Population genetic structure of the mantis shrimpOratosquilla oratoria(Crustacea: Squillidae) in the Yellow Sea and East China Sea*
2018-07-11 01:58:34YANGMei楊梅LIXinzheng李新正
YANG Mei (楊梅) LI Xinzheng (李新正)
1Institute of Oceanology,Chinese Academy of Sciences,Qingdao 266071,China
2University of Chinese Academy of Sciences,Beijing 100049,China
3Laboratory for Marine Biology and Biotechnology,Qingdao National Laboratory for Marine Science and Technology,Qingdao 266000,China
AbstractThe mantis shrimpOratosquilla oratoriais an ecologically and economically important species in the Western Pacific. In present study, the population genetic structure ofOratosquilla oratoria from the Yellow Sea and East China Sea was examined with mitochondrial DNA control region sequences. In total,394 samples were collected from 18 locations and 102 haplotypes were obtained. For the Yellow Sea, the overall nucleotide diversity and haplotype diversity were 0.006 9 and 0.946 8, respectively; while across all the East China Sea locations, the overall nucleotide diversity and haplotype diversity were 0.027 94 and 0.979 0, respectively. The results of AMOVA and pairwiseF ST(0.145 2,P<0.001) revealed moderate differentiation between the Yellow Sea and East China Sea populations ofO. oratoria. However, neither the neighbor-joining tree nor haplotype network showed clades with geographic pattern, which indicated considerable gene flow was existed between the Yellow Sea and East China Sea, and supporting the high larval dispersal ability in this species. Mismatch distribution analysis and neutrality tests suggested thatO.oratoriahas undergone population expansion event, and the Pleistocene glacial cycles might have an impact on the historical demography ofO. oratoria. The genetic information obtained in this study can provide useful information for sustainable improvements for capture fi sheries management strategies.
Keyword: Oratosquilla oratoria; genetic structure; mtDNA control region; Yellow Sea; East China Sea
1 INTRODUCTION
In Northwestern Pacific, the marginal seas of China has well-characterized oceanography and dramatic geological history, which provides one of the best model system for marine organisms population genetics research (Kong and Li, 2009). Recent studies revealed that the historical separation of the sea basins during the Pleistocene glaciers, coastal topography,ocean currents and freshwater outf l ow played discrete but important inf l uences in shaping the present genetic structure of various marine species (Liu et al., 2007;Shen et al., 2011; Dong et al., 2012; Hurwood et al.,2014).
Oratosquillaoratoriais widely distributed in the Western Pacific, and has been regarded as one of the valuable fishery resources for its good taste and rich nutrition (Wang et al., 1996). In recent years, the natural resources ofO.oratoriahave been decreased rapidly due to overfishing, marine pollution and habitat destruction. Inevitably, the decline in wild resource will reduce the intraSpecific genetic diversity and weakening the understanding of population genetics to the species (Lu et al., 2013). With stable maternal inheritance and a generally high rate of base substitutions, mitochondrial DNA (mtDNA) markers have been widely used for studies ofintraSpecific population genetics (de Bruyn et al., 2005; Alam et al., 2015). For example, mtDNA variation inPenaeussemisulcatusreveals two distinct lineages in the Indo-West Pacific region (Alam et al., 2017). And due to high commercial and ecological values of theO.oratoria, there were several researches to estimate the genetic diversity and genetic structure in China Sea (Lui et al., 2010; Zhang et al., 2012, 2016; Du et al., 2016). In present study, based on the analysis of mtDNA control region sequences, we surveyed the population genetics ofO.oratoriain two marginal seas (i.e., the Yellow Sea and East China Sea) by intensive sampling. And we aimed to gain complementary information about the present situation of genetic diversity as well as evolutionary patterns ofO.oratoria.
