組的熏馬腸(P<0.05),主要是由于發酵劑或接種的乳酸菌等微生物作用于內源或添加的糖等碳水化合物,使其分解為乳酸、醋酸或其他有機酸,從而導致pH值降低,pH值降低可以有效抑制雜菌的生長。因此,未接種的對照組pH值下降過程不明顯,發酵結束后pH值為5.3。發酵結束后pH值下降緩慢,這是由于成熟溫度為18 ℃,要低于發酵溫度,導致一些微生物生長速率緩慢,產酸能力受到抑制。直到成熟期21 d,pH值開始回升,主要原因是在成熟過程中,水分蒸發流失,含氮化合物被乳酸菌利用作為能源,蛋白酶分解蛋白質產生堿性緩沖物質如生物胺的積累。在成熟結束后,各組樣品的pH值大多處于4.64~4.92之間,依然能夠為食品提供一個較低的酸性環境,能夠很好地抑制細菌的氨基酸脫羧酶活性,從而控制生物胺的產生。Fanco等[11]報道,pH 5左右的酸性條件有利于維持發酵香腸貨架期的穩定性。但這一結果要低于Lu Shiling等[9]終產品中的pH值,可能是由于所使用的馬肉品質間的差異所致。通過比較分析可得,添加發酵劑組pH值下降幅度顯著(P<0.05)大于未接種發酵劑組,并且在終產品中的pH值顯著低于未接種發酵劑組(P<0.05);與接種發酵劑相比,添加大蒜精油對熏馬腸pH值的作用效果并不明顯(P>0.05),這與Bozkurt[12]的研究結果較為相似。
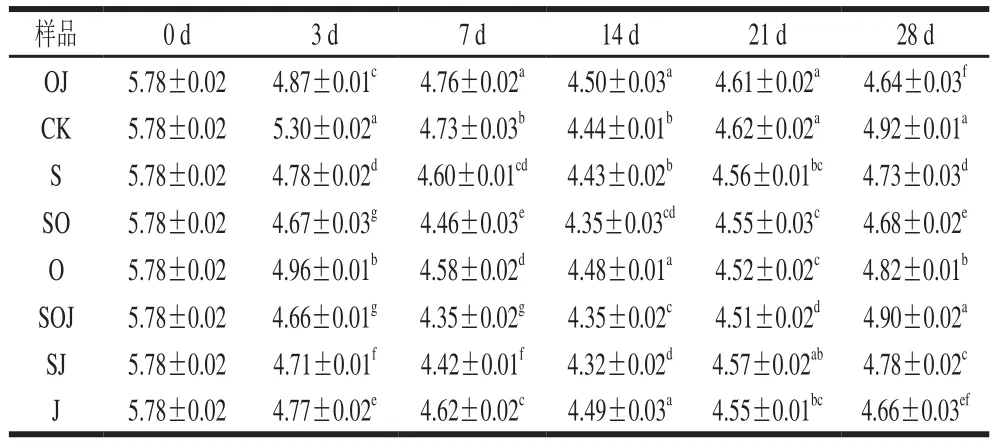
表1 新疆熏馬腸在成熟過程中pH值變化情況Table 1 Change in pH during the ripening of smoked horsemeat sausages
2.2 微生物分布分析2.2.1 乳酸菌分析
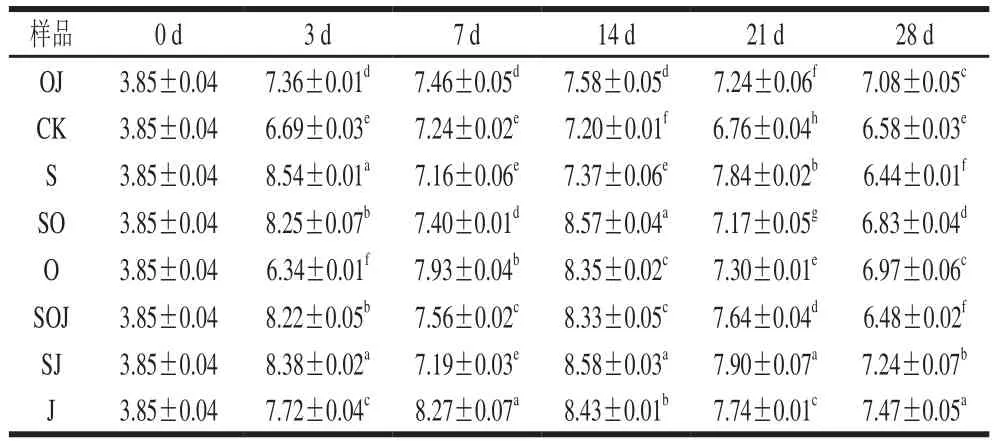
表2 新疆熏馬腸在成熟過程中乳酸菌生長情況(n=6)Table 2 Growth of lactic acid bacteria during the ripening of smoked horsemeat sausages (n= 6)lg(CFU/g)
由表2可知,不同處理的樣品在發酵成熟過程中乳酸菌數量存在差異。在發酵結束時(0~3 d),除CK和O組,其他香腸中的乳酸菌都迅速由原料肉中的3.85(lg(CFU/g))增長到7.36~8.38(lg(CFU/g)),通常情況下生鮮肉中的乳酸菌數量在3~4(lg(CFU/g))[13],其中接種發酵劑組的熏馬腸,乳酸菌數量平均都在8.22(lg(CFU/g)),顯著高于其他未接種發酵劑的熏馬腸(P<0.05)。此結果與Mokhtar等[5]研究結果相比,對照組乳酸菌數量高1.587(lg(CFU/g)),接種發酵劑組平均高1.159(lg(CFU/g)),這可能是由于原料肉品質間的差異引起的,也可能是發酵劑中菌種活性不同所致。在熏馬腸的整個成熟階段,加入大蒜精油對接種產胺菌組的乳酸菌數量有一定抑制作用。在未接種發酵劑的樣品中,乳酸菌依然成為了優勢菌,之前許多國內外學者已有報道[14]。在成熟結束后(第28天),每組熏馬腸中的乳酸菌數量在6.44~7.47(lg(CFU/g))之間,與國際上建議的乳酸菌在功能性食品中的數量范圍6~8(lg(CFU/g))相符[15],說明產品具有一定的可信度。2.2.2 微球菌和葡萄球菌分析
