hsp90α基因四環素誘導調控表達質粒的構建及功能驗證
2018-06-15 16:56:08尚冰清周丹朱宏劉昱郭麗麗湯仁仙寇艷波
中西醫結合心血管病電子雜志 2018年8期
尚冰清 周丹 朱宏 劉昱 郭麗麗 湯仁仙 寇艷波

【摘要】目的 構建Hsp90α基因的單質粒四環素/強力霉素誘導表達系統,并驗證其不同表達量對HepG2肝癌細胞增殖的影響。方法 通過PCR,分別從pRetroX-Tet-On Advanced質粒和pcDNA3.1-EGFP質粒中擴增rtTA表達盒和EGFP基因,依次將rtTA和EGFP基因克隆入pTRE-Tight載體中,pRetroX-Tight載體中,獲得EGFP標記的Tet-on單質粒系統。基于本系統,構建DOX誘導的Hsp90α表達質粒pRetroX-Hsp90α-EGFP-Tet-On,然后采用脂質體轉染HepG2細胞,以不同濃度(0,100,500,2500 ng/mL)的DOX誘導,通過熒光顯微鏡、熒光定量PCR以及Western blot檢測EGFP和目的蛋白Hsp90α的表達。結果 轉染質粒載體后的HepG2細胞經不同濃度的DOX 24 h后,RT-PCR和Western blot結果顯示一致,隨著DOX濃度的增大,Hsp90α基因表達量逐漸增加,DOX 500 ng/mL時時,基因表達的水平達到高峰;加入DOX 48 h后,細胞的增殖明顯高于對照組,且存在劑量依賴性。結論 該研究成功地應用四環素基因表達調控系統調控了Hsp90α基因在HepG2細胞內表達及促腫瘤細胞增殖的功能,為進一步探討Hsp90α基因與腫瘤的增殖和侵襲之間的關系提供理論基礎。
【關鍵詞】四環素;基因表達調控;Hsp90α基因;HepG2細胞
【中圖分類號】R394 【文獻標識碼】B 【文章編號】ISSN.2095-6681.2018.8..04
基因的過表達和沉默是研究基因功能的重要手段。但是由于某些基因的編碼產物在特定的細胞時期對細胞具有毒性或其減少嚴重影響細胞正常生長甚至導致細胞死亡,給這些基因生物學功能的闡明帶來的極大地困難。此外,隨著研究的深入,有報道表明某些基因表達水平的不同所引起的生物學表型也截然不同[1-2]。因此,可人工調控表達系統的建立對基因功能的研究將具有非常重要的意義。
近些年研究者們開發出了許多種類的可誘導表達系統,1995年由Gossen等[3]發現并改造的Tet-on系統尤為受關注。環境中沒有四環素(tetracycline,Tc)或其衍生物如強力霉素(doxycycline,DOX) 存在時下游基因處于表達抑制狀態,當Tc或DOX存在時下游基因啟動表達[4]。傳統的Tet-on系統具有可控性好、本底表達水平低、細胞毒性小等特點,但是它也存在一定的不足,如傳統的Tet-on系統為雙載體系統需要同時對TetR表達載體和目的基因表達載體同時進行遺傳操作,并且目的基因過表達無法統一檢測或實時監測。
熱休克蛋白90(heat shock proteins 90,HSP 90)在細胞中含量豐富,約占細胞蛋白總量的1%-2%,其靶蛋白多在細胞生長調控及存活方面發揮著重要作用。研究發現HSP90α在腫瘤細胞中的表達量為正常細胞的2-10倍,例如:HBV相關性肝細胞癌,其含量的表達與腫瘤分級分期相聯系[5-6],提示其與細胞的惡性轉化密切相關,并且HSP90α已作為腫瘤標記物應用于癌癥的早期篩查。不同表達水平的HSP90α對肝癌發生發展的作用研究將一定程度上加深人們對肝癌發生機制的認識。
該研究通過對傳統雙載體逆轉錄病毒Tet-on系統的改造建立了單載體Tet-on系統,并在啟動子下游引入增強綠色熒光蛋白(Enhancegreen Fluorescent Protein,EGFP)編碼基因使過表達的目的蛋白標記EGFP標簽,便于后續檢測和實時觀察。新的Tet-on-EGFP系統應用于肝癌細胞HepG2中HSP90α的過表達,通過使用不同濃度DOX的誘導不同水平Hsp90α過表達,為進一步機制研究提供了有力的遺傳操作工具。