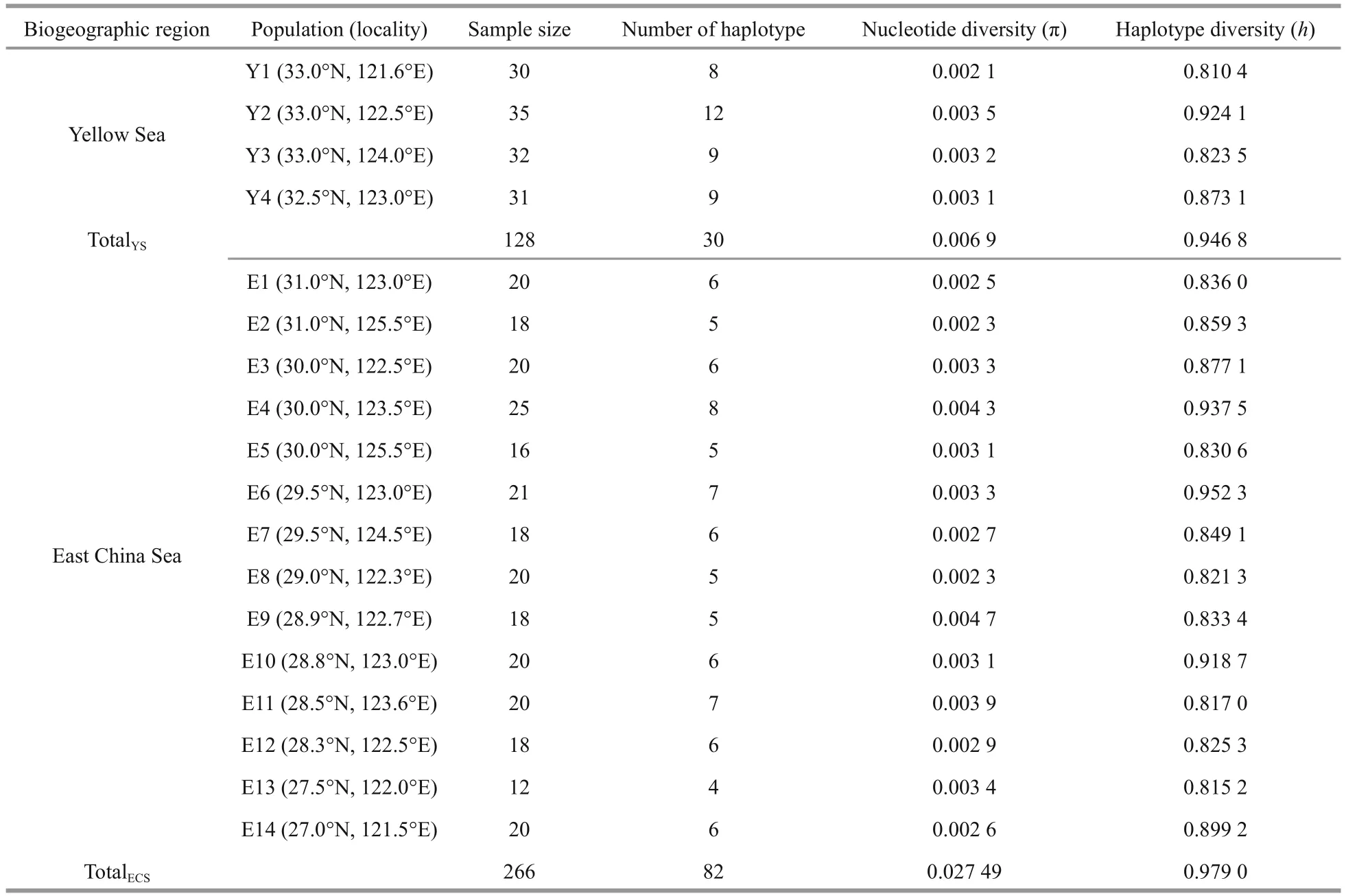
Table 1 Sampling information and genetic diversity index of Oratosquilla oratoria from the Yellow Sea and East China Sea
2 MATERIAL AND METHOD
2.1 Sample collection
All the specimens of wildO.oratoriawere collected between May 2014 and November 2015.We obtained 394 individuals of this species from 18 locations in the Yellow Sea and East China Sea, with a range of 12-35 individuals per site (Fig.1 and Table 1). The materials were sampled and preserved in 95%ethanol for subsequent DNA extraction.
