自噬相關(guān)基因在煙曲霉自噬時(shí)的表達(dá)
2018-07-09 10:58:24邵建業(yè)孫林趙巍王斌
生物化工 2018年3期
關(guān)鍵詞:水平
邵建業(yè),孫林,趙巍,王斌
(1.青島大學(xué)生命科學(xué)學(xué)院,山東青島 266071;2.青島大學(xué)基礎(chǔ)醫(yī)學(xué)院,山東青島 266071)
自噬與癌癥、神經(jīng)退行性疾病、各種人類發(fā)育過程有關(guān),相關(guān)的自噬研究在過去十幾年中顯著增加[1]。在真菌中,cAMP信號(hào)通路參與多種生物學(xué)過程,并且已被證明對免疫抑制的侵襲性曲霉病鼠科動(dòng)物模型的毒力至關(guān)重要。cAMP途徑的主調(diào)控因子PKA是真核生物中重要的應(yīng)激反應(yīng)調(diào)節(jié)劑[2]。PKA為異四聚體,是細(xì)胞中調(diào)節(jié)生長、代謝、增殖的一個(gè)重要的信號(hào)分子。當(dāng)cAMP與調(diào)節(jié)亞基結(jié)合時(shí),發(fā)生構(gòu)象變化,釋放催化亞基使其自磷酸化并磷酸化下游靶標(biāo)。當(dāng)cAMP結(jié)合調(diào)節(jié)亞基時(shí),會(huì)發(fā)生構(gòu)象變化,其釋放催化亞基自動(dòng)磷酸化下游目標(biāo)[3]。PKA信號(hào)通路參與了煙曲霉的生長、孢子的形成過程以及影響煙曲霉的毒力,但其對曲霉菌自噬的影響并不明確。
為研究煙曲霉中PKA與其自噬水平的關(guān)系,本實(shí)驗(yàn)采用ΔPkaR突變株和WT株,從孢子的存活率、菌絲營養(yǎng)獲取能力、自噬小體的形態(tài)以及自噬相關(guān)的基因表達(dá)來反應(yīng)自噬水平,驗(yàn)證煙曲霉中的自噬是否受PKA通路的調(diào)控,為絲狀真菌中的自噬機(jī)制提供新的實(shí)驗(yàn)證據(jù)[4],爭取為預(yù)防和治療IA提供一個(gè)新的靶點(diǎn)。
1 材料和方法
1.1 材料
1.1.1 菌株
煙曲霉(A.fumigatus)野生菌株(簡稱WT)為美國辛辛納提大學(xué)病理和實(shí)驗(yàn)醫(yī)學(xué)系Judith C Rhodes教授惠贈(zèng),PKA調(diào)節(jié)亞基突變株(簡稱ΔPkaR)為本實(shí)驗(yàn)室構(gòu)建,置于37℃恒溫培養(yǎng)箱中孵育。
1.1.2 培養(yǎng)基
AMM液體培養(yǎng)基、AMM固體培養(yǎng)基、YG液體培養(yǎng)基、YG固體培養(yǎng)基、WA培養(yǎng)基。
1.2 方法
1.2.1 營養(yǎng)缺乏時(shí)營養(yǎng)吸收能力的檢測
