千屈菜科中rpl2基因內含子缺失現象探析
2018-06-20 02:32:48施雨彤鄭紹宇顧翠花
浙江林業科技 2018年2期
施雨彤,徐 梁,鄭紹宇,顧翠花
(1. 浙江農林大學 風景園林與建筑學院,浙江 杭州 311300;2. 浙江省林業科學研究院,浙江 杭州 310023)
千屈菜科Lythraceae植物有31個屬600多個種,大多數植物為草本,灌木或喬木,適應性強,千屈菜屬為本科模式屬,廣布于全世界,主要分布于熱帶和亞熱帶地區,我國有1屬,約47屬種,南北均有[1]。根據植物形態、解剖和植物胚胎學角度證明其與柳葉菜科 Onagraceae在生物進化過程中關系密切,同屬于桃金娘目Myrtiflorae[2]。千屈菜科紫薇屬 Lagerstroemia植物為落葉或常綠的喬木或灌木,根據國際植物名稱檢索表(International Plant Names Index)報道,雖部分種類存在爭議但數量達到近80種[3]。紫薇屬植物大多數種類花大色艷,樹形優美可愛,夏季開花時間長,木質優良,紋理美觀,是園林造景、家具用材的優良樹種,具有較高的經濟價值。國內外紫薇屬植物研究主要集中在新品種選育、繁殖、形態分類、生理生化等方面[4-6]。國內關于紫薇葉綠體基因組內含子的研究較少,而在植物進化過程中基因的內含子丟失現象廣泛發生。對鼠耳芥Arabidopsis thaliana的研究發現內含子丟失與植物基因突變率有明顯相關性[7],這為紫薇葉綠體基因組內含子研究及紫薇育種在分子水平上提供了參考。
真核生物細胞基因 DNA中把編碼區域間隔開來的序列為內含子(Intron),在基因轉錄后剪接去除[8],內含子作為真核生物基因組的組成部分,與基因表達過程關系密切,內含子極大的豐富了轉錄產物的數量和種類,并在mRNA剪接過程中起著復雜的調節作用,從而影響基因表達[9-11]。內含子的缺失在物種進化過程中一直不斷發生,比如豆科Leguminosae山螞蝗族Trib. Desmodieae以及單子葉和雙子葉植物中均有記載[12-13]。rpl2基因(核糖體大亞基蛋白2,ribosomal protein gene 2)是葉綠體中組成RNA基因的核糖體蛋白大亞基基因,其編碼核糖體蛋白L2的過程是系統的關鍵,與植物的光合作用密切相關,對植物的抗性和生長發育起著重要作用。在基因種類和排序上,葉綠體DNA非常保守。在進化過程中,cpDNA的某些結構依然保持不變,為研究物種的進化過程提供了行之有效的途徑。
有學者在屋久島紫薇Lagerstroemia fauriei葉綠體基因組研究的過程中,發現其rpl2基因的內含子存在缺失現象[14],因此,本研究對紫薇屬乃至千屈菜科進行了取樣,針對內含子區域設計了引物,進行PCR擴增,測序分析,來研究rpl2基因內含子缺失發生在屬的水平還是科的水平,以期對后續研究紫薇系統發育過程中基因的變化提供了一定的研究基礎。
1 材料與方法
1.1 實驗材料
本研究共采集9屬25種相關材料(表1),其中部分物種取自國外。
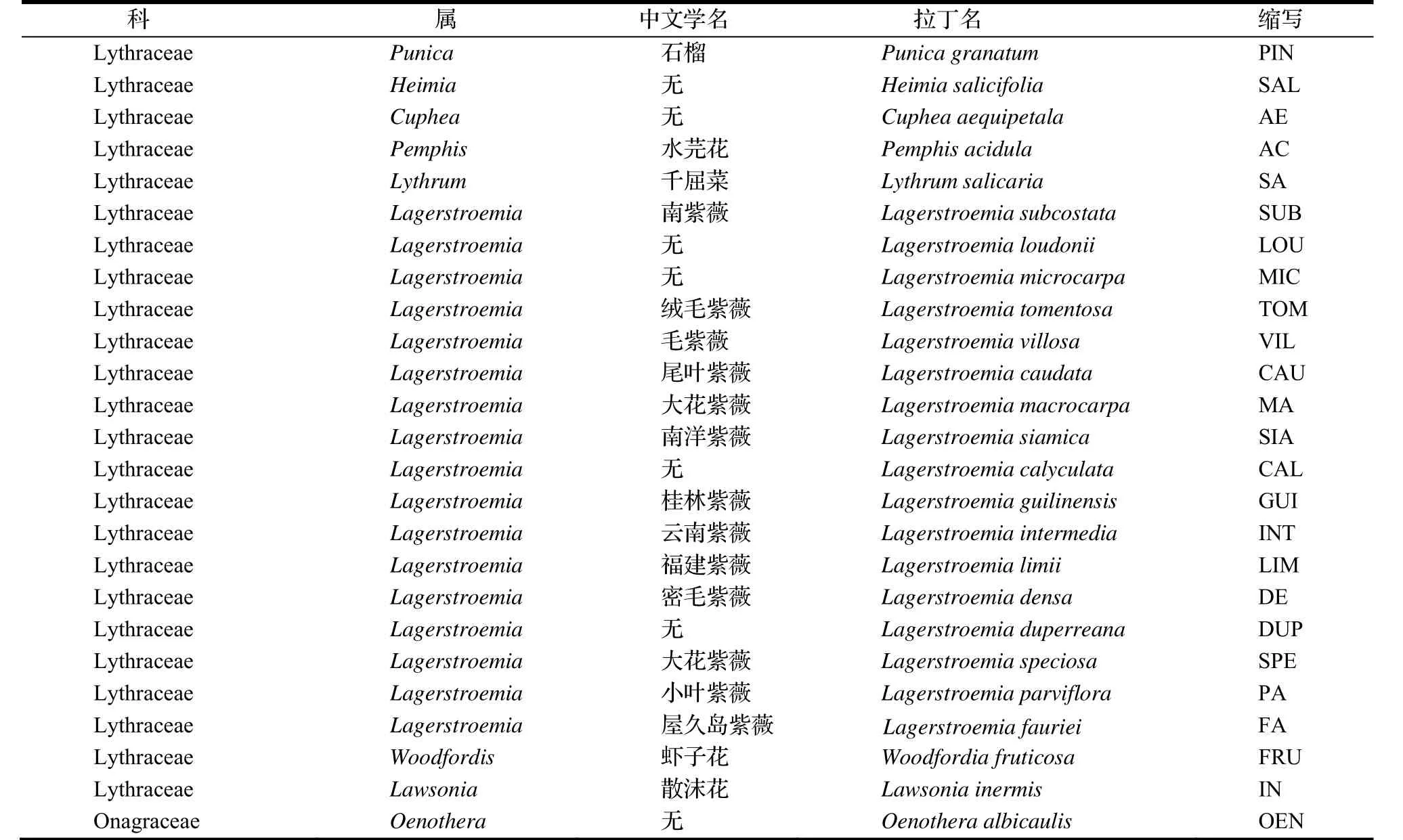
