內蒙古賽罕烏拉國家級自然保護區中華斑羚的季節性食物構成分析
2018-05-28 04:27:58唐書培姜秀麗王曉玲洪美靜孟和達來陳琛韓瑩瑩鮑偉東
四川動物 2018年3期
關鍵詞:植物
唐書培, 姜秀麗, 王曉玲, 洪美靜, 孟和達來, 陳琛, 韓瑩瑩, 鮑偉東*
(1.北京林業大學生物科學與技術學院,北京100083; 2. 內蒙古賽罕烏拉國家級自然保護區管理局,內蒙古大板025150)
食物是聯系動物與環境的紐帶,食性研究是動物生態學中的重要內容,可以直接反映野生動物對棲息地環境的適應程度,也能反映其與同域生存的其他物種的生態關系(Andrews,2003;郭相保等,2011;姚志誠等,2011;劉燕等,2015)。在實踐中,明確野生動物的食物需求、適應和選擇性,有利于開展物種的保護管理(朱世兵,2015;Giottoetal. ,2016),尤其對瀕危珍稀野生動物的保護與管理、馴養與繁殖均具有重要意義(單繼紅,吳建平,2005;司曉艷等,2007)。
有關動物食性研究的方法較多,目前國內外常見的食草動物食性研究方法主要有胃分析法、籠養飼喂法、糞便顯微分析法、直接觀察法和利用法以及糞樣內容物高通量測序技術(高中信等,1991;鄭榮泉,鮑毅新,2004;劉剛等,2018),其中,糞便顯微分析法具有對動物干擾較小、操作簡單、實驗成本低等優點,被國內外學者廣泛使用(昶野,2010;李邦等,2014;Rduch,2016)。張春楊等(2011)采用模擬動物野外采食及糞便顯微分析法相結合的方式研究了圈養梅花鹿Cervusnippon的食性,找出了糞便中組織碎片與其采食量之間的關系,表明糞便顯微分析法可以準確地分析梅花鹿的食物組成比例。
雖然國內外對于瀕危野生動物的食性開展了廣泛的研究,但是瀕危野生動物的棲息地往往交通閉塞、環境惡劣,導致野外調查、樣品采集工作難以順利開展,因此,目前的研究大多局限于對圈養條件下瀕危動物的食性分析,或是局限于某個物種在某一區域、單一季節的食性研究,缺乏對某個種群長期的、涵蓋各個季節的食性研究(Ilyas & Khan,2003;Rduch,2016)。中華斑羚Naemorhedusgriseus主要分布于我國內蒙古、河北、北京、河南、陜西、山西、甘肅、寧夏、四川、貴州、重慶、湖北、湖南、廣西、廣東、江西、福建、浙江、上海、江蘇、安徽和印度、緬甸、泰國、越南,被列入《瀕危野生動植物種國際貿易公約(CITES)》附錄Ⅰ,在世界自然保護聯盟(IUCN)瀕危物種紅色名錄和《中國脊椎動物紅色名錄》中被列為瀕危(VU)物種(蔣志剛等,2015,2016)。本研究著眼于內蒙古賽罕烏拉國家級自然保護區內中華斑羚種群,收集了該區域中華斑羚2015年12月—2016年9月各季節的糞便樣品。利用糞便顯微分析法對該地區中華斑羚的食性進行分析,以期能獲取全面、精準的食性數據,為后期評估其生境選擇、環境容納量、探討生存適應等保護生物學問題提供理論基礎。
1 材料和方法
1.1 研究區域
