超高壓對草魚肌原纖維蛋白結構的影響
2018-06-05 08:41:49閆春子夏文水許艷順
食品與生物技術學報 2018年4期
閆春子, 夏文水, 許艷順
(江南大學 食品學院,江蘇 無錫 214122)
草魚是重要的經濟淡水魚,其肌原纖維蛋白占主要肌肉蛋白質量的55%~65%。因此,了解肌原纖維蛋白結構變化對控制產品品質有著重要意義。
超高壓技術是一種新興的非熱加工技術手段,物料經過一定的壓力處理后,其微生物大部分被殺滅,內源酶大部分被滅活。相比加熱殺菌,超高壓處理可較大幅度地保留食品原有的營養和風味[1],因此,超高壓技術已經作為商業保鮮手段被應用。近來,有報道表明,超高壓處理對蛋白結構有很大影響[2],蛋白結構的變化又會影響魚肉品質的變化。但是關于超高壓對草魚肉肌原纖維蛋白結構方面的影響卻鮮有報道。作者以草魚為原料,研究超高壓處理對草魚片肌原纖維蛋白結構的影響,包括總巰基和活性巰基、Ca2+-ATPase、表面疏水性、內源熒光光譜、以及拉曼光譜分析,為超高壓技術在淡水魚加工保鮮中的應用提供一定的理論基礎。
1 材料與方法
1.1 材料
草魚:市售。
1.2 設備
UHPF-600 MPa-50 L型超高壓處理設備:包頭科發食品機械有限公司產品;F-7000型熒光光譜儀:日本日立公司產品;RamTracer-200-HS型拉曼光譜儀:美國OptoTrace Technologies公司產品;精密電子天平:梅特勒-托利多儀器(上海)有限公司產品;4K-15型高速冷凍離心機:美國Sigma Sartorius公司產品;D-8941型真空包裝機:Multivac公司產品;實驗室用pH計 FE20:梅特勒-托利多儀器有限公司(上海)產品;紫外-可見分光光度計:上海天美科學儀器有限公司產品。
1.3 方法
1.3.1 樣品預處理 將草魚宰殺去內臟、魚頭、魚皮,用蒸餾水沖洗干凈,取背部魚肉作為研究對象,切成約3 cm×3 cm×1 cm方塊,于蒸煮袋真空密封好后用超高壓設備進行處理,壓力分別為200、300、400、500、600 MPa,保壓時間為 15 min。 未經處理組記為0.1MPa,作為對照組。超高壓后所有樣品,在兩小時內完成蛋白提取,并進行結構測定。
1.3.2 肌原纖維蛋白的提取 制備方法參照Lina R[3]的方法,并略作修改。取適量經過不同超高壓處理的草魚肉,加入5倍體積緩沖液(pH 7.0),混勻,9 000 r/min 4℃離心10 min,棄去上清液,重復上述操作2次。沉淀加入4倍體積緩沖液,10 000 r/min勻漿。靜置30 min以充分溶解蛋白,9 000 r/min 4℃離心20 min,所得上清液即為肌原纖維蛋白溶液。肌原纖維蛋白濃度的測定采用雙縮脲法測定,以牛血清白蛋白(BSA)作為標準蛋白。
1.3.3 總巰基和活性巰基含量測定 參考Yongsawatdigul[4]等的方法,并略作修改。用 0.6 mol/L KCl溶液將肌原纖維蛋白稀釋到 2 mg/mL左右,取1 mL加入4 mL 50 mmol/L磷酸緩沖液。混勻,取 4 mL溶液,加入0.4 mL Tris-HCl緩沖液,40℃水浴 25 min。以0.6 mol/L KCl溶液作為空白,在紫外分光光度計412 nm處測定吸光值。
1.3.4 Ca2+-ATPase活性測定 用0.6 mol/L KCl溶液將肌原纖維蛋白溶液稀釋到4.0~6.0 mg/mL左右。然后取1 mL蛋白溶液,分別加入0.5 mL 0.5mol/L pH 7.0 Tris-HCl緩沖液、0.5 mL 0.1 mol/L CaCl2、1 mL 1 mol/L KCl溶液、6.5 mL 去離子水,最后加入20 mmol/L ATP溶液。置于25℃水浴預熱10 min,之后加入肌原纖維懸濁液,搖勻,等到加入20 mmol/L ATP溶液的時候開始記時,反應5 min后在反應體系中加入2.5 mL冰浴后的體積分數15%的三氯乙酸(TCA)溶液終止反應。另一組預先加入TCA溶液作為空白對照。反應結束后,將所得溶液離心(3 500 g,10 min),采用鉬酸銨法測定上清液中釋放的無機磷含量。