表1 實驗樣本Table 1 Experimental samples
實驗設計采用了與千屈菜科紫薇屬近緣的月見草屬Oenothera植物O. albicaulis作為參照,同時選取模式植物鼠耳芥、近緣科桃金娘科Myrtaceae的物種Angophora costata,Corymbia eximia,Eucalyptus aromaphloia,Eugenia uniflora,Stockwellia quadrifida , Syzygium cumini[15-20]等6種植物的樣品(表2)。取樣后立即投入液氮,于-80℃保存備用。通過植物基因組DNA提取試劑盒提取實驗樣本的總DNA,試劑盒為北京天根生化科技有限公司生產。提取到25種樣本的合格DNA,以進行下一步實驗。部分實驗樣品為國外采集,《中國植物志》中并未收錄,缺乏與之對應的中文名稱,在表格中以拉丁文為準,如 Angophora costata,Corymbia eximia,Eucalyptus aromaphloia,Eugenia uniflora,Stockwellia quadrifida and Syzygium cumini,Heimia salicifolia,Cuphea aequipetala,Oenothera albicaulis等;同時涉及新種如密毛紫薇,參照相關文獻,表中關于石榴屬的分類參照國際最新分類系統[16],具體實驗樣本名錄見表2。

表2 實驗樣本Table 2 Experimental samples
1.2 DNA序列的PCR擴增
根據GenBank中登錄的rpl2基因已知序列,設計了2對引物,正向引物(CAAAACTTCTACCCCAAGCA)和反向引物(TCTTCTTCCAAGTGCAGGAT),以基因組DNA作為模板對樣品進行40 μL體系特異性擴增,擴增體系為:4 μL 10×Taq buffer,0.8 μL dNTP(10 mM),0.4 μL Taq polymerase(5 U·μl-1),0.5 μL 上下游引物(20 pmol·μL-1),所有體系來自上海生工,0.5 μL DNA 模板和 33.3 μL ddH2O,反應程序為,94℃預變性 5 min;94℃變性45 s,55℃預變性45 s,72℃延伸45 s,72℃延伸2 min,共32個循環;4℃保存。反應結束后量取8 μL反應產物,用1%的瓊脂糖凝膠電泳進行檢測。
1.3 PCR產物的回收
按照biomega純化試劑盒說明書進行PCR產物純化回收。
1.4 基因結構和內含子信息分析
利用NCBI數據庫提供的SIpidey[21]工具分析所獲得的rpl2基因結構,并利用PlantCARE[22]對rpl2基因內含子進行轉錄位點分析。
2 結果與分析
2.1 千屈菜科植物與月見草屬物種O. albicaulis rpl2基因比較