(1)收取AMM培養(yǎng)基上新鮮的孢子;(2)在4℃、4500r/min條件下離心5min,再用無菌蒸餾水清洗兩次,然后重懸于無菌蒸餾水中;(3)顯微計(jì)數(shù),稀釋孢子濃度至1×104個(gè)/mL,用移液槍吸取孢子懸浮液,用三角玻璃棒均勻涂布于平板上;(4)在37℃恒溫培養(yǎng)箱中孵育24h,然后使用吸管從平板上取單個(gè)菌,移到WA平板的中心排布;(5)在37℃恒溫培養(yǎng)箱中孵育4~5天,孵育24h時(shí),開始測量菌落直徑。
1.2.2 Real-time PCR法檢測自噬相關(guān)基因的表達(dá)
將WT和ΔPkaR菌株的孢子以1.0×107個(gè)/mL濃度分別接種于AMM液體培養(yǎng)基,培養(yǎng)8.5~9h,然后轉(zhuǎn)移到1.5mLEP管中離心棄上清,將菌體沉淀用滅菌過的濾紙進(jìn)行進(jìn)一步干燥處理。采用液氮研磨法破碎菌體后加Trizol提取總RNA,用反轉(zhuǎn)錄試劑盒轉(zhuǎn)錄為cDNA,再以cDNA為模板擴(kuò)增。Real-time PCR反應(yīng)體系見表1,反應(yīng)參數(shù):95℃預(yù)變性1min;95℃、15s,56℃、15s,72℃、1min,共40個(gè)循環(huán)。擴(kuò)增反應(yīng)完成后,應(yīng)用軟件設(shè)定閾值,使其處于擴(kuò)增的指數(shù)增長期,并確定閾值循環(huán)數(shù)(Ct),計(jì)算Δ Ct(實(shí)驗(yàn)組內(nèi)參照、目的基因的Ct值減去對照組內(nèi)參照、目的基因的Ct值)和ΔΔ Ct(目的基因的Δ Ct值減去內(nèi)參照基因的Δ Ct值),根據(jù)2-ΔΔCt計(jì)算實(shí)驗(yàn)組目的基因相對于對照組目的基因變化的倍數(shù)。擴(kuò)增引物由上海生工生物工程有限公司合成,引物序列見表2,內(nèi)參為GAPDH。

表1 Real-time PCR反應(yīng)體系
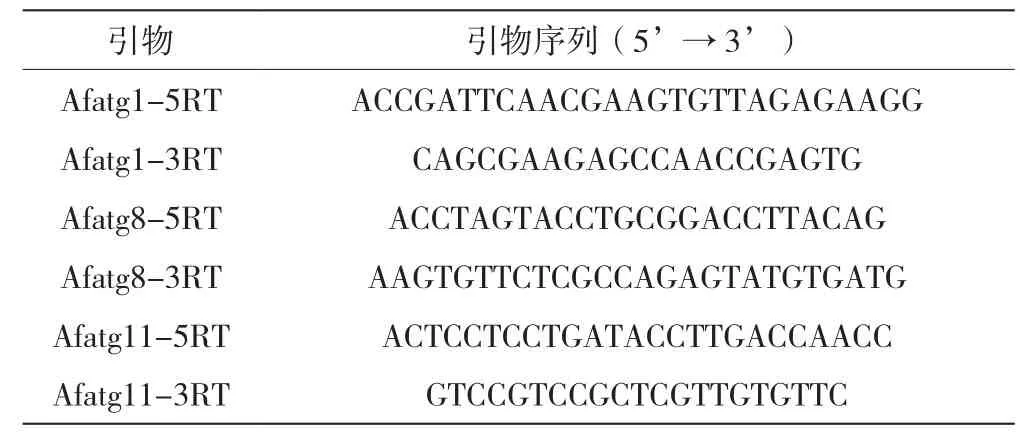
表2 實(shí)驗(yàn)所用引物
2 結(jié)果與分析
2.1 PkaR敲除對煙曲霉菌絲的覓食能力的影響
將已培養(yǎng)24h的WT和ΔPkaR菌的單菌落轉(zhuǎn)移到Y(jié)G和缺乏營養(yǎng)的WA培養(yǎng)基上,24h測一次菌落直徑,結(jié)果如圖1所示。由圖1可知,WT 72h長滿了整個(gè)平板,菌落直徑6.5cm,ΔPkaR 120h達(dá)到最大直徑4.2cm,表明PkaR的敲除影響了頂端菌絲的延伸,抑制了菌絲的伸長。轉(zhuǎn)移到WA培養(yǎng)基上的菌株,WT菌絲生長速度減緩,168 h達(dá)到最大直徑,但ΔPkaR72 h時(shí)停止生長,菌落直徑1.5cm,WA培養(yǎng)基上缺乏營養(yǎng),饑餓可以誘導(dǎo)自噬,表明自噬對煙曲霉菌絲的延伸沒有影響。