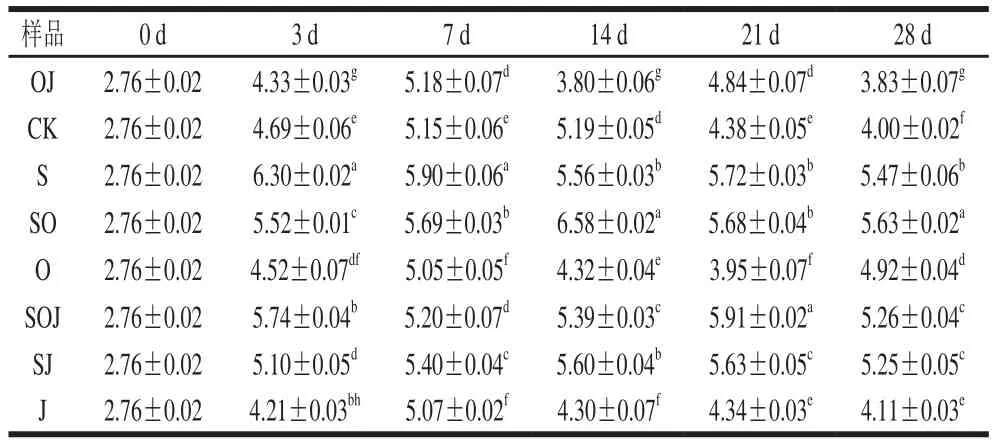
表3 新疆熏馬腸在成熟過程中微球菌和葡萄球菌生長情況(n=6)Table 3 Growth of Micrococcus and Staphylococcus during the ripening of smoked horsemeat sausages (n= 6)lg(CFU/g)
由于微球菌和葡萄球菌具有解朊作用和分解脂肪活性,因此與乳酸菌一起作為發酵劑被廣泛應用于發酵香腸中。為研究發酵劑和大蒜精油對微球菌和葡萄球菌數量的影響,分別對8 組熏馬腸樣品中檢測出的微球菌和葡萄球菌數量進行統計分析及多重比較,結果見表3。接種發酵劑組的熏馬腸中微球菌和葡萄球菌數量遠遠高于其他未接種發酵劑組(P>0.05),數量在5.5(lg(CFU/g))左右,而未接種發酵劑的熏馬腸中微球菌和葡萄球菌數量在4.5(lg(CFU/g))左右,接種發酵劑組比未接種發酵劑組高1(lg(CFU/g))左右。該結果與Gücüko?lu等[16]研究接種不同發酵劑的發酵香腸在發酵結束后,微球菌和葡萄球菌數量在5.66~5.80(lg(CFU/g))相近。微球菌和葡萄球菌在發酵階段雖然數量增長的幅度遠低于乳酸菌,也可能是乳酸菌的快速產酸不利于其增殖,但接種發酵劑對微球菌和葡萄球菌的快速增殖還是有一定促進作用。到熏馬腸成熟的第21天時,添加大蒜精油對添加發酵劑的熏馬腸中微球菌和葡萄球菌數量沒有顯著抑制作用。在熏馬腸終產品中,各組樣品中的微球菌和葡萄球菌數量在3.83~5.63(lg(CFU/g))之間,與一些學者報道的發酵香腸中微球菌數量相近(4.1~5.9(lg(CFU/g)))[17]。
2.2.3 腸細菌分析
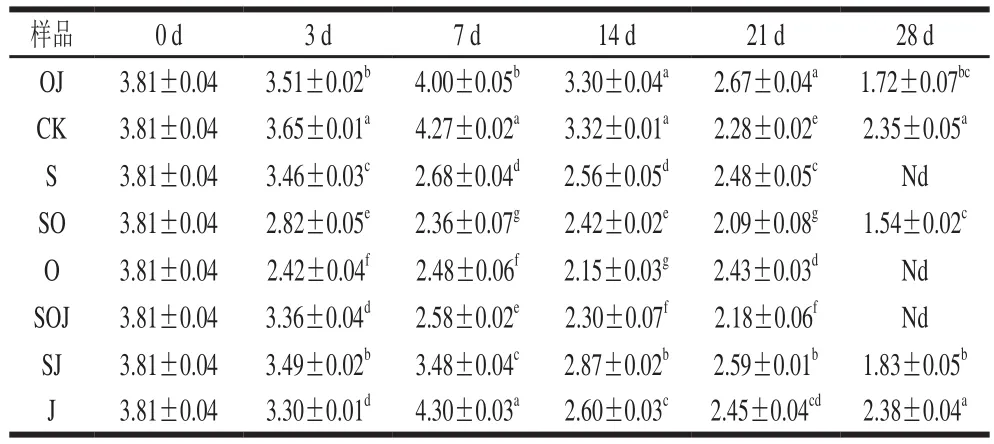
表4 熏馬腸在成熟過程中腸細菌生長情況(n=6)Table 4 Growth of Enterobacteria during the ripening of smoked horsemeat sausages (n= 6)lg(CFU/g)
腸細菌比起乳酸菌、微球菌和葡萄球菌,可以說是一類不受歡迎的菌種,大量的腸細菌存在于發酵香腸時可同時產生尸胺和腐胺。其常在肉中出現,如果在不適宜的環境下加工肉制品,很可能導致致病性危害以及對發酵香腸的感官品質造成影響[18];同時它也能通過水解蛋白、分解脂肪和分解糖等作用為發酵香腸貢獻芳香化合物[19]。在熏馬腸成熟時期中取樣檢測腸細菌數量,從表4可以看出,各組在發酵結束后,腸細菌數量變化不大,添加大蒜精油組腸細菌數量顯著低于未添加大蒜精油組(P<0.05)。