1 材料與方法
1.1 試劑與儀器
肝癌細胞HepG2(徐州醫科大學免疫與代謝重點實驗室);pRetroX-Tet-on雙載體系統 (中國醫學科學院醫藥生物技術研究所);限制性內切酶及連接酶(Thermo Fisher Scientific);質粒提取試劑盒,PCR產物純化試劑盒,凝膠回收試劑盒 (Omega);PCR試劑盒 (Takara); DMEM培養基(Hyclone);胎牛血清 (Serana);Lipofectamine 2000 (Invitrogen);嘌呤霉素 (Sigma);強力霉素 (生工生物);逆轉錄試劑盒(Takara);熒光定量PCR試劑盒(Takara);EGFP、GAPDH蛋白抗體 (Abcam);ECL發光劑(Bio-Rad);CCK-8試劑盒(日本同仁);恒溫CO2培養箱(Thermo Fisher Scientific);熒光顯微鏡 (Olympus)。
1.2 HepG2細胞培養
HepG2細胞于含有10% 胎牛血清的DMEM高糖培養基中,37℃,5% CO2,一般2-3天傳代一次。
1.3 單載體Tet-On-EGFP系統載體構建
1.3.1 目的基因hsp90αORF的擴增與純化 由NCBI基因庫中獲取hsp90α基因編碼序列。利用軟件Primer 5.0設計引物并分別在上游和下游引物5'端引入限制性內切酶NotI和MluI識別位點及對應的保護堿基,上游引物序列:5'AAGGAAAAAAGCGGCCGCATGCCCCCGTGTTCGGGCG 3',下游引物序列:5' CGACGCGTTTAGACTACTTCTTCCATGCGTGATGTGTCG 3'。以肝癌細胞HepG2 cDNA為模板經PCR擴增獲得hsp90α全長基因片段(產物大小2579 bp)。利用純化試劑盒純化PCR產物,純化產物進行電泳分析。
1.3.2 表達質粒
pRetroX-Hsp90a-EGFP-Pur的構建 以質粒pcDNA3.1-EGFP為模板PCR擴增EGFP編碼基因,通過MluI和EcoRI雙酶切連接連入pRetroX-Tight-Pur,獲得質粒pRetroX-EGFP-Pur。將1.3.1中純化的PCR產物和pRetroX-EGFP-Pur用NotI和MluI雙酶切,連接獲得質粒pRetroX-Hsp90a-EGFP-Pur,測序無誤后用于后續實驗。
1.3.3 rtTA表達盒的擴增和單質粒Tet-on系統的構建
以pRetroX-Tet-On Advanced為模板PCR 擴增rtTA表達盒,并在其兩端引入限制性內切酶BglII識別位點。將rtTA表達盒和pRetroX-Hsp90a-EGFP-Pur經BglII酶切及去磷后連接獲得最終質粒pRetroX-Hsp90a-EGFP-Tet-On。
1.4 細胞轉染及篩選
HepG2生長至80%時利用Lipfectamine 2000進行轉染,轉染后5 h更換含有10% 胎牛血清的培養基,并于48 h后胰酶消化重新接種于含有1 mg/ml 嘌呤霉素的新鮮培養基中進行篩選,能夠正常生長的細胞即含有pRetroX-Hsp90a-EGFP-Tet-On質粒。
1.5 RT-PCR
收集不同濃度DOX誘導后的HepG2細胞,提取RNA,取總RNA 1 μg進行逆轉錄獲得總cDNA,以不同組的cDNA為模板利用Roche Light Cycler 480熒光定量PCR儀檢測hsp90α轉錄情況,內參為β-actin,引物序列如下:5'GGCATCGTGATGGACTCCG 3',5'GCTGGAAGGTGGACAGCGA 3' (β-actin);5'TGCCTCTGGTGATGAGATGGTT 3' ,5'GGTGACTGACACTAAAGTCTTCCCC 3' (hsp90α)。