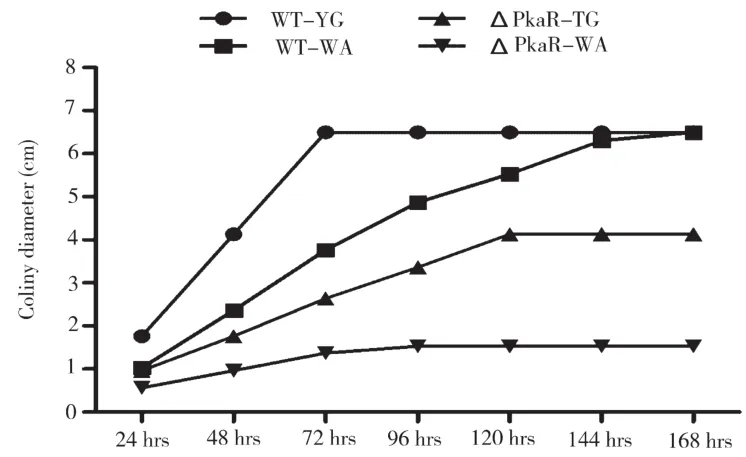
圖1 YG和WA培養(yǎng)基上菌落的變化
2.2 PkaR敲除對煙曲霉Atg1的mRNA水平的影響
對AMM培養(yǎng)基中培養(yǎng)了一段時(shí)間后的WT和ΔPkaR菌株提取RNA,進(jìn)行Real-time PCR定量分析,結(jié)果如圖2所示。由圖2可知,與WT組相比,ΔPkaR組Atg1的mRNA水平顯著上升,差別具有統(tǒng)計(jì)學(xué)意義(P<0.01)。以上結(jié)果表明,PkaR敲除后能增加Atg1的mRNA水平,PkaR可能抑制細(xì)胞自噬。

圖2 PkaR敲除對煙曲霉Atg1的mRNA水平的影響
2.3 PkaR敲除對煙曲霉Atg8的mRNA水平的影響
對AMM培養(yǎng)基中培養(yǎng)了一段時(shí)間后的WT和ΔPkaR菌株提取RNA,進(jìn)行Real-time PCR定量分析,結(jié)果如圖3所示。由圖3可知,與WT組相比,ΔPkaR組Atg8的mRNA水平顯著上升,差別具有統(tǒng)計(jì)學(xué)意義(P<0.01)。以上結(jié)果表明,PkaR敲除后能增加Atg8的mRNA水平,PkaR可能抑制細(xì)胞自噬。
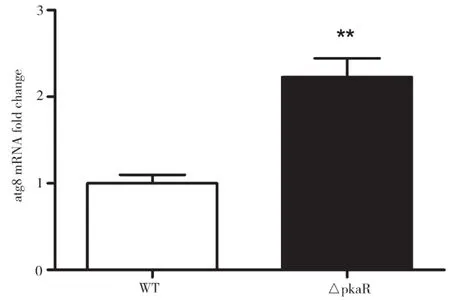
圖3 PkaR敲除對煙曲霉煙曲霉Atg1的mRNA水平的影響。
2.4 PkaR敲除對煙曲霉Atg11的mRNA水平的影響
對AMM培養(yǎng)基中培養(yǎng)了一段時(shí)間后的WT和ΔPkaR菌株提取RNA,進(jìn)行Real-time PCR定量分析,結(jié)果如圖4所示。由圖4可知,與WT組相比,ΔPkaR組Atg11的mRNA水平顯著上升,差別具有統(tǒng)計(jì)學(xué)意義(P<0.01)。以上結(jié)果表明,PkaR敲除后能增加Atg11的mRNA水平,PkaR可能抑制細(xì)胞自噬。
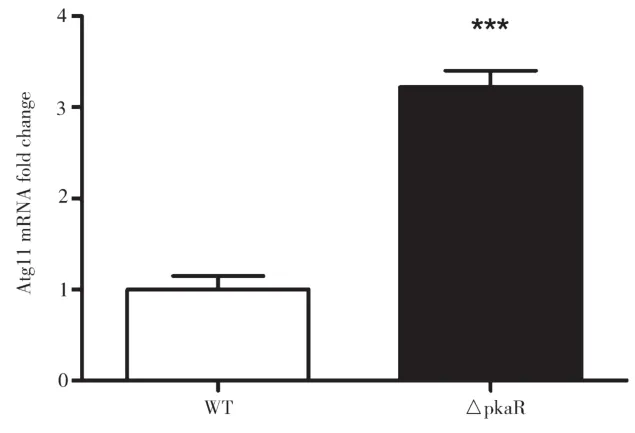
圖4 PkaR敲除對煙曲霉Atg11的mRNA水平的影響
3 結(jié)論
(1)PKA調(diào)節(jié)亞基促進(jìn)煙曲霉的自噬的發(fā)生。(2)PKA調(diào)節(jié)亞基可能是通過促進(jìn)Afatg1、Afatg8和Afatg11的表達(dá)來提高煙曲霉的自噬水平。