本實驗運用PCR技術特異性擴增了紫薇屬的17種和千屈菜科內其他7屬中的物種rpl2基因區域,同時也擴增了千屈菜科外的一種月見草屬物種O. albicaulis作為對比參照。對PCR產物進行1%瓊脂糖電泳檢測,結果如圖1(樣品來自表1)。從圖1中可直觀地看出,不含有rpl2內含子的目標片段的長度約為750 bp,而O. albicaulis內含子的完整片段約為1 400 bp,從片段的長度上直觀的說明了除O. albicaulis之外,千屈菜科植物存在rpl2基因內含子缺失,除了缺失的內含子之外,兩個外顯子部分與其他物種的外顯子序列存在高度相似性。
2.2 紫薇屬植物與千屈菜科內其他物種的比較
同時對紫薇屬內17個物種和千屈菜科內其他7個物種的PCR產物進行了測序,見圖2。由圖2測序結果進一步證實了rpl2基因內含子在24種測試樣本(千屈菜科)中缺失。如圖2A所示,本研究選取對比了7種植物的rpl2基因結構,包括模式植物鼠耳芥、近緣科桃金娘科的物種Angophora costata,Corymbia eximia,Eucalyptus aromaphloia,Eugenia uniflora,Stockwellia quadrifida and Syzygium cumini以及柳葉菜科月見草屬物種O. albicaulis,在這7個物種的rpl2基因中,兩個外顯子之間都包含一個660 bp左右的內含子,圖2B為這7個物種的內含子兩個邊界的部分外顯子序列,圖2C主要對千屈菜科代表物種進行了rpl2基因的測序,藍色背景區域與黃色背景區域均為外顯子序列,中間未出現任何核苷酸。測序結果與比對證明rpl2基因內含子在紫薇屬內發生了缺失現象。

圖1 PCR產物電泳檢測結果Figure 1 Result of electrophoresis test of PCR

圖2 rpl2基因測序結果Figure 2 Result of gene sequencing
3 結論與討論
本研究通過收集的25種樣本,其中紫薇屬樣本17種,千屈菜科其它屬7種,利用柳葉菜科月見草屬物種O. albicaulis為對照。通過試劑盒法提取DNA,并PCR擴增樣本rpl2目標區域基因片段,利用PCR擴增測序測出實驗樣本的rpl2目標區域長度短于非千屈菜科植物,對擴增片段的Sanger測序結果進行比對,確認rpl2內含子缺失現象發生在紫薇屬及千屈菜科內。通過分子生物學的研究,了解其基因組信息所對應的性狀以及其進化歷程,對培育紫薇新品種提供一定的基礎。
之前的研究表明80%~85%的高等植物的葉綠體基因組都有內含子,但是不同物種內含子數目不同[18]。陸生植物的葉綠體基因組在進化上相對保守,其基因內容,基因順序,結構和大小變化都比較小,即便在苔蘚植物中大量內含子的剪接位點仍與鼠耳芥Arabidopsis thaliana相同[10,23-24]。由于數量較少,其基因組的主要結構突變(如倒置,插入或基因和內含子的缺失等)通常可以為特定植物群落的單系進化提供強有力的證據。
