基于線粒體細(xì)胞色素b基因序列探討長江流域斑點(diǎn)蛇種群遺傳結(jié)構(gòu)和地理分化
2018-05-28 04:27:45李小兵唐瓊英俞丹劉煥章
四川動物 2018年3期
李小兵, 唐瓊英, 俞丹, 劉煥章*
(1. 中國科學(xué)院水生生物研究所,水生生物多樣性與保護(hù)重點(diǎn)實(shí)驗(yàn)室,武漢430072; 2. 中國科學(xué)院大學(xué),北京100049)
長江是我國第一大河流,跨越我國大陸地勢的三大階梯,地貌類型多樣,氣候差異顯著,形成了多樣的生境,呈現(xiàn)出豐富的魚類多樣性及較高程度的種群分化(Chengetal. ,2015a,2015b)。如,基于線粒體細(xì)胞色素b(Cytb)基因的研究表明,長江流域的寬鰭Zaccoplatypus包含4個獨(dú)立進(jìn)化的線粒體譜系,且譜系間存在較大的遺傳分化,可能對應(yīng)4個不同物種(Perdicesetal. ,2004)。類似的研究結(jié)果在銅魚Coreiusheterodon、圓口銅魚C.guichenoti、長薄鰍Leptobotiaelongata和中華倒刺鲃Spinibarbussinensis中也有報道(袁希平等,2008;田輝伍等,2013;任瀧等,2014)。這些研究充分體現(xiàn)了長江流域魚類豐富的種群遺傳多樣性(Zhangetal. ,2015)。
線粒體DNA(mtDNA)具有分子結(jié)構(gòu)簡單、嚴(yán)格的母系遺傳、不同區(qū)域進(jìn)化速度存在差異等特點(diǎn),已被廣泛應(yīng)用于系統(tǒng)發(fā)育及種群遺傳研究(Patwardhanetal. ,2014;Lakshmananetal. ,2015)。其中,Cytb基因由于進(jìn)化速度適中,且易擴(kuò)增,已被廣泛用于種群遺傳多樣性的研究(Chiuetal. ,2015)。本文擬通過對采自長江上游赤水河江段、長江上游干流江段(包括合江江段和宜賓江段)及長江中游江段(洞庭湖、贛江及鄱陽湖)的斑點(diǎn)蛇種群,進(jìn)行Cytb基因的序列測定,分析不同地理種群的遺傳結(jié)構(gòu),探討其分化過程,以期為長江魚類資源的保護(hù)提供科學(xué)的理論依據(jù)。
1 材料與方法
1.1 材料
本研究所用樣本于2010—2015年采集于長江流域20個樣點(diǎn),共計303尾。根據(jù)地理分布,將其分為3個地理種群。來自長江上游赤水河江段10個樣點(diǎn)的為赤水河種群,四川省宜賓市和合江縣2個樣點(diǎn)屬于長江上游干流種群,其余8個樣點(diǎn)分別來自湖南洞庭湖水系、江西贛江及鄱陽湖水系,屬于長江中游種群。標(biāo)本均用95%乙醇固定保存于中國科學(xué)院水生生物研究所淡水魚類博物館(圖1,表1)。
1.2 方法
1.2.1基因組DNA的提取、PCR擴(kuò)增及測序基因組DNA的提取采用高鹽抽提法。基因序列片段通過PCR擴(kuò)增所得。擴(kuò)增所用引物為L14724和H15915。PCR反應(yīng)的總體積為60 μL,約50 ng基因組DNA為模板,反應(yīng)體系包括:10×Buffer 6 μL,dNTPs 0.75 μL(10 mmol·L-1),引物各1.5 μL(10 μmol·L-1),Taq DNA聚合酶1.5 U,加滅菌雙蒸水至60 μL。PCR反應(yīng)條件為:94 ℃預(yù)變性3 min;94 ℃變性45 s,52~58 ℃退火45 s,72 ℃延伸1 min,共35個循環(huán);最后72 ℃延伸8 min。將獲得的PCR產(chǎn)物送生工生物工程(上海)股份有限公司完成測序。