1.3.5 表面疏水性分析 參考Riebroya[5]等的方法測定其熒光強度,激發波長370.0 nm,發射波長480.0 nm。空白用氯化鉀緩沖液替代。
1.3.6 內源熒光測定 把肌原纖維蛋白溶液用0.6mol/L KCl溶液稀釋到0.05 mol/mL,選取激發波長295 nm,激發和發射狹縫寬度為2.5 nm,波長掃描范圍為300~400 nm,掃描速度為12 000 nm/min,每個樣品做3次平行。
1.3.7 拉曼光譜分析 拉曼光譜用 RamTracer?-200-HS便攜式拉曼儀測量,激發波長785 nm,掃描范圍100~3 300 cm-1,光譜分辨率 6 cm-1。
1.3.8 數據處理 數據結果采用Origin 9.1進行分析處理。
2 結果與討論
2.1 總巰基和活性巰基含量的變化
肌原纖維蛋白分子約含有42個巰基,其中分布在球狀頭部結構的約有24或者26個,反應活性很強,是肌原纖維蛋白活性的重要組成部分。因此可以根據蛋白質巰基的含量變化情況進而預測蛋白質的變性程度[3]。
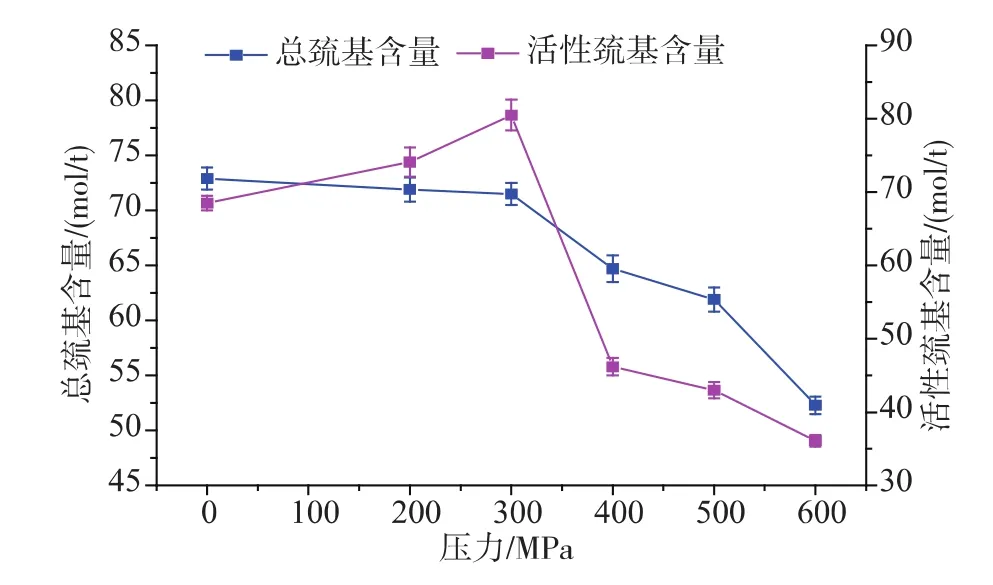
圖1 不同壓力處理后草魚肌原纖維蛋白總巰基和活性巰基變化Fig.1 Changes in total sulfhydryl content and surfacereactive sulfhydryl content activity of myofibrilproteinof grass carp subjected to different high pressure processing
由圖1可以看出,經過超高壓處理過的草魚片,肌原纖維蛋白總巰基含量隨著壓力的增大呈現減小的趨勢,而活性巰基隨著壓力的增大呈現先增大后下降的趨勢。在300 MPa時達到最大值,這與Hormann[6]和Chapleau[7]得到的結果一致。這是由于超高壓處理之后,肌原纖維蛋白的活性位點發生變化或者移動,導致蛋白質分子發生變性從而展開,隱藏在蛋白結構內部的殘基、巰基開始逐漸暴露于蛋白表面,并且隨著壓力的升高,巰基暴露逐漸增多,300 MPa處理后達到最大,而當壓力繼續升高時,蛋白質分子結構因為聚集而變得更加緊密,導致活性巰基含量減少[8]。此外,暴露于表面的巰基易于氧化生成二硫鍵,從而進一步導致總巰基含量的下降。
2.2 表面疏水性的變化
肌原纖維蛋白質的表面疏水性反映的是蛋白質分子表面的疏水性殘基的相對含量,其變化可以表明蛋白質的表面性質發生改變,因此可作為蛋白質結構改變的表征指標。超高壓對蛋白質空間結構的改變對疏水性氨基酸殘基在蛋白質分子內外的分配產生影響,當肌原纖維蛋白分子發生變性時,內部的疏水性氨基酸暴露于表面,能與ANS(1-苯氨基萘-8-磺酸)探針結合呈現熒光,且肌原纖維蛋白的相對熒光強度與其表面疏水性成正相關[9]。
不同的超高壓處理對草魚肌原纖維蛋白表面疏水性的影響見圖2,從圖中可以看出,隨著壓力的不斷增大,表面疏水性一直呈現上升的趨勢,200~600 MPa處理后表面疏水性值分別上升了11%、55%、73%、76%、82%。主要的原因是超高壓處理改變了肌原纖維蛋白的空間構型。
新鮮的草魚肉蛋白質的疏水性基團大多位于蛋白質分子內部,故具有較低的表面疏水性。在蛋白質中具有疏水相互作用的基團為芳香族殘基,它們均勻地分布在肌球蛋白分子尾部,包埋于α-雙螺旋之間的界面內。
經過超高壓處理之后,折疊態的肌原纖維蛋白分子開始伸展,位于蛋白質分子內部的部分疏水性基團外露,引起蛋白質分子表面疏水性的上升(見圖2)。隨著壓力的逐步增大,疏水性基團外露的越來越多,導致疏水相互作用增強,同時這些疏水性殘基就會重新分布,有更多的殘基暴露于蛋白表面,從而表面疏水性增加[7]。