到熏馬腸成熟第7天時,腸細菌的數量出現了兩種變化情況,添加大蒜精油和接種發酵劑組熏馬腸中的腸細菌數量顯著減少,且添加大蒜精油組比接種發酵劑組減少效果更顯著(P<0.05),而其余各組熏馬腸中的腸細菌數量均有一定程度的增長,說明大蒜精油對腸細菌的生長具有一定的抑制效果,而發酵劑在熏馬腸成熟階段對減少腸細菌數量上也做出一定的貢獻,這與Simion等[20]研究結果相符。成熟第28天時,各組熏馬腸樣品中的腸細菌數量都相對較低,說明使用的原料肉符合良好的衛生標準,添加發酵劑和精油對腸細菌有顯著的抑制作用,但接種發酵劑和添加大蒜精油處理組之間腸細菌數量沒有顯著性的差異(P>0.05)。
2.3 微生物多樣性分析
通過與數據庫進行比對,對OTU進行物種分類并在科分類等級對各個樣品作物種分布柱狀圖。圖1為41 個熏馬腸樣品在科水平中物種相對豐度柱狀圖,不同色塊寬度表示不同物種相對豐度比例。如圖1所示,在科水平上除SJ-21其他所有樣品乳桿菌科(Lactobacillaceae)相對豐度占細菌類群的50%以上,其中S-3、OJ-7、O-14、SO-14的相對豐度高達90%,說明乳酸菌的確在熏馬腸整個發酵成熟過程中是優勢菌群。而假單胞菌科(Pseudomonadales)在SJ-21樣品中占主導地位相對豐度約為60%,其次在SOJ-7中也出現一定比例,說明這兩組樣品在加工成熟過程中有可能受到污染,但在成熟結束時,在所有熏馬腸樣品中都沒有發現假單胞菌的存在,可能是水分缺失,不利于其生長。由表5中可以看出,SOJ-7與同一時期其他樣品相比,酪胺含量較高,達到82.40 mg/kg,而SJ-21組產生的尸胺和酪胺的含量均顯著高于其他組,Li Miaoyun等[21]也證實了假單胞菌通常是腐胺、酪胺和組胺的產生菌,這與本實驗所研究結論相符。除此之外,葡萄球菌科(Staphylococcaceae)在J-3、SJ-14、O-21、S-21、SO-21、OJ-28和SO-28這7 個樣品中相對豐度相對較高,約為10%~20%,由此可以看出,葡萄球菌常出現在發酵肉制品的成熟后期,是為發酵香腸的風味貢獻產香物質的主要菌群。Essid等[7]研究表明,葡萄球菌在發酵香腸中有兩大貢獻,一是酸化作用,二是反硝化作用,如脂解作用和蛋白質水解作用,同時還能生成低分子質量化合物如肽類、氨基酸等賦予發酵肉香氣成分。
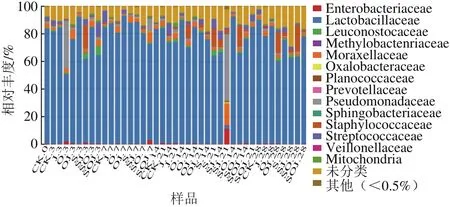
圖1 樣品科分類水平中物種相對豐度柱狀圖Fig. 1 Relative abundance of sequences assigned to families
2.4 生物胺含量分析
如表5所示,每種樣品中各種生物胺數據的變化可能要結合一些能夠影響生物胺產生的因素進行解釋分析,如原料肉的新鮮程度和衛生質量、加工和成熟階段引起的微生物種類和數量的變化等。