1.2.2數(shù)據(jù)分析為確保序列準(zhǔn)確性,對獲得的序列,通過DNASTAR中的Seqman手動拼接。用Clustal X 2.1進(jìn)行序列比對(Larkinetal. ,2007)。為避免序列中存在測序誤讀等錯誤,采用MEGA6.0將其翻譯成相應(yīng)的氨基酸序列進(jìn)行校正(Tamuraetal. ,2013)。使用PAUP* 4.0b10(Swofford,2002)和MEGA6.0對序列進(jìn)行初步分析,包括堿基組成及核苷酸同質(zhì)性檢驗(yàn)等。
采用DnaSP 5.10.1統(tǒng)計種群遺傳多樣性參數(shù),并獲得單倍型序列數(shù)據(jù)組。基于單倍型序列數(shù)據(jù),采用鄰接法(NJ)、最大似然法(ML)和貝葉斯法(BI)構(gòu)建分子系統(tǒng)發(fā)育樹。使用MrMTgui、Modeltest 3.7和PAUP* 4.0b10選擇最適合的堿基替代模型,對數(shù)據(jù)的擬合度通過AIC來檢測。NJ分析在PAUP* 4.0b10中進(jìn)行,系統(tǒng)樹的分支置信度采用自引導(dǎo)法進(jìn)行1 000次重復(fù)檢測。BI分析用MrBayes 3.2.5,以后驗(yàn)概率來表示各分支的可信性(Ronquist & Huelsenbeck,2003)。ML分析用PhyML 3.0,通過1 000次自引導(dǎo)法檢驗(yàn)各分支的置信度(Guindon &

表1 長江流域斑點(diǎn)蛇采樣點(diǎn)地理信息、樣本數(shù)量及單倍型信息Table 1 Sample location, size and haplotype information of Saurogobio punctatus in the Yangtze River basin
1.貴州省敘永縣赤水鎮(zhèn), 2. 貴州省大方縣田坎鄉(xiāng), 3.貴州省仁懷市茅臺鎮(zhèn), 4. 貴州省赤水市葫市鎮(zhèn), 5. 貴州省赤水市復(fù)興鎮(zhèn), 6. 貴州省赤水市大同鎮(zhèn), 7. 貴州省赤水市, 8. 四川省合江縣先市鎮(zhèn), 9. 四川省合江縣密溪鄉(xiāng), 10. 貴州省赤水市長沙鎮(zhèn), 11. 四川省合江縣, 12. 四川省宜賓市, 13. 湖南省岳陽市, 14. 江西省武寧縣, 15. 江西省上饒市, 16. 江西省新余市, 17. 江西省都昌縣, 18. 江西省鄱陽縣, 19.江西省鷹潭市, 20. 江西省婺源縣
1. Chishui town, Xuyong county, Guizhou province, 2. Tiankan town, Dafang county, Guizhou province, 3. Maotai town, Renhuai city, Guizhou province, 4. Hushi town, Chishui city, Guizhou province, 5. Fuxing town, Chishui city, Guizhou province, 6. Datong town, Chishui city, Guizhou province, 7. Chishui city, Guizhou province, 8. Xianshi town, Hejiang county, Sichuan province, 9. Mixi town, Hejiang county, Sichuan province, 10. Changsha town, Chishui city, Sichuan province, 11. Hejiang county, Sichuan province, 