賽罕烏拉國家級自然保護區位于內蒙古自治區赤峰市巴林右旗北部(118°18′~118°55′E,43°59′~44°27′N),面積10.04×104hm2,屬于大興安嶺山地南部典型森林生態系統(圖1)。保護區劃分為正溝、王汶溝和烏蘭壩3個核心區,現有的中華斑羚只分布于正溝核心區內,該區由3條山脊大體圍成“Ψ”狀,涵蓋闊葉林、針闊混交林、針葉林、灌木以及草甸等植被類型,主要包括白樺Betulaplatyphylla、蒙古櫟Quercusmongolica、華北落葉松Larixprincipis-rupprechtii、山楊Populusdavidiana等喬木,虎榛子Ostryopsisdavidiana、胡枝子Lespedezabicolor、興安杜鵑Rhododendrondauricum、山杏Armeniacasibirica等灌木,以及華蟲實Corispermumstauntonii、華北剪股穎Agrostisclavata、柄狀薹草Carexpediformis等草本(雍世鵬等,2011)。
1.2 研究方法
1.2.1糞便樣品及植物樣品的采集利用非損傷性取樣方法在保護區的正溝核心區內收集中華斑羚糞便樣品。沿中華斑羚足跡收集新鮮糞便(顏色較深,表面濕潤光澤)于一次性自封袋中,記錄收集時間、地理坐標、生境類型等數據,-20 ℃保存備用。根據當地氣候條件,四季時間為春季3—5月、夏季6—8月、秋季9—10月、冬季11月—翌年2月(王安夢,2010)。
收集糞便樣品的同時,在中華斑羚棲息地內隨機設置10個10 m×10 m樣方,依次采集并記錄各樣方中的植物種類,放在大信封內,并在信封上標明編號、采集日期、生境類型等信息。采用恩格勒植物分類系統,對照本區域植物名錄(雍世鵬等,2011)鑒定物種,并將采集的植物樣品劃分為喬木、灌木、禾本科Poaceae草本和非禾本科草本,制備標準植物表皮細胞圖片,用于對照鑒定中華斑羚糞樣中植物種類。
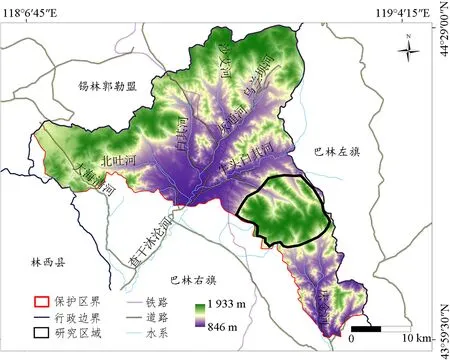
圖1 研究區域Fig. 1 Survey area
1.2.2顯微片制備中華斑羚糞便樣品進行2次抽樣。按照不同季節劃分,將每個采樣點的糞便樣品分別取一些糞粒組成混合糞便樣本,經烘干、研缽研碎;取過篩的植物碎片樣品于濃硝酸中消化后洗凈,用0.1%甲苯胺藍水溶液染色,再制片(李邦等,2014)。
1.2.3鏡檢及統計分析方法每種植物和每份中華斑羚糞便混合樣本均制作3張玻片。將玻片在放大100倍顯微鏡下鏡檢,每張玻片按照從上到下、從左到右的順序隨機選取10個視野,記錄出現的特征性植物表皮角質碎片,拍照留存。同時,將鏡檢糞便混合樣本可辨認的植物表皮角質碎片與標準植物表皮結構照片對比,統計每種植物出現在糞便樣品中的相對密度(RD),RD=每種植物的碎片數/各種植物的碎片總數×100%,代表中華斑羚食物中某植物的組成比例。
鏡檢時,植物表皮細胞的可鑒別特征包括:細胞的形態、大小、密度、排列方式等;表皮毛基細胞的大小、形狀、密度等;氣孔器的形狀、大小、方向等,以及氣孔與表皮毛基細胞的相對大小等(Johnson,1982;高中信等,1991;昶野,2010;蘇軍虎等,2017)。
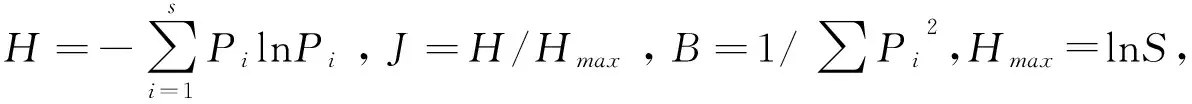
使用單樣本Kolmogorov-Smirnov檢驗各季節中華斑羚取食食物種類和同一食物組成(包括喬木、灌木、禾本科草本、非禾本科草本)中不同季節植物種類的正態分布情況,發現變量不呈正態分布。因此,利用非參數的Kruskal-WallisH檢驗分析變量的差異性,顯著水平設為0.05;Mann-WhitneyU檢驗分析2個季節間食物種類構成的差異,經Bonferroni校正后的顯著水平為0.008 3。數據分析均在SPSS 24.0中完成。