由于生物胺存在于發酵香腸對消費者健康造成潛在的威脅,因此酪胺、組胺、尸胺和腐胺已被建議作為發酵肉制品受腐敗微生物污染的標志[22]。許多研究表明,尸胺和腐胺是肉制品中主要的胺類[23],雖然它們通常不被認為具有毒性作用,但是它們能夠增強一些芳香族胺的毒性作用,如酪胺和組胺;此外,它們能夠與一些胺氧化酶反應,從而降低氧化酶降解生物胺的活性。腐胺主要是由假單胞菌產生的,尸胺主要是由腸桿菌產生的[24]。本研究中腐胺和尸胺的結果如表5所示,在0~3 d內,兩種胺的含量平均都低于50 mg/kg,腐胺含量到成熟結束也一直保持這樣的水平,該值被Ladero等[25]報道作為安全限量。而在熏馬腸成熟后期(第28天),與空白組相比,添加大蒜精油可使尸胺和腐胺含量減少75.50%和37.98%,接種發酵劑并添加大蒜精油可使尸胺和腐胺含量分別減少75.56%、70.34%,說明大蒜精油和發酵劑的結合使用能夠大幅度抑制這兩種生物胺的產生。從表5可看出,接種發酵劑的S組對腐胺的影響不是很大,但Zhang Qilin等[26]研究表明添加不產胺的乳酸菌作為發酵劑可以降低腐胺和尸胺的積累,并且能夠快速產酸,抑制不受歡迎的細菌如假單胞菌和大腸桿菌等的產生。而添加大蒜精油的O和SO組腐胺產生的數量要顯著低于其他組(P<0.05),說明可能是大蒜精油對鳥氨酸脫羧酶活性具有抑制作用,也可能是大蒜精油和發酵劑聯合使用阻礙了腐胺的產生。

表5 新疆熏馬腸中生物胺含量Table 5 Concentrations of biogenic amines in smoked horsemeat sausages mg/kg
從表5中可得,在熏馬腸發酵成熟的各個時期,接種發酵劑和添加大蒜精油組(S和SO組),尸胺含量比對照組(CK)平均減少了60%以上,此結果高于Simion等[20]研究在發酵香腸僅接種發酵劑對尸胺含量的影響,結果說明尸胺的形成受發酵劑的影響較大,而將大蒜精油與發酵劑相結合,對產生尸胺的微生物起到了良好的抑制效果。朱志遠[27]研究稱pH值對氨基酸脫羧酶活性具有很大的影響。在熏馬腸中快速降低pH值,能夠抑制產胺菌的生長,特別對于腸細菌作用最顯著,因此其尸胺含量也顯著低于其他胺類。而接種產胺菌組對尸胺的產生影響不大,說明屎腸球菌和糞腸球菌不是主要產生尸胺的菌種。
已有大量研究表明,組胺的產生與尸胺和腐胺有關,尤其與尸胺關系更為密切,因為它們都主要由腸細菌產生[28]。從表5可以看出,各個時期的8 組熏馬腸樣品中組胺含量都不超過50 mg/kg,遠低于一些學者提出的發酵肉制品中組胺的最低限量100 mg/kg[29]。接種發酵劑和添加大蒜精油的熏馬腸中組胺的含量顯著少于空白對照組(P<0.05),且在第28天時,組胺含量相較對照組減少了68.88%,說明發酵劑和大蒜精油在抑制組胺形成過程中具有一定作用。