12. Yibin city, Sichuan province, 13. Yueyang city, Hunan province, 14. Wuning county, Jiangxi province, 15. Shangrao city, Jiangxi province, 16. Xinyu city, Jiangxi province, 17. Duchang county, Jiangxi province, 18. Poyang county, Jiangxi province, 19. Yingtan city, Jiangxi province, 20. Wuyuan county, Jiangxi province
Gascuel,2003)。利用Network 4.613中的中接法構(gòu)建單倍型的網(wǎng)絡(luò)關(guān)系圖,分析單倍型之間的進(jìn)化關(guān)系。
使用Arlequin 3.5進(jìn)行分子方差分析(AMOVA),計算遺傳分化指數(shù)(FST)、錯配分布值及Tajima’sD和 Fu’sFs值(Excoffier & Lischer,2010)。使用BEAST 1.8.2進(jìn)行種群動態(tài)的Bayesian skyline plot(BSP)分析(Drummondetal. ,2012)。
2 結(jié)果
2.1 Cyt b基因的序列變異分析
2.2 單倍型系統(tǒng)發(fā)育關(guān)系及進(jìn)化網(wǎng)絡(luò)分析
單倍型進(jìn)化網(wǎng)絡(luò)關(guān)系圖(圖3)顯示:整體上,單倍型間的網(wǎng)絡(luò)進(jìn)化關(guān)系呈星狀分布,單倍型間彼此相連、相差一步或多步的突變距離。單倍型Hap3(為長江上游干流種群和赤水河種群共享)為較原始的單倍型,位于星狀網(wǎng)絡(luò)圖最中心的位置。其他單倍型均由Hap3直接演化或逐步演化而來。其中,經(jīng)過Hap3多次演化后的Hap11(為長江上游干流種群和赤水河種群共享)又演化出更多的單倍型。
2.3 不同地理種群的遺傳多樣性、遺傳分化及分子方差分析
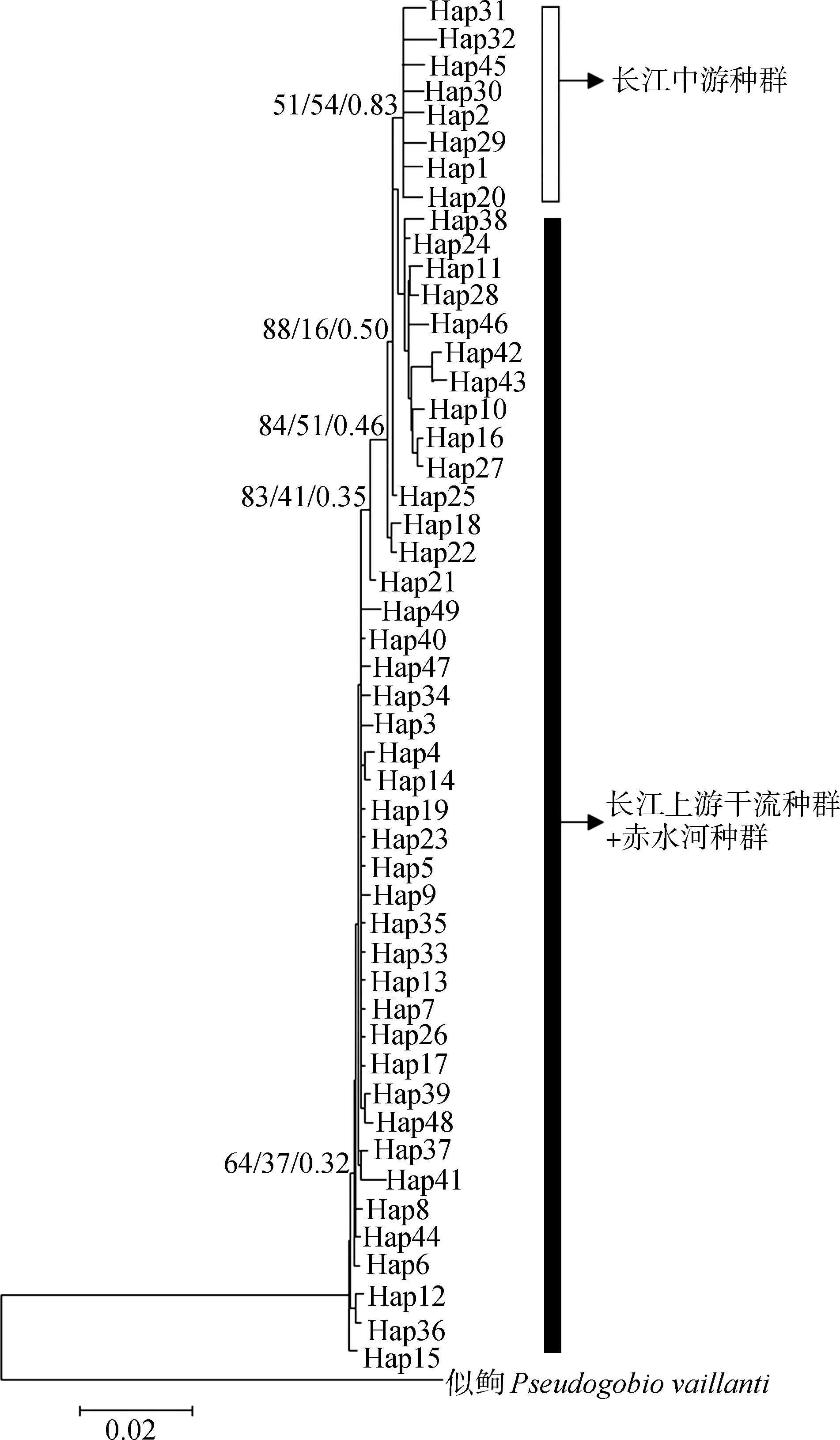
圖2 基于線粒體Cyt b基因單倍型構(gòu)建的長江流域 斑點(diǎn)蛇不同地理種群的鄰接樹、最大似然樹、貝葉斯樹Fig. 2 Neighbor-Joining/Maximum-Likelihood/Bayesian inference tree of Saurogobio punctatus populations based on mtDNA Cyt b gene in the Yangtze River basin
節(jié)點(diǎn)處的數(shù)值代表NJ/ML/BI分析獲得的支持率, 支長代表分歧度
Values at the node represent support values in NJ/ML/BI analysis, respectively; the length of branch indicates the percentage of divergence

圖3 基于線粒體Cytb基因的長江流域斑點(diǎn)蛇不同地理種群的單倍型網(wǎng)絡(luò)圖
Fig. 3 The haplotype network ofSaurogobiopunctatusbased onCytbgene in the Yangtze River basin
灰色部分、黑色部分和白色部分分別表示赤水河種群、長江上游干流種群及長江中游種群所擁有的單倍型; 圓圈的大小表示單倍型包含的個體數(shù); 中間小黑點(diǎn)代表缺失的單倍型