煙曲霉是免疫缺陷宿主的重要致病性真菌病原體,普遍存在于人們周邊,其孢子極小,漂浮于空氣中,可隨呼吸進(jìn)入肺中[5]。在水分和足夠的營養(yǎng)物質(zhì)的存在下,如在哺乳動(dòng)物肺中,休眠的分生孢子破壞代謝和細(xì)胞周期的休眠。經(jīng)過一段時(shí)間的核分裂和各向同性生長(膨脹)后,每個(gè)分生孢子建立一個(gè)極軸,然后發(fā)展成侵入性菌絲,引起侵染性曲霉病[6]。因此,分生孢子萌發(fā)代表了煙曲霉生命周期中的關(guān)鍵過程,并且是吸入后,成為侵入性曲霉菌病發(fā)展中的第一個(gè)關(guān)鍵步驟。免疫功能健全的人,吸入的孢子會(huì)被自身免疫清除;而對免疫受損的個(gè)體,吸入的孢子則會(huì)引起侵染性曲霉病或過敏反應(yīng),危及生命[7]。
煙曲霉基因測序完成后,發(fā)現(xiàn)了大量未知功能的基因堿基序列。RNA干擾、基因敲除等基因功能的研究技術(shù)和方法,開始被應(yīng)用于研究未知功能的基因,來試圖找到煙曲霉的關(guān)鍵毒力因子[8]。而自噬是一種通常由饑餓應(yīng)激觸發(fā)的細(xì)胞存活反應(yīng),在真菌中,自噬的報(bào)道相對較少,但真菌的滲透性,菌絲體的生長發(fā)育和它們能侵入不均勻、營養(yǎng)缺乏的物質(zhì)的非凡能力,有力地表明自噬可能是真菌生活方式的基礎(chǔ)。
本實(shí)驗(yàn)證明,自噬的發(fā)生對煙曲霉分生孢子的存活率以及菌絲的生長有影響,但對其侵染性的作用未知。自噬通常被認(rèn)為是促生存的過程,它對于細(xì)胞在細(xì)胞外環(huán)境中應(yīng)對營養(yǎng)不足至關(guān)重要[9]。因此,當(dāng)營養(yǎng)供應(yīng)不足以滿足細(xì)胞能量需求,或者當(dāng)細(xì)胞暴露于不同形式的應(yīng)激時(shí),自噬就會(huì)上調(diào)。在這些條件下,一些研究表明自噬作用可以保護(hù)細(xì)胞免受各種真核生物的死亡。然而,自噬也被證明是細(xì)胞死亡的一個(gè)促成因素,是真核生物中存在一種非凋亡性程序性細(xì)胞死亡途徑,這種途徑取決于自噬基因[10]。在大多數(shù)情況下,這些自噬的雙重作用可能取決于當(dāng)時(shí)的細(xì)胞外環(huán)境條件。已經(jīng)了解TOR激酶在感染相關(guān)自噬的啟動(dòng)中和PKA信號(hào)潛在的相互作用,對其附著胞形態(tài)發(fā)生是必需的,因此,在煙曲霉中了解TOR激酶和PKA信號(hào)的潛在聯(lián)系,以及煙曲霉自噬的基因組分析,對于了解煙曲霉感染至關(guān)重要[11]。如果能驗(yàn)證自噬在煙曲霉侵染過程中的重要性,控制煙曲霉自噬的發(fā)生,可能成為開發(fā)新型抗真菌藥物提供新的靶標(biāo)。
[1]Ohsumi Y. Historical Landmarks of Autophagy Research[J]. Cell Research,2014,24(1):9-23.