酪胺在食物中引起中毒的劑量范圍為100~800 mg/kg[30],而本實驗8 組熏馬腸樣品只有J組在發酵結束時酪胺的含量最高為117 mg/kg,而且即使在接種發酵劑或添加大蒜精油的樣品中,酪胺的含量也都在75 mg/kg以上,說明酪胺主要是由屎腸球菌和糞腸球菌等乳酸菌產生的,Pircher等[31]研究也認為,酪胺的累積通常與一些污染菌如非發酵型乳酸菌有關,特別是腸球菌屬;Linares等[32]也曾報道稱酪胺通常是由一些具有生物胺脫羧酶活性的乳酸菌屬產生的,如彎曲乳桿菌、短乳桿菌等。高濃度的酪胺通常在發酵階段產生,并且與Parente等[33]報道相似。在成熟第28天,接種發酵劑和添加大蒜精油的樣品酪胺平均含量為30 mg/kg,而接種產胺菌組酪胺含量平均在78 mg/kg,都沒超過酪胺在食品中規定的限量。
通常苯乙胺是伴隨著大量的酪胺出現的[34],有些學者認為苯乙胺是有非特異活性的酪氨酸脫羧酶形成的,而Montel等[35]則報道稱該胺是由腸球菌和乳酸菌(彎曲乳桿菌、屎腸球菌和糞腸球菌)產生的。由表5中可以看出,原料肉中苯乙胺的含量非常低,說明該生物胺不是內源性胺。而大量的苯乙胺的形成是在發酵結束第3天時,接種產胺菌組中的苯乙胺含量要顯著高于未接種產胺菌組(P<0.05),經對比發現,大蒜精油對產胺菌產生物胺的抑制作用要優于發酵劑,而聯合使用效果最佳。在Gücüko?lu等[16]研究中也證實了接種發酵劑能夠阻礙苯乙胺的形成。在成熟結束后,接種產胺菌平均可產生苯乙胺39.78 mg/kg、酪胺5.35 mg/kg,只添加大蒜精油或接種發酵劑可使苯乙胺含量分別減少60.88%和68.57%,使酪胺含量分別減少61.31%和57.38%,接種發酵劑和添加大蒜精油組的熏馬腸中苯乙胺含量在11.55~18.20 mg/kg之間,稍高于Gücüko?lu等[16]的研究結果,但與Bover-Cid等[36]測定的苯乙胺在發酵香腸中的含量5~45 mg/kg相近。
精胺和亞精胺是天然存在的胺類,它們會經常存在于新鮮的肉中。它們既會出現在用于制作香腸的瘦肉中,也會存在于脂肪里。并且它們受香腸成熟過程中的影響變化非常小,精胺和亞精胺的含量在加工成熟過程中幾乎保持不變,甚至精胺會被某些微生物當作氮源利用導致含量降低。從表5中可以看出,所有組的熏馬腸樣品中精胺和亞精胺的含量都相當低,而且各組之間的差異性不顯著(P>0.05),說明使用的原料肉中自身所含的精胺和亞精胺就相對較少。
3 結 論
將大蒜精油和發酵劑聯合使用能夠很好地減少熏馬腸中尸胺、腐胺、組胺、酪胺和苯乙胺的產生,對控制生物胺在熏馬腸中的含量起到非常顯著的作用。添加大蒜精油可使尸胺和腐胺含量減少75.50%和37.98%;接種發酵劑并添加大蒜精油尸胺、腐胺和組胺含量平均減少75.56%、70.34%和68.88%。接種產胺菌平均可產生苯乙胺39.78 mg/kg、酪胺5.35 mg/kg;只添加大蒜精油或接種發酵劑可使苯乙胺含量分別減少60.88%和68.57%,使酪胺含量分別減少61.31%和57.38%;而復合添加物可苯乙胺和酪胺含量平均減少75.17%和63.30%。
將41 個樣品進行聚類分析共產生276 個OTU,說明熏馬腸中含有豐富的微生物物種,通過樣品科分類水平中物種分析,得出乳酸菌在熏馬腸整個加工過程都是優勢菌,而葡萄球菌在成熟后期數量較多。大蒜精油對一些腐敗菌如大腸桿菌、假單胞菌具有一定的抑制作用,加之發酵劑中的乳酸菌迅速產酸,從而減少了生物胺在熏馬腸中的積累。