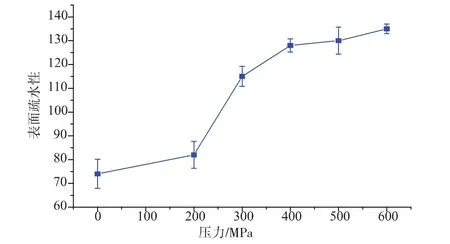
圖2 不同壓力處理后草魚肌原纖維蛋白表面疏水性的變化Fig.2 Changes in surface hydrophobicityof myofibril protein of grass carp subjected to different high pressure processing
2.3 Ca2+-ATPase活性的變化
Ca2+-ATPase活性源于肌原纖維蛋白的球狀頭部結構,其活性與頭部區域緊密相關,是分析蛋白質品質的重要指標[11]。
圖3為不同壓力處理對肌原纖維蛋白Ca2+-ATPase活性的影響,由圖可以看出,隨著壓力的增大,Ca2+-ATPase的活性呈現一直下降的趨勢。經200 MPa處理,Ca2+-ATPase的活性由對照組的0.52 μmol/(mg·min)降低至 0.46 μmol/(mg·min),其活性下降12%,當壓力超過200 MPa時,Caμ-ATPase的失活速度逐漸加快,該研究結果同KO[10]等人一致。300 MPa以上的超高壓處理可以明顯降低Ca2+-ATPaes的活性,由圖 3 分析可知,300、400、500、600 MPa分別降低了27%、63%、79%、85%,隨著壓力的增大,其活性顯著下降。其中600 MPa處理下降的最為明顯。雒莎莎等人研究顯示,超高壓處理明顯抑制了肌球蛋白Ca2+-ATPase的活性,當壓力≥150 MPa時,Ca2+-ATPase的失活速度明顯加快。另外,總巰基含量的減少,導致蛋白分子內作用加強,會加劇Ca2+-ATPase的活性下降。
Ca2+-ATPase活性的降低表明超高壓能夠誘導肌原纖維蛋白頭部變性,在相對較低的壓力下,肌原纖維蛋白(粗絲的主要成分)就開始聚集,兩個頭部S1和S2首先融合成一個,接著它們聚集成一簇,尾部呈放射狀的伸在外面。肌球蛋白分子的聚集不斷增加,蛋白質的疏水性上升,這是因為超高壓作用使肌球蛋白頭部的疏水基團暴露出來,使肌球蛋白頭部間的疏水作用加強,從而導致肌原纖維蛋白聚集[11]。
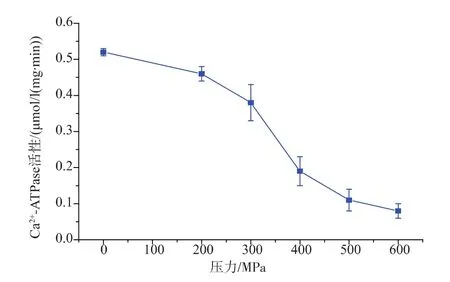
圖3 不同壓力處理后草魚肌原纖維蛋白Ca2+-ATPase活性的變化Fig.3 Changes in Ca2+-ATPase activity of myofibril protein of grass carp subjected to different high pressure processing
2.4 內源熒光光譜分析
超高壓處理會引起蛋白構象的改變,甚至引起蛋白質分子變性。在此過程中,部分氨基酸中的內源熒光生色基團的微環境和位置會發生相應的改變,因此,研究內源熒光光譜是監測超高壓處理對蛋白質構象影響的有效手段。內源熒光分析的主要是色氨酸(Trp)殘基,在295 nm處可只激發色氨酸殘基,從而可以排除酪氨酸殘基的干擾。圖4為不同壓力處理后得到的內源熒光光譜圖,從圖4中可以看出,在壓力的作用下,肌原纖維蛋白的熒光光譜形狀變化不明顯,而對最大峰值以及強度的變化較為明顯。肌原纖維蛋白的λmax集中在336 nm附近。有研究學者研究表明,λmax與色氨酸殘基所處的微環境有著一定的關系[12]。λmax<330 nm,表明色氨酸殘基位于蛋白質分子內部的非極性環境中。λmax>330 nm,表明色氨酸殘基位于蛋白質分子外部的極性環境中。如圖中所示,λmax位于336 nm附近,表明色氨酸殘基位于蛋白質分子外部的極性環境中。隨著壓力的不斷增大,熒光光譜的熒光強度不斷增大,到600 MPa時達到最大。并且,λmax發生了一定的紅移現象。這說明色氨酸殘基可能是轉移到蛋白分子外部,暴露在了極性環境中,而經過超高壓處理后,熒光強度不斷增強,色氨酸殘基都暴露到分子表面極性強的環境中,這表明超高壓處理改變了色氨酸的微環境,對肌原纖維蛋白的三級結構產生影響。