2 研究結果
2.1 中華斑羚各季節的食物組成和比例
全年共收集到中華斑羚糞便樣品554份,其中春季181份,夏季84份,秋季78份,冬季211份。由于中華斑羚生活于山頂開闊地帶,警惕性很高,毛色與環境的相似度高,且容易發現人類干擾,很難直接觀察,因此,本研究未進行中華斑羚取食行為觀察。樣方法共鑒定出植物18科35屬41種。
糞便混合樣本顯微照片與標準植物表皮結構照片比對結果表明,中華斑羚全年共采食植物16科27屬31種。其中,春季取食14科25種,華蟲實、虎榛子、華北剪股穎為主要食物,RD值分別為27.97%、27.31%和13.00%。取食植物組成中,非禾本科草本占比最高,為41.03%,其次為灌木,占37.21%,禾本科草本占14.36%,喬木占7.40%。
中華斑羚夏季取食植物16科29種,其中,華北剪股穎(16.16%)、柄狀薹草(11.26%)、胡枝子(7.27%)、華蟲實(6.65%)、鵝觀草Roegneriakamoji(6.24%)、蒙古櫟(6.13%)、地榆Sanguisorbaofficinalis(5.83%)、虎榛子(5.68%)、興安杜鵑(5.54%)等占夏季食物總量的70.76%。取食植物組成中,非禾本科草本占比最高,為40.43%,其次為禾本科草本,占24.98%,灌木占23.53%,喬木占11.06%。
中華斑羚秋季取食植物13科24種,其中,虎榛子(18.26%)、華北剪股穎(17.35%)、柄狀薹草(7.27%)、華蟲實(7.17%)、線葉菊Filifoliumsibiricum(6.77%)、地榆(6.43%)、蒙古櫟(5.91%)等占秋季食物總量的69.16%。取食植物組成中,非禾本科草本占比最高,為36.34%,其次為灌木,占30.47%,禾本科草本占25.41%,喬木占7.78%。
中華斑羚冬季取食植物8科11種,虎榛子、華北剪股穎、柄狀薹草、線葉菊、華蟲實組成了中華斑羚冬季的主要食物,其RD值分別為44.78%、23.88%、8.96%、5.97%和5.97%。取食植物組成中,灌木占比最高,為48.41%,其次為禾本科草本,占25.16%,非禾本科草本占22.23%,喬木僅有白樺1種,占4.20%。
2.2 中華斑羚食物的季節性差異
非參數Kruskal-WallisH檢驗結果顯示,不同季節中華斑羚食物種類構成的差異有高度統計學意義(χ2=28.865,df=3,P<0.001)。Mann-WhitneyU檢驗結果顯示,冬季與其他季節間食物種類的差異均有高度統計學意義(冬季與春季:Z=-3.574,P<0.001;冬季與夏季:Z=-4.739,P<0.001;冬季與秋季:Z=-3.303,P=0.001),其他季節之間的差異無統計學意義(春季與夏季:Z=-1.503,P=0.133;春季與秋季:Z=-0.309,P=0.757;夏季與秋季:Z=-1.788,P=0.074)。