Gray, black and white portions represent the Chishui River population, the upper Yangtze River main stream population, and the middle Yangtze River population, respectively; black dot represents the lacked haplotype
表2 長江流域斑點(diǎn)蛇不同地理種群的遺傳多樣性統(tǒng)計參數(shù)Table 2 Genetic diversity parameters of Saurogobio punctatus in different populations of the Yangtze River basin
注Notes:*P<0.05,**P<0.01; 下同the same below
表3 長江流域斑點(diǎn)蛇不同地理種群的遺傳分化指數(shù)Table 3 Genetic differentiation index among differentgeographic populations of Saurogobio punctatusin the Yangtze River basin
表4 基于線粒體Cyt b基因?qū)﹂L江流域斑點(diǎn)蛇的分子方差分析Table 4 AMOVA analysis based on Cyt b of Saurogobio punctatus among 3 populations in the Yangtze River basin
2.4 長江流域斑點(diǎn)蛇種群的動態(tài)分析
3 討論
3.1 長江流域斑點(diǎn)蛇不同地理種群的遺傳分化與變異
由于地理隔離,同一物種的不同地理種群可能會產(chǎn)生遺傳分化(Brownetal. ,1979)。本研究結(jié)果顯示,長江上游干流和赤水河斑點(diǎn)蛇種群間的FST值較低,為0.029 4。而長江中游與長江上游干流和赤水河種群的FST值均較高,且差異有高度統(tǒng)計學(xué)意義。Wright(1978)認(rèn)為,若種群的FST值為0~0.05,說明不存在分化;0.05~0.15為中度分化;0.15~0.25為高度分化。因此,斑點(diǎn)蛇長江上游干流種群與赤水河種群間未發(fā)生遺傳分化或分化程度較低,而長江中游種群與上游種群則已發(fā)生高度分化。長江上游不同江段同種種群間不存在遺傳分化的現(xiàn)象在其他魚類中也有報道,如,田輝伍等(2013)發(fā)現(xiàn)長江干流與赤水河、岷江及嘉陵江的長薄鰍種群間基因交流頻繁。但長江中游與上游的斑點(diǎn)蛇間遺傳分化顯著,種群間缺乏廣泛的交流,這與長江流域Hemiculterleucisculus的研究結(jié)果一致(范啟,何舜平,2014)。

圖4 長江流域斑點(diǎn)蛇種群動態(tài)隨時間變化 的Bayesian skyline plots
Fig. 4 Bayesian skyline plots for population dynamics with time ofSaurogobiopunctatusin the Yangtze River basin
(a) 長江流域種群, (b) 長江上游干流種群, (c) 長江中游種群; 黑色線表示中值, 陰影部分為95%的置信區(qū)間