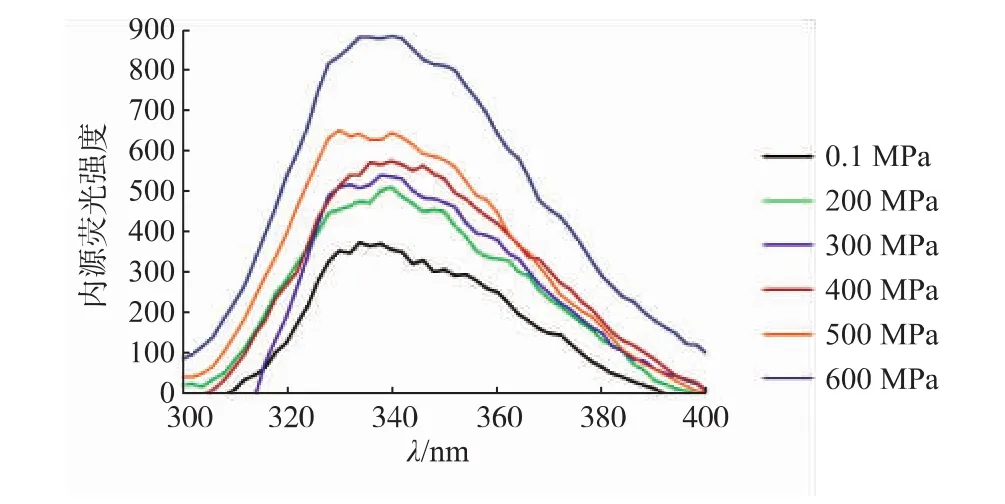
圖4 不同壓力處理后草魚肌原纖維蛋白內源熒光圖Fig.4 Changes inEndogenous fluorescence spectrumof myofibrilprotein ofgrass carp subjected to different high pressure processing
2.5 拉曼光譜分析
拉曼光譜是一種振動光譜技術,具有指紋式的高分辨率,對于分子結構分析有顯著作用,是研究蛋白構象的一種重要手段。
經超高壓處理后,草魚片肌原纖維蛋白的拉曼圖譜見圖5。在拉曼圖譜上最突出的譜帶位于1 450 cm-1左右區域和1 650 cm-1左右區域,1 450 cm-1左右區域是C-H(CH2)彎曲振動譜帶。C-H彎曲振動的譜帶強度變化與蛋白質的三級結構的變化相關,其強度減弱表明部分內部疏水性殘基的暴露[13]。酰胺I帶集中于1 650 cm-1左右區域,常常用來被表征蛋白質二級結構[14]。有研究顯示,位于1 600~1 700 cm-1左右的酰胺I帶與蛋白骨架構象類型相關:主要來自于C=O伸縮振動和N-H平面彎曲振動。該譜帶由位于1 645~1 660 cm-1(α-螺旋),1 665~1 680 cm-1(β-折疊),1 660~1 665 cm-1(無規則卷曲)三部分重疊的譜帶構成。
3 結 語
超高壓處理導致草魚肌原纖維蛋白空間構象顯著變化。隨著壓力的增大,Ca2+-ATPase活性、總巰基含量均顯著下降,活性巰基先增加后降低。肌原纖維肌球蛋白的球狀頭部結構在壓力作用下展開,疏水集團暴露,二硫鍵增多,并隨著壓力的進一步增大,蛋白質出現聚集。內源熒光和拉曼光譜表明,超高壓處理改變了肌原纖維蛋白二級、三級結構,導致蛋白質變性。
[1]JIANG Bo,LI Yungao,MIU Ming,et al.Effects of high hydrostatic pressure on activity of enzymes and the mechanisms[J].Journal of Food Science and Biotechnology,2015,34(6):561-568.(in Chinese)
[2]QIU C,XIA W,JIANG Q.Pressure-induced changes of silver carp (Hypophthalmichthys molitrix) myofibrillar protein structure[J].European Food Research and Technology,2014,238(5):753-761.
[3]LINA R,YANSHUN X,QIXING J,et al.Investigation on structural changes of myofibrillar proteins from silver carp(Hypophthalmichthys molitrix) during frozen storage[J].Food Science and Technology Research,2013,19(6):1051-1059.
[4]YONGSAWATDIGUL J,PARK J W.Thermal denaturation and aggregation of threadfin bream actomyosin[J].Food Chemistry,2003,83(3):409-416.
[5]RIEBROY S,BENJAKUL S,VISESSANGUAN W,et al.Acid-induced gelation of natural actomyosin from Atlantic cod(Gadus morhua) and burbot(Lota lota)[J].Food Hydrocolloids,2009,23(1):26-39.