2.2 DNA extraction, amplifi cation, and sequencing
Genomic DNA was extracted from muscle tissue by the Tissue DNA Kit (OMEGA). For each individual, fragments of mtDNA control region were amplified with primer pairs: CR-F1(5′-TAACCGCGACTGCTGGCAC-3′), CR-F2(5′-GGGTATGAGCCCATTAGCTT-3′) (Lui et al.,2010). PCR amplifi cations were carried out in 25 μL volume containing 12.5 μL 2X Premix Ex Taq(TaKaRa Co., China), 1 μL of each primer, 1 μL template DNA and 9.5 μL ddH2O. The cycling conditions were as follows: initial denaturation at 94°C for 2 min, followed by 30 cycles of 94°C for 1 min, 56°C for 45 s, 72°C for 50 s, then a fi nal extension at 72°C for 10 min. PCR products were purified using the PCR clean-up Kit (Axygen) and sequenced on ABI 3730 automatic sequencer (Sangon Inc., China).
2.3 Data analyses

Fig.1 Sampling locations of Oratosquilla oratoriaYellow Sea: Y1-Y4; East China Sea: E1-E14. The red line indicates the path of the Kuroshio Current and its tributaries in the East China Sea,N-KBBC: an nearshore Kuroshio bottom branch current; O-KBBC: an offshore Kuroshio bottom branch current.
Sequences were aligned with ClustalX (Thompson et al., 1997) using default settings and corrected by hand. The nucleotide diversity (π) and haplotype diversity (h) were calculated using the DnaSP(Librado and Rozas, 2009). Phylogenies of haplotypes were reconstructed by the neighbor-joining (NJ)(Saitou and Nei, 1987) (Bootstrap=1 000) in MEGA 6.0. In addition, a median-joining (MJ) was generated for haplotypes using the Network (Bandelt et al.,1999). Population genetic structure was evaluated withFSTstatistics and AMOVA (analysis of molecular variance) (Excoffer et al., 1992) in ARLEQUIN(Excoffer and Lischer, 2010). The Tajima’sD(Tajima, 1989) statistics, Fu’sFS(Fu, 1997) test of neutrality and the frequency distribution of pairwise differences between mtDNA haplotypes (i.e.,mismatch distribution) were conducted to infer the population demographic history ofO.oratoria. The tau value (τ), which reflects the location of the mismatch distribution crest, provides a rough estimate of the time when rapid population range expansion started. According tot=τ/2u, calculate the expansion time ofO.oratoriain the Yellow Sea and East China Sea, respectively. And the mutation rate of 3.44%/Myr for Mitochondrial control region was applied to in the present study (Alam, 2016).
3 RESULT
3.1 Genetic diversity
A 923-bp segment of mtDNA control region was amplified and total of 102 different haplotypes were identified based on the sequence variation in 394 individuals from 18 locations of the Yellow Sea and East China Sea. There were 30 haplotypes of 128 individuals detected in the Yellow Sea, while 82 haplotypes of 266 individuals in the East China Sea(Table 1). And among these, 10 of the haplotypes were shared. The most common haplotype, Hap6,was shared by a total of 62 individuals from the Yellow Sea and East China Sea.
The estimates of nucleotide diversity (π) and haplotype diversity (h) for different locations were showed in Table 1. The highest value of π was in E9(0.004 7) of the East China Sea and the lowest was in Y1 (0.002 1) of the Yellow Sea; the highest value of h was in E6 (0.952 3) of the East China Sea and the lowest was in Y1 (0.810 4) of the Yellow Sea. For the Yellow Sea, the overall nucleotide diversity and haplotype diversity were 0.006 9 and 0.946 8,respectively; while across all the East China Sea locations, the overall nucleotide diversity and haplotype diversity were 0.027 94 and 0.979 0,respectively.
3.2 Population structure
Genetic differentiation among eighteen different locations was evaluated usingFSTvalues and AMOVA analyses. In general, the pairwiseFSTwere range from-0.032 3 to 0.132 4 (Table 2), the largest genetic differentiation occurred between Y3 and E12(FST=0.132 4,P<0.001). The pairwiseFSTvalues between the Yellow Sea (Y1-Y4) and East China Sea(E1-E14) populations was 0.145 2 (P<0.001),indicated a moderate degree of differentiation was existed. Furthermore, we did an analysis of genetic differentiation between the Kuroshio tributary and other populations in the East China Sea. According to previous studies about Kuroshio in the East China Sea(Yang, 2011), we considered the individuals from E4,E6, E9, E10 and E12 as a population in Kuroshio tributary (N-KBBC: a nearshore Kuroshio bottom branch current), while remaining individuals (E1-E3,E5, E7-E8, E11, E13-E14) as the other population(Fig.1). The result showed that differentiation existed between above populations (FST=0.0495,P<0.001).