(a) the population from the Yangtze River basin, (b) the population from the upper Yangtze River main stream, (c) the population from the middle Yangtze River; the black line indicates the median, the shadow part shows the 95% highest probability density
3.2 長江流域斑點(diǎn)蛇不同地理種群的遺傳多樣性
作為生物多樣性的重要組成部分,遺傳多樣性是經(jīng)歷突變及長時間自然選擇后產(chǎn)生的結(jié)果,也是一個物種生存適應(yīng)和發(fā)展進(jìn)化的前提(Taoetal. ,2013)。其中,Hd和Pi是衡量種群遺傳變異程度的2個重要指標(biāo)(Brooksetal. ,2015)。Hd和Pi較高說明種群遺傳多樣性高,遺傳資源豐富(Chen & He,2002)。在長江流域斑點(diǎn)蛇3個種群中共檢測到49個單倍型,整體的Hd和Pi平均值分別為0.803和0.003 71,呈現(xiàn)較高的單倍型多樣性和較低的核苷酸多樣性,暗示該種群可能經(jīng)歷過瓶頸效應(yīng)或奠基者效應(yīng)后迅速擴(kuò)張(Goodall-Copestakeetal. ,2012;Yangetal. ,2014)。在擴(kuò)張后的群體中,遺傳變異雖導(dǎo)致了單倍型多樣性的增加,但核苷酸多樣性依然維持在相對較低的水平(Arundelletal. ,2015)。
3.3 長江流域斑點(diǎn)蛇的種群歷史
在種群動態(tài)分析中,Tajima’sD值和 Fu’sFs值是2種通過中性檢驗(yàn)研究種群在歷史上是否發(fā)生過擴(kuò)張的較常用參數(shù)。Tajima’sD值和Fu’sFs值接近0時,表明種群處于穩(wěn)定狀態(tài),兩者為負(fù)且P值小于0.05被認(rèn)為種群在歷史上有擴(kuò)張跡象。此外,對于發(fā)生過擴(kuò)張的種群,錯配分布圖一般呈現(xiàn)單峰(Ramos-Onsins & Rozas,2002)。本研究中,中性檢驗(yàn)及錯配分布圖均表明,整體上,長江流域斑點(diǎn)蛇經(jīng)歷過種群的快速擴(kuò)張;長江上游干流種群和赤水河種群曾發(fā)生過擴(kuò)張現(xiàn)象,而長江中游種群則未能檢測到種群擴(kuò)張,處于穩(wěn)定狀態(tài)。此外,斑點(diǎn)蛇種群動態(tài)隨時間變化的BSP圖顯示,整體上,長江流域斑點(diǎn)蛇從0.20 Ma BP開始發(fā)生種群擴(kuò)張,一直持續(xù)到末次間冰期(MIS5)晚期(75~128 ka BP)(Thieletal. ,2015);而后種群得到迅速擴(kuò)張,一直持續(xù)到0.05 Ma BP左右;此后,種群保持相對穩(wěn)定。另外,據(jù)單倍型網(wǎng)絡(luò)圖,Hap3(長江上游干流和赤水河種群共享的單倍型)是最原始的單倍型,而長江中游種群的單倍型均由其間接演化而來。因此推測,斑點(diǎn)蛇的起源中心可能在長江上游,然后通過種群擴(kuò)張逐漸擴(kuò)散到長江中游,進(jìn)化成遺傳分化較大的種群。
致謝:感謝課題組劉飛助理研究員、但勝國高級實(shí)驗(yàn)師及張富斌等同學(xué)在采樣方面給予的幫助。貴州省赤水市畜牧水產(chǎn)局的婁必云、黎良等同志在采樣方面提供諸多便利,一并致以謝忱!
:
陳宜瑜. 1998. 中國動物志 硬骨魚綱 鯉形目(中卷)[M]. 北京: 科學(xué)出版社: 379-389.
丁淑荃, 萬全, 李飛, 等. 2013. 斑點(diǎn)蛇精子結(jié)構(gòu)研究[J]. 安徽農(nóng)業(yè)大學(xué)學(xué)報, 40(6): 959-963.