1.6 Western Blot
收集不同濃度DOX誘導后的HepG2細胞,RIPA裂解液裂解,BCA法測定蛋白濃度。每孔上樣總蛋白15 mg進行SDS-PAGE,轉印至PVDF膜后用5%脫脂奶粉室溫封閉2 h,一抗室溫孵育2 h洗滌后二抗孵育1 h再次洗滌后檢測EGFP和內參GAPDH強度。
1.7 細胞增殖實驗
HepG2細胞消化后血球板計數,調整細胞濃度至4×104個/ml,分為四組(分別加入DOX使終濃度為0,100,250,500 ng/mL)接種于96孔板中每孔100 μl,每組15孔(3個時間點,每個時間點5個平行),于0 h,24 h和48 h通過CCK-8試劑盒檢測細胞增殖情況。
1.8 統計學方法
采用SPSS 16.0統計軟件進行統計分析,實驗數據采用“x±s”表示,多個樣本均數的比較用方差分析,P<0.05為差異有顯著性意義。
2 結 果
2.1 單載體Tet-On-EGFP系統載體構建
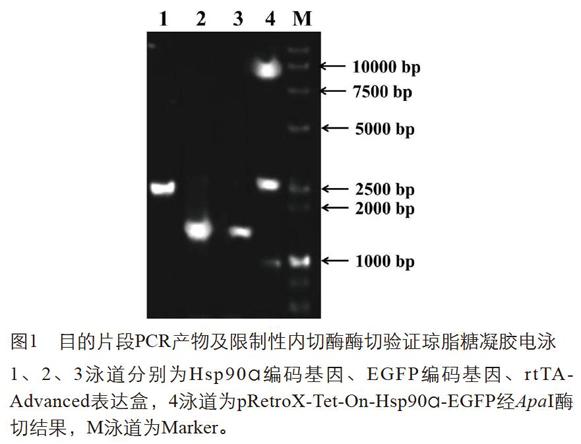
分別通過PCR擴增EGFP編碼基因、HSP90α編碼基因、rtTA-Advanced表達盒,目的產物經凝膠回收后依次進行酶切、純化、連接得到目的基因表達載體pRetroX-Tet-On-Hsp90α-EGFP,如圖1所示,1、2、3泳道為PCR擴增結果,Hsp90α編碼基因2565 bp,EGFP編碼基因1607 bp,rtTA-Advanced表達盒長1525 bp。pRetroX-Tet-On-Hsp90α-EGFP通過ApaI限制性內切酶消化驗證,產生條帶分別為8247 bp和2666 bp和998 bp,載體測序無誤后用于后續實驗。
2.2 DOX誘導目的蛋白的表達
通過脂質體法將目的基因表達載體pRetroX-Tet-On-Hsp90α-EGFP轉染至HepG2細胞中,經G418(500 μg/ml)篩選后得到穩定轉染細胞株,分別添加終濃度為0,100,500,2500 ng/ml的DOX誘導基因表達,24 h后通過熒光顯微鏡、熒光定量PCR以及Western blot檢測EGFP和目的蛋白的表達,并比較不同濃度DOX誘導能力的差異。如圖2A所示,轉染基因表達載體pRetroX-Tet-On-Hsp90α-EGFP的HepG2細胞添加DOX誘導24 h后能夠觀察到明顯的綠色熒光,而未添加DOX基本無熒光,同樣Western blot結果表明EGFP隨DOX濃度不同出現表達量的差異,DOX為500 ng/ml時EGFP表達量最高(圖2B)。與之相一致的是,熒光定量PCR結果表明隨著DOX存在時hsp90α存在明顯的誘導表達現象,且500 ng/mL誘導作用最強(圖2C)。
2.3 hsp90α表達水平影響HepG2增殖
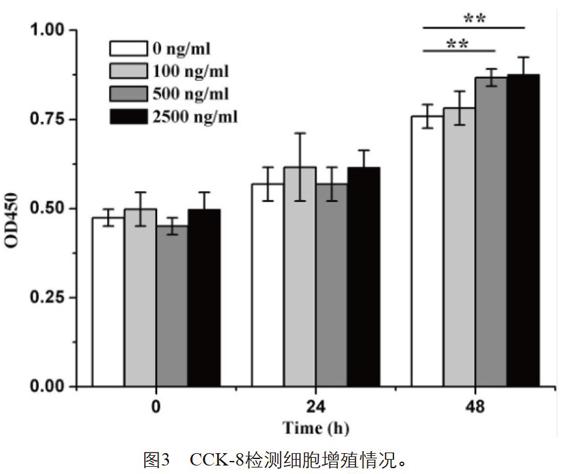
為進一步檢測,不同濃度DOX誘導Hsp90α表達對HepG2增殖的影響,進行了CCK-8實驗。穩定轉染pRetroX-Tet-On-Hsp90α-EGFP的HepG2細胞消化后調整細胞濃度,分為四組,接種于96孔板中分別用不同濃度DOX誘導,每隔24 h利用CCK-8試劑盒檢測細胞增殖情況。由圖3可知,未經DOX誘導前,四組細胞數目基本一致,誘導48 h后添加500 ng/ml或2500 ng/ml DOX組增殖明顯高于對照,而100 ng/ml DOX組與對照組沒有統計學差異,說明Hsp90α促進HepG2增殖具有一定的劑量依賴性。
3 討 論