One-group AMOVA analysis showed that theoverall population differentiation was 14.46% and genetic variation within populations was 85.54%(P<0.001). Two groups AMOVA (Yellow Sea and East China Sea) showed that the source of genetic differentiation was mainly found within populations(89.5%), while the rest was distributed among populations within groups (7.2%) and among groups(3.3%).
The median-joining network ofO.oratoriaproved that the haplotypes could not be clearly divided into different geographical groups, and was characterized by a star-like phylogeny with closely related haplotypes derived from the most common and ancestral haplotype (Fig.2). Moreover, the topology of the NJ tree was shallow and displayed no significant genealogical branches of samples corresponding to sampling locations (data not presented).
3.3 Historical demography
The mismatch distribution ofO. oratoriafor the Yellow Sea and East China Sea were consisted of a distinct unimodal curve, which closely fi tted to the expected distributions under a rapid population expansion model (Fig.3). In addition, both Tajima’sDstatistics (Tajima’sDYS=-1.94,P<0.001; Tajima’sDECS=-2.29,P<0.001) and Fu’sFStest (FS-YS=-6.05;FS-ECS=-11.79) were all showed significant negative values. All of these results suggested that theO.oratoriahad undergone a recent colonization and sudden population expansion events in the Yellow Sea and East China Sea. And the estimate of time of expansion for the Yellow Sea was 176 800 years,while for the East China Sea, this estimate was 206 600 years ago.
4 DISCUSSION
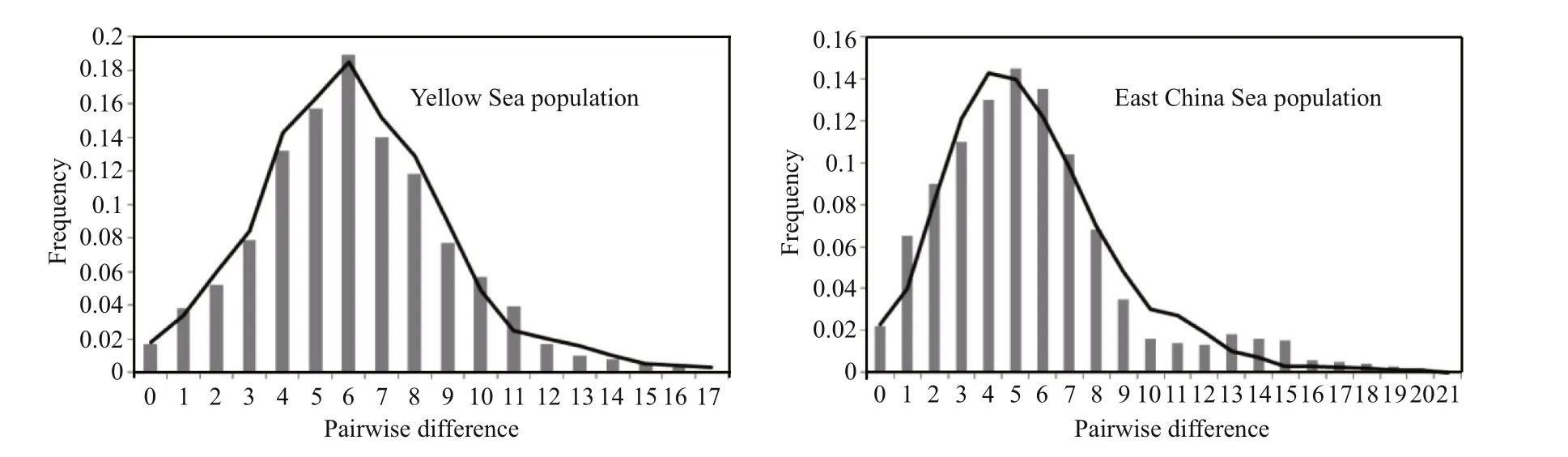
Fig.3 Observed pairwise differences (bars), and the expected mismatch distributions under the sudden expansion model(line) for the control region in Oratosquilla oratoria
Generally, due to high dispersal potential(planktonic egg, larval or adult history stages) and absence of physical barriers, marine organisms are expected to have low degree of genetic differentiation among geographical regions (Palumbi, 1994; Hewitt,2000). However, in present study, the results of pairwiseFSTand AMOVA analyses revealed moderate population structure ofO.oratoriabetween the Yellow Sea and East China Sea populations. But neither the neighbor-joining tree nor haplotype network showed clades with geographic pattern,which indicated effective gene flow was existed between these marginal seas. Considering theO.oratoriahas a long planktonic larval phase of between 36 to 59 days (Hamano and Matsuura, 1987), larval dispersal driven by oceanographic circulation might play important role in maintenance of population connectivity. In the Yellow Sea and East China Sea,the composition of ocean currents circulation is complex, including China Coastal Current,Changjiang (Yangtze) River outf l ow, Kuroshio tributary and so on (Li et al., 2000). Previous studies have been confi rmed that the China Coastal Current transport certain fl oating marine organisms from the Yellow Sea to the East/South China Sea, such as fragments of Sargassum andCalanussinicus(Hwang and Wong, 2005; Cheang et al., 2010). Our results suggested that planktonic larvae ofO.oratoriain the Yellow Sea and East China Sea might travel certain distances with ocean currents, yielding gene flow among different geographical regions.