內含子的大量丟失導致葉綠體基因組主要結構的突變、縮小[24]。同時根據突變率與內含子丟失的相關性可以推斷出,高突變率的基因導致的內含子丟失速率更高,同時相應的內含子的丟失有時也會影響基因的表達效應,造成表型的變化[25]。內含子多態點位可以作為植物性狀篩選的分子指標,千屈菜科紫薇屬內rpl2內含子的缺失研究結果可以在后續研究中作為性狀篩選的指導。內含子通過mRNA介導反轉錄重組丟失,根據內含子丟失已知模型,內含子的準確丟失在較長的基因中檢測到具有位置偏好性[26]。內含子的丟失在一定程度上反映了物種進化速率;進化較快的物種保留更少的祖先內含子[27,28],該研究中千屈菜科紫薇屬以及其他屬的植物都發生內含子丟失,后續可以通過內含子多態性位點、物種相同基因內含子的丟失來鑒定植物進化程度。
rpl2內含子并不存在于所有的陸生植物。DNA測序結果表明rpl2內含子存在于茄科Nicotiana debneyi中,但在菠菜Spinacia oleracea中未發現。隨后的研究顯示,rpl2內含子在被檢測的6種陸生植物的葉綠體DNA中存在,其中包括2種雙子葉植物,3種單子葉植物和一種苔蘚類植物。這6種陸生植物的內含子的同源性是通過其在基因中的保守位置和其高度保守的一級序列來表示。因此,rpl2內含子存在于被子植物的共同祖先中,但隨后消失在譜系中,從而導致了菠菜中rpl2內含子的缺失[11]。
目前已經確定rpl2基因分布于116科390種被子植物中。rpl2內含子的缺失現象已經在雙子葉植物的其他五個科中出現,虎耳草科Saxifragaceae,旋花科Convolvulaceae,睡菜科Menyanthaceae,牻牛兒科Geraniaceae的兩個屬,和茅膏菜科Droseraceae的一個屬。該內含子缺失表明千屈菜科植物和采樣的其他千屈菜科其他屬的種存在結構上的差異。由此可知,在物種進化過程中rpl2基因內含子已經發生了多次獨立的缺失[14,29]。
[1] 中國科學院中國植物志編輯委員會. 中國植物志[M]. 北京:科學出版社,1983,52(2):62.
[2] GRAHAM S A,HALL J,SYSMA K,et al. Phylogenetic analysis of the Lythraceae based on four gene regions and Morphology[J]. Int J Plant Sci,2005,166(6):995-1017.
[3] 張潔,王亮聲,張晶晶,等. 紫薇屬植物研究進展[J]. 園藝學報,2007,34(1):251-256.
[4] 張啟翔. 紫薇品種分類及其在園林中的應用[J]. 北京林業大學學報,1991,13(4):57-66.
[5] 賈文慶,劉會超,姚連芳. 紫薇花粉萌發特性研究[J]. 西北林學院學報,2007,22(6):18-20.
[6] 童俊,葉要妹,馮彪,等. 秋水仙素誘導三種紫薇多倍體的研究[J]. 園藝學報,2009,36(1):127-132.
[7] 楊宇飛. 鼠耳芥和琴葉鼠耳芥中內含子丟失和突變率關系[D]. 北京:北京師范大學,2013,10-20.
[8] Alberts B,Johnson A D,LEWIS J. Molecular biology of the cell [J]. Garland Sci,2008,10(9):8153-4072.