范啟, 何舜平. 2014. 長江流域種群遺傳多樣性和遺傳結(jié)構(gòu)分析[J]. 水生生物學(xué)報, 38(4): 627-635.
任瀧, 吳金明, 李雷, 等. 2014. 赤水河中華倒刺鲃的形態(tài)分化及Cytb基因序列比較研究[J]. 淡水漁業(yè), 44(6): 58-64.
田輝伍, 段辛斌, 汪登強(qiáng), 等. 2013. 長江上游長薄鰍Cytb基因的序列變異與遺傳結(jié)構(gòu)分析[J]. 淡水漁業(yè), 43(6): 12-18.
袁希平, 嚴(yán)莉, 徐樹英, 等. 2008. 長江流域銅魚和圓口銅魚的遺傳多樣性[J]. 中國水產(chǎn)科學(xué), 15(3): 377-385.
Arundell K, Dunn A, Alexander J,etal. 2015. Enemy release and genetic founder effects in invasive killer shrimp populations of Great Britain[J]. Biological Invasions, 17(5): 1439-1451.
Brooks TM, Cuttelod A, Faith DP,etal. 2015. Why and how might genetic and phylogenetic diversity be reflected in the identification of key biodiversity areas?[J]. Philosophical Transactions of the Royal Society B, 370: 20140019. DOI: 10.1098/rstb.2014.0019.
Brown WM, George M, Wilson AC. 1979. Rapid evolution of animal mitochondrial DNA[J]. Proceedings of the National Academy of Sciences of USA, 176(4): 1967-1971.
Chen YF, He DK. 2002. Biodiversity in the Yangtze River basin fauna and distribution of fishes[J].Journal of Ichthyology, 42: 161-171.
Cheng F, Li W, Castello L,etal. 2015a. Potential effects of dam cascade on fish: lessons from the Yangtze River[J]. Reviews in Fish Biology and Fisheries, 3: 569-585.
Cheng F, Li W, Klopfer M,etal. 2015b. Population genetic structure and its implication for conservation ofCoreiusguichenotiin the upper Yangtze River[J]. Environmental Biology of Fishes, 98(9): 1999-2007.
Chiu TH, Kuo CW, Lin HC,etal. 2015. Genetic diversity of ivory shell (Babyloniaareolata) in Taiwan and identification of species using DNA-based assays[J]. Food Control, 48: 108-116.