參考文獻:
[1] AYHAN K, KOLSARICI N, ?ZKAN G A. The effects a starter culture on the formation of biogenic amines in Turkish soudjoucks[J]. Meat Science, 1999, 53(3): 183-188. DOI:10.1016/S0309-1740(99)00046-7.
[2] GARAI G, DUE?AS M T, IRASTORZA A, et al. Biogenic amines in natural ciders[J]. Journal of Food Protection, 2006, 69(12): 3006-3012.
[3] SPANO G, RUSSO P, LONVAUD-FUNEL A, et al. Biogenic amines in fermented foods[J]. European Journal of Clinical Nutrition, 2010,64: 95-100. DOI:10.1038/ejcn.2010.218.
[4] ALVAREZ M A, MORENO-ARRIBAS M V. The problem of biogenic amines in fermented foods and the use of potential biogenic amine-degrading microorganisms as a solution[J]. Trends in Food Science & Technology, 2014, 39(2): 146-155. DOI:10.1016/j.tifs.2014.07.007.
[5] MOKHTAR S, MOSTAFA G, TAHA R, et al. Effect of different starter cultures on the biogenic amines production as a critical control point in fresh fermented sausages[J]. European Food Research &Technology, 2012, 235(3): 527-535. DOI:10.1007/s00217-012-1777-9.
[6] LEROY F, VERLUYTEN J, VUYST L D. Functional meat starter cultures for improved sausage fermentation[J]. International Journal of Food Microbiology, 2006, 106(3): 270-285. DOI:10.1016/j.ijfoodmicro.2005.06.027.