[9] 王玲平. 不同生物中內含子大小和豐度系統分析[D]. 北京:中國科學院大學,2014.
[10] KREBS J E,GOLDSTEIN E S,KILPATRICK S T. Lewin’s Gene X [M].Beijing: Science Press,2009:58-62.
[11] CHOW L T,GELINAS R E,BROKER T R,et al. An amazing sequence arrangement at the 5' ends of adenovirus 2 messenger RNA[J]. Cell,1977,12(1):1-8.
[12] DOWNIE S R,OLMSTEAD R G,ZURAWSKI G,et al. Six independent losses of the chloroplast DNA rpl2 intron in Dicotyledons: molecular and phylogenetic implications[J]. Evolution,1991,45:1245-1259.
[13] BAILEY C D,DOYLE J J,KAJITA T,et al. The Chloroplast rpl2 intron and ORF184 as phylogenetic markers in the Legume Tribe Desmodieae[J]. Syst Bot,1997,22:133-138.
[14] GU C H,TEMBROCK L R,JOHNSON N G,et al. The complete plastid genome of Lagerstroemia fauriei and loss of rpl2 intron from Lagerstroemia (Lythraceae)[J]. Plos One,2016,11(3):1-18.
[15] GRAHAM S A,HALL J,SYTSMA K,et al. Phylogenetic analysis of the Lythraceae based on four gene regions and morphology[J]. Int J Plant Sci,2005,166(6):995-1017.
[16] HILL K D,JOHNSON L A S. Systematic studies in the eucalypts7. A revision of the bloodwoods, genus Corymbia (Myrtaceae)[J]. Telopea,1995,6(2-3):185-504.
[17] CHAPPILL J,LADIGES P Y,BOLAND D. Eucalyptus aromaphloia Pryor & Willis-a redifinition of geographical and morphological boundaries[J]. Aust J Bot,1986,34(4):395-412.
[18] TURCHETTO-ZOLET A C,SALGUEIRO F,TURCHETTO C,et al. Phylogeography and ecological niche modelling in Eugenia uniflora(Myrtaceae) suggest distinct vegetational responses to climate change between the southern and the northern Atlantic Forest[J]. Bot J Lin Soc,2016,182(3):670-688.
[19] CARR D J,CARR S G M,HYLAND B P M,et al. Stockwellia quadrifida (Myrtaceae), a new Australian genus and species in the eucalypt group[J]. Bot J Lin Soc,2015,139(4):415-421.
[20] HELMST?DTER A. Syzygium cumini (L.) Skeels(Myrtaceae)against diabetes-125 years of research [J]. Pharmazie,2008,63(2):91-101.
[21] KAPUSTIN Y,SOUVOROV A,TATUSOVA T,et al. Splign: algorithms for computing spliced alignments with identification of paralogs[J].Biol Direct,2008,3(1):20.
[22] LESCOT M,DéHAIS P,THIJS G,et al. A database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences [J]. Nucl Acids Res,2002,30(1):325-327.
[23] 楊楨. microRNA相關的生物信息學與進化分析[D]. 上海:復旦大學,2011,80-90.
[24] ROESLER K R,SSHORROSH B S,OHLROGGE J B. Structure and expression of an Arabidopsis actyle-coemzyme A carboxylase gene[J].Plant Physiol,1994,105:611-617
[25] 陳兵,文建凡. 內含子在生物信息學研究和基因工程中的應用[J]. 生命的化學,2010,30(1):59-63.
[26] ROY S W,PENNY D. Patterns of intron loss and gain in plants: intron loss-dominated evolution and genome-wide comparison of O. sativa and A. thaliana[J]. Mol Biol Evolut,2007,24(1):171-181.
[27] 金谷雷. 內含子進化模式研究[D]. 福州:福建農林大學,2007,29-32.
[28] 張尚宏,屈良鵠. 基因組的進化與內含子中的基因的進化[J]. 中山大學學報,1999,38(1):50-52.
[29] 靳霞,呂占軍. 內含子的進化及基因表達調節[J]. 生命的化學,2008,28(1):33-35.
猜你喜歡
英語世界(2023年10期)2023-11-17 09:18:18
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