中華斑羚全年取食植物組成中,喬木(7.61%)的比例最低,且每個季節取食喬木均最少;非禾本科草本最多(35.00%),其次為灌木(34.91%)、禾本科草本(22.48%)(圖2)。非參數Kruskal-WallisH檢驗結果顯示,不同季節中華斑羚取食的非禾本科草本種類間的差異有高度統計學意義(χ2=17.213,df=3,P<0.001);同時Mann-WhitneyU檢驗結果顯示,中華斑羚取食的非禾本科草本種類在冬季與夏季、冬季與秋季間的差異有高度統計學意義(Z=-3.824,P<0.001;Z=-2.708,P=0.007),其他季節間的差異無統計學意義(P>0.008 3);此外,非參數Kruskal-WallisH檢驗結果顯示,中華斑羚不同季節取食的喬木、灌木及禾本科草本間的差異無統計學意義。
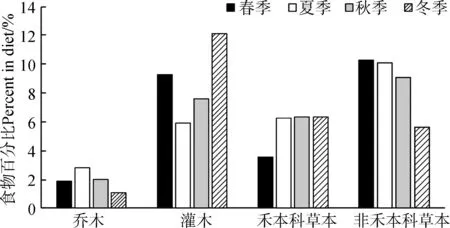
圖2 中華斑羚各季節取食植物類別的差異Fig. 2 Relative percentage of plants categories in the diets of Naemorhedus griseus during 4 seasons
各季節中華斑羚食物的Shannon-Wiener指數和食物生態位寬度由高到低依次為夏季、秋季、春季、冬季,Pielou均勻度指數由高到低依次為夏季、秋季、冬季、春季(表1)。

表1 中華斑羚各季節食物多樣性、Shannon-Wiener指數、 均勻度指數和生態位寬度指數Table 1 Food diversity, Shannon-Wiener index, evenness index and niche width index ofNaemorhedus griseus during 4 seasons
3 討論
野生動物取食策略是長期進化的結果,受環境因素、食物豐富度、自身生理結構、消化率等多方面的影響(Krebsetal. ,1995;劉丙萬,蔣志剛,2002;蔣志剛,2004;昶野,2010)。吳建平等(2005)利用糞便顯微組織學技術結合野外調查,對小興安嶺通河林區斑羚Naemorhedusgoral的冬季食性研究表明,斑羚的取食對策受環境的影響,并隨取食生境中食物的豐富程度而變化。張維等(2006)利用糞便顯微組織學技術對黑龍江龍口自然保護區斑羚的冬季食性研究表明,斑羚冬季很少取食粗蛋白和粗脂肪含量高的針葉類植物,而選擇易獲取的林下植物和落葉,這是長期進化、自然選擇的結果。
內蒙古賽罕烏拉國家級自然保護區中華斑羚的冬季取食植物種類與其他季節間的差異均有高度統計學意義,即春、夏、秋3個季節取食植物種類明顯多于冬季,而春、夏、秋間的差異無統計學意義,3個季節相似的環境條件導致其可采食到的植物種類及質量相近,不同之處在于每種植物隨物候變化決定了在季節間呈現不同的形態及營養成分含量,使中華斑羚在可采食植物的種類上差異不明顯,而具體到每種植物的采食比例出現了變化。冬季草木凋零,并且長期被大雪覆蓋,導致中華斑羚棲息地內多數低矮的植物被大雪掩埋,冬季可采食的植物種類較其他3個季節大大減少,因此,適宜高度的灌木成為中華斑羚冬季取食的大宗食物,占冬季食物組成的48.41%;同時,中華斑羚的采食策略也因采食時間而變化,這一點可從中華斑羚各季節食物多樣性差異看出,春季、夏季和秋季食物比冬季豐富,中華斑羚食物多樣性指數和生態位寬度明顯增加。春季時,植物在某個時間段迅速生長,中華斑羚取食植物種類增加但食物量不穩定,因而春季的均勻度指數最低。同樣,冬季處于漫長的冰雪天氣,食物種類單一且匱乏,以至冬季的均勻度同樣不高;而夏季和秋季的食物量穩定,各種取食植物所占比例較均勻。這些情況說明,中華斑羚的冬季取食策略以能量支出和攝入的權衡關系為主,而其他季節的取食策略還包括了植物營養質量、生理需求、適口性和喜好等因素(姚志誠等,2011)。