[6]HOFMANN K,HAMM R.Sulfhydryl and disulfide groups in meats[J].Advances in Food Research,1978,24:1-111.
[7]CHAPLEAU N,MANGAVEL C,COMPOINT J P,et al.Effect of high-pressure processing on myofibrillar protein structure[J].Journal of the Science of Food and Agriculture,2004,84(1):66-74.
[8]CHAPLEAU N J,de Lamballerie-Anton M I.Changes in myofibrillar proteins interactions and rheological properties induced by high-pressure processing[J].European Food Research and Technology,2003,216(6):470-476.
[9]CHOI S M,MA C Y.Structural characterization of globulin from common buckwheat (Fagopyrum esculentum Moench) using circular dichroism and Raman spectroscopy[J].Food Chemistry,2007,102(1):150-160.
[10]BENJAKUL S,SEYMOUR T A,MORRISSEY M T,et al.Physicochemical changes in Pacific whiting muscle proteins during iced storage[J].Journal of Food Science,1997,62(4):729-733.
[11]KO W C,JAO C L,HSU K C.Effect of hydrostatic pressure on molecular conformation of tilapia (Orechromis niloticus) myosin[J].Journal of Food Science,2003,68(4):1192-1195.
[12]ZHOU A,LIN L,LIANG Y,et al.Physicochemical properties of natural actomyosin from threadfin bream (Nemipterus spp.)induced by high hydrostatic pressure[J].Food Chemistry,2014,156:402-407.
[13]LEFEVRE F,FAUCONNEAU B,THOMPSON J W,et al.Thermal denaturation and aggregation properties of Atlantic salmon myofibrils and myosin from white and red muscles[J].Journal of Agricultural and Food Chemistry,2007,55(12):4761-4770.
[14]LOPES D C F,DELVIVO F M,SILVESTRE M P C.Use of activated carbon for removing phenylalanine from reconstituted skim milk powder hydrolysates[J].LWT-Food Science and Technology,2005,38(5):447-453.