大量研究已證實:四環素調控系統能夠有效降低基因泄露[7-8]、時間空間特異性調控基因表達[9],且相對于其他系統的誘導因子如熱休克、激素,Tc作為抗生素對細胞毒性小,誘導所需的劑量低,可應用于長期調控表達的細胞、動物模型。
四環素調控系統包括Tet-on與Tet-off系統。該實驗選擇Tet-on系統,在有DOX時,tetR可與tetO結合目的基因得到表達,撤去dox 轉錄即被抑制[10]。由于活體攝取要比清楚快得多,因此其在基因轉錄方面要快于Tet-off系統。
相比較于傳統的Tet-on系統,需要兩次篩選轉染,操作繁瑣,該實驗設計的Tet-on單質粒載體系統,經一次轉染即可用于實驗,操作相對簡單。在啟動子下游引入EGFP編碼序列使過表達的目的蛋白C端標記EGFP標簽,便于后續檢測和實時觀察。不同濃度DOX展現出不同水平的誘導能力,一定程度上可用于某些蛋白的定量表達,為這些蛋白生物學功能的揭示提供了更有力的工具。
臨床統計數據表明HSP90α與肝癌的發生具有明顯的相關性,其在腫瘤細胞的增殖和分化、生存和移動及血管形成中發揮重要作用[11]。有研究表明,腫瘤細胞內過表達的HSP90α通過外泌體分泌于細胞外,與底物蛋白結合,促進腫瘤細胞的侵襲遷移,在腫瘤轉移過程中發揮作用。其中,基質金屬蛋白酶2(MMP-2)是腫瘤細胞外HSP90α的底物之一,分泌于細胞外的 HSP90α可能通過激活其底物蛋白MMP-2增強肝癌細胞侵襲遷移能力。同時,HSP90α也可以與其他伴侶分子聯合作用,實驗結果顯示,HSP90α和HSP70可能通過結合形成復合體調控MMP-2的成熟和活化,參與肝癌轉移潛能的形成[12]。HSP90α也可以通過調節細胞周期促進細胞增殖。腫瘤細胞在低氧、營養缺乏微環境中生存尤其依賴HSP90α。Hsp90α參與低氧誘導因子的信號傳導通路。HSP90α可作為一種正向調節因子參與血管再生[13]。
繼2013年在世界上首次證明HSP90α是一個全新腫瘤標志物并可用作肺癌檢測后,清華大學生命科學學院羅永章團隊在世界上首次證明,HSP90α可用于肝癌患者的檢測,靈敏度比現行檢測方法甲胎蛋白(AFP)大幅提高。目前,這一發現已被國家食品藥品監督管理總局批準臨床使用[14]。除此之外,亦有實驗證實,HSP90α高表達與肝癌分化程度相關,而與患者性別、病理類型、病理分期無關。楊婧[15]等證明HSP90α單克隆抗體不僅能明顯抑制肝癌移植瘤生長,而且具有抑制肝癌干細胞侵襲、成球和耐藥的能力。說明HSP90α在腫瘤干細胞侵襲和存活過程中起重要作用,具有作為肝癌干細胞治療靶點的潛力。
臨床統計數據表明Hsp90α與肝癌的發生具有明顯的相關性,且Hsp90α已作為腫瘤標記物用于肝癌早期的篩查。雖然已有研究提示Hsp90α高表達能夠促進肝癌細胞侵襲和增殖[16],但是并沒有進一步研究揭示Hsp90α表達量與肝癌細胞侵襲和增殖的具體關系。該研究結果顯示hsp90α轉錄上調可以促進肝癌細胞HepG2的增殖,且增殖能力與hsp90α表達量存在一定的正相關關系,具體的分子機制還有待于深入探討。
參考文獻
[1] Tang R,Kong F,Fan B,et al.HBx activates FasL and mediates HepG2 cell apoptosis throughMLK3-MKK7-JNKs signal module[J].World J Gastroenterol,2012,18(13):1485-1495.
[2] 湯仁仙,石紅麗,范寶峰,等.乙肝病毒X基因對人肝癌細胞株HepG2凋亡的影響[J].四川大學學報(醫學版),2010,41(1):95-100.