中華斑羚全年、各季節取食喬木的比例均最小,且冬季只取食1種(白樺),低于其他季節。這種食性特點的產生原因是多方面的,首先,受體型矮小影響,中華斑羚取食喬木的難度較高,因此,其各季節取食喬木的比例均最低。其次,除冬季外,其他季節喬木枝繁葉茂,因而具有較高營養價值的喬木嫩枝葉成為中華斑羚春、夏和秋季的食物,以致這3個季節取食喬木的種類多于冬季。同時,白樺是陰坡面積最大的森林植被(李桂林,2005),因此成為了中華斑羚全年、各季節取食的大宗食物之一。再者,自然選擇促使中華斑羚取食喬木的比例低于其他植被類別。Ilyas和Khan(2003)對印度Binsar野生動物保護區內體型相似的赤麂Muntiacusmuntjak和喜馬拉雅斑羚的食性研究發現,赤麂的食物構成以高能量的嫩葉為主,而喜馬拉雅斑羚的食物構成以能量不高的草本為主,少量取食嫩枝葉,與本研究結果相似,這可能是受自然選擇長期進化的結果。
在野外工作中發現,冬季在中、下坡位的蒙古櫟林中很少存在中華斑羚的臥跡及糞便。同時研究發現,冬季中華斑羚對蒙古櫟林下的非禾本科草本取食種類也低于其他季節,而各季節對上坡位的灌木取食種類間的差異無統計學意義。因此,本研究認為出現該現象的主要原因是環境差異。冬季冰雪覆蓋、水資源豐富,但中華斑羚可取食植物種類較少,主要為生長在上坡位的灌木植物(如虎榛子),相反,其他3個季節可供中華斑羚取食的植物種類繁多,食物不再是限制因子,中華斑羚的食物構成也發生變化,取食植物種類顯著增加。由此可見,棲息地植物的種類豐富度、營養成分以及自身季節性活動規律直接影響中華斑羚取食選擇,反映出中華斑羚的食性對棲息地植被變化具有一定的適應性。
野生動物的食性分析在國內外已有廣泛研究,研究方法趨于成熟,通過對動物的食性研究,可以更加明確各地野生動物的采食策略,評估野生動物對環境的適應狀況(Razaetal. ,2005;單繼紅,吳建平,2005)。本研究對中華斑羚全年各季節的采食習性進行了比較研究,但其中仍存在一些問題,需要后期進行深入探討。根據文獻記載,中華斑羚除采食喬木、灌木、草本植物之外,還會以少量的苔蘚、地衣等為食(雍世鵬等,2011)。這些植物的細胞結構簡單、角質層較薄、易消化,能夠為中華斑羚提供適量的營養來源,但在樣品采集及后期鏡檢中效果不佳、不易辨認,所以本研究并未涉及這些植物種類。
:
昶野. 2010. 賀蘭山巖羊食性分析[D]. 哈爾濱: 東北林業大學.
高中信, 陳化鵬, 王筱平. 1991. 糞便分析法測定植食動物食性的評價[J]. 獸類學報, 11(3): 186-193.
郭相保, 王振龍, 陳菊榮, 等. 2011. 河南太行山自然保護區獼猴冬春季食性分析[J]. 生態學雜志, 30(3): 483-488.
蔣志剛, 馬勇, 吳毅, 等. 2015. 中國哺乳動物多樣性及地理分布[M]. 北京: 科學出版社.
蔣志剛, 江建平, 王躍招, 等. 2016. 中國脊椎動物紅色名錄[J]. 生物多樣性, 24(5): 500-551.
蔣志剛. 2004. 動物行為原理與物種保護方法[M]. 北京: 科學出版社.
李邦, 林恭華, 謝久祥, 等. 2014. 托勒南山半圈養白唇鹿食性分析[J]. 四川動物, 33(6): 840-845.