[2]Berry A, Han K, Trouillon J, et al. cAMP and Vfr Control Exolysin Expression and Cytotoxicity of Pseudomonas Aeruginosa Taxonomic Outliers[J]. Journal of Bacteriology,2018:18.
[3]Schrevens, S., et al., Methionine is Required for cAMP-PKA-mediated Morphogenesis and Virulence of Candida Albicans[J]. Molecular Microbiology,2018.
[4]Klionsky D, Aghi M, Agam G, et al. Guidelines for the Use and Interpretation of Assays for Monitoring Autophagy (2nd edition)[J].Autophagy,2016,4(1):151-175.
[5]Richie D, Fuller K, Fortwendel J, et al. Unexpected Link between Metal Ion Deficiency and Autophagy inAspergillus fumigatus[J]. Eukaryotic Cell,2007,6(12):2437-2447.
[6]Nadal M, SE Gold. The Autophagy Genes ATG8 and ATG1 Affect Morphogenesis and Pathogenicity in Ustilago maydis[J].Molecular Plant Pathology,2010,11(4):463-478.
[7]Pollack J K, Harris S D, Marten M R. Autophagy in Filamentous Fungi[J].Fungal Genetics & Biology Fg & B,2009,46(1):1-8.
[8]JS Stephan, YY Yeh, V Ramachandran, et al. The Tor and PKA Signaling Pathways Independently Target the Atg1/Atg13 Protein Kinase Complex to Control Autophagy[J]. Proc Natl Acad Sci U S A,2009,106(40):17049-17054.
[9]Feng D, Liu L, Zhu Y, et al. Molecular Signaling Toward Mitophagy and its Physiological Significance[J]. Experimental Cell Resear ch,2013,319(12):1697-1705.
[10]Richie, DL, DS Askew0 Autophagy: A role in metal ion homeostasis?[J].Autophagy,2014,4(1):115-117.
[11]Li X, Cullere X, Nishi H, et al. PKC-δ Activation in Neutrophils Promotes Fungal Clearance[J]. J Leukoc Biol,2016,100(3):581-588.
猜你喜歡
美與時(shí)代·美術(shù)學(xué)刊(2022年3期)2022-04-27 01:18:15
火花(2019年12期)2019-12-26 01:00:28
人大建設(shè)(2019年6期)2019-10-08 08:55:48
人大建設(shè)(2019年12期)2019-05-21 02:55:32
雜文月刊(2018年21期)2019-01-05 05:55:28
人大建設(shè)(2017年6期)2017-09-26 11:50:44
學(xué)苑創(chuàng)造·A版(2015年11期)2016-01-14 09:03:27
俄羅斯問題研究(2012年1期)2012-03-25 09:54:45
中國火炬(2010年12期)2010-07-25 13:26:22
中國火炬(2010年8期)2010-07-25 11:34:30