[3] Gossen M,Freundlieb S,Bender G,et al.Transcriptional activation by tetracyclines in mammalian cells[J].Science,1995 Jun 23;268(5218):1766-1769.
[4] 宮秀群,馬敏敏,徐格林.四環素基因調控系統的研究進展[J].基礎醫學與臨床,2012,32(2):233-236.
[5] Beasley RP.Hepatitis B virus,the major etiology of hepatocellular carcinoma[J].Cancer ,1988;61:1942-1956.
[6] 李 慧.熱休克蛋白90α在HBV相關性肝細胞癌中的表達及臨床意義[D].廣州:廣州醫科大學,2014.
[7] Urlinger S,Baron U,Thellmann M,et al. Exploring the sequence space for tetracycline dependent transcriptional activators: Novel mutations yield expanded range and sensitivity[J].Proc Natl Acad Sci USA,2000,97(14): 7963-7968.
[8] Deuschle U,Meyer WK,Thiesen HJ.Tetracycline-reversiblesilencing of eukaryoticpromoters[J].Mol Cellul Biol,1995,15(4):1907-1914.
[9] Gallagher AR,Schonig K,BroWn N,et al.Use of the tetracycline system for inducible protein synthesis in the kidney[J].JAm Soc Nephrol,2003,14(8):2042-2051.
[10] Gossen M,Freundlieb S,Bender G,et al.Transcriptional activation by tetracyclines in mammalian cells[J].Science,1995,268(5218): 1766-1769.
[11] Li W,Miao X,Qi Z,et al.Hepatitis B virus X protein upregulates HSP90α expression via activation of c-Myc in human hepatocarcinoma cell line,HepG2[J].Virology Journal,2010,7.45-57.
[12] 楊 婧,曹凱悅,孫立新,等.HSP90α和HSP90β在肝癌中共同上調表達[J].中國腫瘤,2016,5:391-394.
[13] 孔巧巧,鄒志強.熱休克蛋白90α與原發性肝癌關系的研究進展[J].濱州醫學院學報,2016,39(4):289-290.
[14] 羅永章.全新廣譜腫瘤標志物可大幅提高肝癌檢測靈敏度[J].醫藥前沿,2017,3:3-4.
[15] 楊 婧.肝癌干細胞靶向治療的研究[D].北京:北京協和醫學院,2016.
[16] Li W,Miao X,Qi Z, Zeng W,et al.Hepatitis B virus X protein upregulates HSP90alpha expression via activation of c-Myc in human hepatocarcinoma cell line,HepG2[J].Virology Journal,2010,7: 45-57.
本文編輯:李 豆