[7] ESSID I, HASSOUNA M. Effect of inoculation of selected Staphylococcus xylosus and Lactobacillus plantarum strains on biochemical, microbiological and textural characteristics of a Tunisian dry fermented sausage[J]. Food Control, 2013, 32(2): 707-714.
[8] LU S L, XU X L, ZHOU G H, et al. Effect of starter cultures on microbial ecosystem and biogenic amines in fermented sausage[J]. Food Control, 2010, 21(4): 444-449. DOI:10.1016/j.foodcont.2009.07.008.
[9] LU Shiling, JI Hua, WANG Qingling, et al. The effects of starter cultures and plant extracts on the biogenic amine accumulation in traditional Chinese smoked horsemeat sausages[J]. Food Control,2015, 50: 869-875. DOI:10.1016/j.foodcont.2014.08.015.
[10] 馬宇霞, 盧士玲, 李開雄, 等. 熏馬腸中生物胺氧化酶菌株的篩選與鑒定[J]. 現代食品科技, 2014, 30(5): 49-55. DOI:10.13982/j.mfst.1673-9078.2014.05.002.
[11] FANCO I, PRIETO B, CRUZ J M, et al. Study of the biochemical changes during the processing of Androlla, a Spanish dry-cured pork sausage[J]. Food Chemistry, 2002, 78(2): 339-345. DOI:10.1016/S0308-8146(02)00118-8.
[12] BOZKURT H. Utilization of natural antioxidants: green tea extract and Thymbra spicata oil in Turkish dry-fermented sausage[J]. Meat Science, 2006, 73(3): 442-450. DOI:10.1016/j.meatsci.2006.01.005.
[13] 盧士玲, 李開雄, 徐幸蓮, 等. 傳統香腸中產生物胺腸細菌和乳酸菌分離方法的研究[J]. 食品與發酵工業, 2012, 38(8): 47-52.DOI:10.13995/j.cnki.11-1802/ts.2012.08.031.
[14] ELíAS M, CARRASCOSA A V. Characterisation of the Paio do Alentejo, a traditional Portuguese Iberian sausage, in respect to its safety[J]. Food Control, 2010, 21(1): 97-102. DOI:10.1016/j.foodcont.2009.04.004.
[15] SANTIAGO R M, ALBERTO M, MARíA JOSé B, et al. Application of Lactobacillus fermentum HL57 and Pediococcus acidilactici SP979 as potential probiotics in the manufacture of traditional Iberian dryfermented sausages[J]. Food Microbiology, 2011, 28(5): 839-847.DOI:10.1016/j.fm.2011.01.006.
[16] GüCüKO?LU A, ?ZLEM KüPLüLü. The effect of different starter cultures and ripening temperatures on formation of biogenic amine in Turkish fermented sausages[J]. European Food Research &Technology, 2010, 230(6): 875-884. DOI:10.1007/s00217-010-1220-z.
[17] MARTíN A, COLíN B, ARANDA E, et al. Characterization of Micrococcaceae isolated from Iberian dry-cured sausages[J]. Meat Science, 2007, 75(4): 696-708. DOI:10.1016/j.meatsci.2006.10.001.
[18] HUGAS M, GARRIGA M, AYMERICH M T. Functionalty of enterococci in meat products[J]. International Journal of Food Microbiology, 2003, 88: 223-233. DOI:10.1016/S0168-1605(03)00184-3.
[19] SARANTINOPOULOS P, ANDRIGHETTO C, GEORGALAKI M D, et al. Biochemical properties of enterococci relevant to their technological performance[J]. International Dairy Journal, 2001, 11(8):621-647. DOI:10.1016/S0958-6946(01)00087-5.
[20] SIMION A M C, VIZIREANU C, ALEXE P, et al. Effect of the use of selected starter cultures on some quality, safety and sensorial properties of Dacia sausage, a traditional Romanian dry-sausage variety[J]. Food Control, 2014, 35(1): 123-131. DOI:10.1016/j.foodcont.2013.06.047.
[21] LI Miaoyun, TIAN Lu, ZHAO Gaiming, et al. Formation of biogenic amines and growth of spoilage-related microorganisms in pork stored under different packaging conditions applying PCA[J]. Meat Science,2014, 96(2): 843-848. DOI:10.1016/j.meatsci.2013.09.023.