李桂林. 2005. 賽罕烏拉自然保護區志[M]. 赤峰: 內蒙古科學科技出版社.
劉丙萬, 蔣志剛. 2002. 普氏原羚的采食對策[J]. 動物學報, 48(3): 309-316.
劉剛, 寧宇, 夏曉飛, 等. 2018. 高通量測序技術在野生動物食性分析中的應用與展望[J]. 生態學報, 38(9): 1-9.
劉燕, 畢俊懷, 武曉東. 2015. 內蒙古中部蒙古野驢春季食性分析[J]. 動物學雜志, 50(3): 321-328.
駱穎. 2011. 賀蘭山巖羊(Pseudoisnayaur)和馬鹿(Cervuselaphusalxaicus)的食性及生境選擇比較研究[D]. 哈爾濱: 東北林業大學.
單繼紅, 吳建平. 2005. 食草動物食性研究的主要方法及其評價[J]. 野生動物學報, 26(3): 47-49.
司曉艷, 趙天飆, 毛永強, 等. 2007. 食草動物食性研究方法的評價[J]. 醫學動物防制, 23(12): 957-959.
蘇軍虎, Weihong JI, 徐長林, 等. 2017. 甘南草原主要草食動物的食性及其生態位特征[J]. 動物學雜志, 52(3): 381-389.
王安夢. 2010. 內蒙古賽罕烏拉國家級自然保護區鳥類群落研究[D]. 北京: 北京林業大學.
吳建平, 單繼紅, 王志平. 2005. 小興安嶺通河林區斑羚冬季食性分析[J]. 動物學雜志, 40(4): 40-44.
姚志誠, 劉振生, 王兆錠, 等. 2011. 賀蘭山野化牦牛冬春季食性[J]. 生態學報, 31(3): 673-679.
雍世鵬, 邢蓮蓮, 李桂林. 2011. 賽罕烏拉國家級自然保護區生物多樣性編目[M]. 呼和浩特: 內蒙古大學出版社.
張春楊, 吳建平, 黃信勇. 2011. 梅花鹿食性分析中糞便分析法的研究[J]. 野生動物學報, 32(4): 199-202.
張維, 王文, 唐珞, 等. 2006. 黑龍江龍口自然保護區斑羚冬季食性分析[J]. 東北林業大學學報, 34(3): 32, 44.
鄭榮泉, 鮑毅新. 2004. 有蹄類食性研究方法及研究進展[J]. 生態學報, 24(7): 1532-1539.
朱世兵. 2015. 基于遷移行為、食性分析的貂熊冬季生境利用和評價[D]. 哈爾濱: 東北林業大學.
Andrews K. 2003. Activity budgets and feeding behaviour of the buton macaqueMacacabrunnecens[D]. Scotland: University of Aberdeen.
Giotto N, Picot D, Moussa E,etal. 2016. Compared diets of sympatric beira antelopes and domestic goats in the growing season[J]. European Journal of Wildlife Research, 62(6): 687-693.
Ilyas O, Khan JA. 2003. Food habits of barking deer (Muntiacusmuntjak) and goral (Naemorhedusgoral) in Binsar Wildlife Sanctuary, India[J]. Mammalia, 67(4): 521-532.
Johnson MK. 1982. Frequency sampling for microscopic analysis of botanical compositions[J]. Journal of Range Management Archives, 35(4): 541.
Krebs JR, Davies NB, Wells KD. 1995. An introduction to behavioral ecology[M]. Oxford: Blackwell Scientific Publications.
Raza FS, Okamoto M, Takemori H,etal. 2005. Manganese superoxide dismutase activity in the rat adrenal[J]. Journal of Endocrinology, 184(1): 77-84.
Rduch V. 2016. Diet of the puku antelope (Kobusvardonii) and dietary overlap with selected other bovids in Kasanka National Park, Zambia[J]. Mammal Research, 61(3): 289-297.
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