Drummond AJ, Suchard MA, Xie D,etal. 2012. Bayesian phylogenetics with BEAUti and the BEAST 1.7[J]. Molecular Biology and Evolution, 29: 1969-1973.
Excoffier L, Lischer HEL. 2010. Arlequin suite ver 3.5: a new series of programs to perform population genetics analyses under Linux and Windows[J]. Molecular Ecology Resources, 10: 564-567.
Goodall-Copestake WP, Tarling GA, Murphy EJ. 2012. On the comparison of population-level estimates of haplotype and nucleotide diversity: a case study using the gene cox1 in animals[J]. Heredity, 109: 50-56.
Guindon S, Gascuel O. 2003. A simple, fast and accurate method to estimate large phylogenies by maximum-likelihood[J]. Systematic Biology, 52: 696-704.
Lakshmanan LK, Gruber J, Halliwell B,etal. 2015. Are mutagenic non D-loop direct repeat motifs in mitochondrial DNA under a negative selection pressure?[J]. Nucleic Acids Research, 43(8): 4098-4108.
Larkin MA, Blackshields G, Brown NP,etal. 2007. Clustal W and clustal X version 2.0[J]. Bioinformatics, 23: 2947-2948.
Patwardhan A, Ray S, Roy A. 2014. Molecular markers in phylogenetic studies-a review[J]. Phylogenetic and Evolutionary Biology, 2: 131.
Perdices A, Cunhaa C, Coelhoa MM. 2004. Phylogenetic structure ofZaccoplatypus(Teleostei, Cyprinidae) populations on the upper and middle Chang Jiang (=Yangtze) drainage inferred from cytochrome b sequences[J]. Molecular Phylogenetics and Evolution, 31(1): 192-203.
Ramos-Onsins SE, Rozas J. 2002. Statistical properties of new neutrality tests against population growth[J]. Molecular Biology and Evolution, 19(12): 2092-2100.
Ronquist F, Huelsenbeck JP. 2003. MrBayes 3: Bayesian phylogenetic inference under mixed models[J]. Bioinformatics, 19: 1572-1574.
Swofford DL. 2002. PAUP*: phylogenetic analysis using parsimony (* and other methods), version 4[M]. Sunderland, Massachusetts: Sinauer Associates.
Tamura KG, Stecher D, Peterson AF,etal. 2013. MEGA6: molecular evolutionary genetics analysis version 6.0[J]. Molecular Biology and Evolution, 30: 2725-2729.
Tang QY, Li XB, Yu D,etal. 2018.Saurogobiopunctatussp. nov. , a new cyprinid gudgen (Teleostei: Cypriniformes) from the Yangtze River, based on both morphological and molecular data[J]. Journal of Fish Biology, 92(2): 347-364.
Tao W, Jin XX, Xu TJ. 2013. The first complete mitochondrial genome fromBostrychusgenus (Bostrychussinensis) and partitioned Bayesian analysis of Eleotridae fish phylogeny[J]. Journal of Genetics, 92(2): 247-257.
Thiel C, Tsukamoto S, Tokuyasud K,etal. 2015. Testing the application of quartz and feldspar luminescence dating to MIS 5 Japanese marine deposits[J]. Quaternary Geochronology, 29: 16-29.
Wright S. 1978. Evolution and the genetics of populations volume 4: variability within and among natural populations[M]. Chicago: University of Chicago Press.
Yang M, Tian C, Liang XF,etal. 2014.Genetic structure and diversity in natural and stocked populations of the mandarin fish (Sinipercachuatsi) in China[J]. Genetics and Molecular Research, 14(2): 5153-5160.
Zhang X, Gao X, Wang JW,etal. 2015. Extinction risk and conservation priority analyses for 64 endemic fishes in the upper Yangtze River, China[J]. Environmental Biology of Fishes, 98(1): 261-272.