[22] IGNATENKO N A, BESSELSEN D G, ROY U K, et al. Dietary putrescine reduces the intestinal anticarcinogenic activity of sulindac in a murine model of familial adenomatous polyposis[J]. Nutrition &Cancer, 2006, 56(2): 172-181. DOI:10.1207/s15327914nc5602_8.
[23] FILIPPIS F D, PENNACCHIA C, PASQUA R D, et al. Decarboxylase gene expression and cadaverine and putrescine production by Serratia proteamaculans in vitro and in beef[J]. International Journal of Food Microbiology, 2013, 165(3): 332-338. DOI:10.1016/j.ijfoodmicro.2013.05.021.
[24] HALáSZ A, BARáTH á, SIMON-SARKADI L, et al. Biogenic amines and their production by microorganisms in food[J]. Trends in Food Science and Technology, 1994, 5(94): 42-49. DOI:10.1016/0924-2244(94)90070-1.
[25] LADERO V, CALLES-ENRíQUEZ M, FERNáNDEZ M,et al. Toxicological effects of dietary biogenic amines[J].Current Nutrition & Food Science, 2010, 6(2): 145-156.DOI:10.2174/157340110791233256.
[26] ZHANG Qilin, LIN Shenglin, NIE Xiaohua. Reduction of biogenic amine accumulation in silver carp sausage by an amine-negative Lactobacillus plantarum[J]. Food Control, 2013, 32(2): 496-500.DOI:10.1016/j.foodcont.2013.01.029.
[27] 朱志遠. 發酵劑對發酵香腸生物胺含量及理化品質的影響研究[D].南京: 南京農業大學, 2009: 18-20. DOI:10.7666/d.Y1762735.
[28] W?HRL S, HEMMER W, FOCKE M, et al. Histamine intolerancelike symptoms in healthy volunteers after oral provocation with liquid histamine[J]. Allergy and Asthma Proceedings, 2004, 25(5): 305-311.[29] RAUSCHER-GABERNIG E, GROSSGUT R, BAUER F, et al.Assessment of alimentary histamine exposure of consumers in Austria and development of tolerable levels in typical foods[J]. Food Control,2009, 20(4): 423-429. DOI:10.1016/j.foodcont.2008.07.011.
[30] VIDAL-CAROU M C, IZQUIERDO-PULIDO M L, MARTíNMORRO M C, et al. Histamine and tyramine in meat products:relationship with meat spoilage[J]. Food Chemistry, 1990, 37(4): 239-249. DOI:10.1016/0308-8146(90)90104-C.
[31] PIRCHER A, BAUER F, PAULSEN P. Formation of cadaverine,histamine, putrescine and tyramine by bacteria isolated from meat,fermented sausages and cheeses[J]. European Food Research &Technology, 2007, 226(1/2): 225-231. DOI:10.1007/s00217-006-0530-7.
[32] LINARES D M, DEL RíO B, LADERO V, et al. Factors influencing biogenic amines accumulation in dairy products[J]. Frontiers in Microbiology, 2012, 3: 180. DOI:10.3389/fmicb.2012.00180.
[33] PARENTE E, MARTUSCELLI M, GARDINI F, et al. Evolution of microbial populations and biogenic amine production in drysausages produced in Southern Italy[J]. Journal of Applied Microbiology, 2001,90(6): 882-891. DOI:10.1016/0008-6215(96)00070-5.
[34] KURT S, ZORBA O. Biogenic amine formation in Turkish dry fermented sausage (sucuk) as affected by nisin and nitrite[J]. Journal of the Science of Food & Agriculture, 2010, 90(15): 2669-2674.DOI:10.1002/jsfa.4138.
[35] MONTEL M C, MASSON F, TALON R. Comparison of biogenic amine content in traditional and industrial French dry sausages[J].Sciences des Aliments, 1999, 19(2): 247-254.
[36] BOVER-CID S, IZQUIERDO-PULIDO M, VIDAL-CAROU M C.Mixed starter cultures to control biogenic amine production in dry fermented sausages[J]. Journal of Food Protection, 2000, 63(11):1556-1562. DOI:10.4315/0362-028X-63.11.1556.