Besides ocean currents, the dispersal of larval can also be affected by freshwater runoff from the Changjiang River. As a euryhaline species,O.oratoriaappears to tolerance wide raged salinity (20-40) but sensitive to low salinity (lower than 15) (Liu et al., 2006, 2012). Meanwhile, the peak spawing period (May to July) ofO.oratoriawas consist with the large discharge of Changjiang River in summer,which greatly reduced the seawater salinity in the estuary area (Wang et al., 1998). It is likely that, to some extent, the Changjiang River form a physical barrier, which acted to prevent transport of larval or adults ofO.oratoriaamong different biogeographic regions. And the barrier effect of the Changjiang River outf l ow have been ascertained by relevant studies (Du et al., 2016). Additionally, in recent years there were several studies have focused on population genetic structure of marine species between the Yellow Sea and East China Sea (Ni et al., 2014).Genetic studies onNibeaalbif l ora,Portunus trituberculatus,CyclinasinensisandCharybdis bimaculatafound no significant genetic structure between the Yellow Sea and East China Sea populations, showed the Changjiang River outf l ow was not validated for these species because their prejuveniles and juveniles are able to survive over a wide range of salinities (Feng et al., 2008; Han et al.,2008, 2015a; Ni et al., 2012). However, similar researches on limpetCellanatoreumaand cocktail shrimpTrachypenaeuscurvirostrisindicated that significant genetic differentiation in line with the outf l ow, which limited the dispersal of larvae along the Chinese coast (Dong et al., 2012; Han et al.,2015b). These results revealed that the Changjiang River outf l ow affect some species depending on habitat Specificity and differentiation in biological characteristics (such as salinity tolerance and dispersal ability) (Ni et al., 2014).
Furthermore, in our study, differentiation was detected between the Kuroshio tributary and other regions populations of the East China Sea. The Kuroshio Current originates in the westward-f l owing North Equatorial Current of the central Pacific then def l ects towards the northeast offshore of the Philippine Islands (Dou et al., 2012), and producing striking inf l uence on biodiversity and genetic structure of marine species in the Northwestern Pacific (Yasuda et al., 2009; Nakajima et al., 2014). Along the coastline of Taiwan, the Kuroshio Current flows south to north at an average rate of 25 cm/s (Ju et al., 2013) and can carry larval ofO.oratoriafrom the South China Sea to the East China Sea. Previous studies have showed the formation of Taiwan limited the gene flow ofO.oratoriabetween the East China Sea and South China Sea (Zhang et al., 2014, 2016). Additionally, ocean currents of the East China Sea exhibited characteristics of counterclock wise cyclonic circulation, while the surface circulation of the South China Sea has clockwise circulation due to the monsoon drift, the opposite ocean currents driving the planktonic larvae ofO.oratoriain opposite directions during the reproductive season further deepened their divergence(Zhang et al., 2014, 2016).
Evidence for the occurrence of population expansion ofO.oratoriain the Yellow Sea and East China Sea were confi rmed by the mismatch distribution analyses and neutrality tests (Fig.3).Based on values of the nucleotide and haplotype diversity of mtDNA, four basic scenarios of population history were proposed by Grant and Bowen (1998).Oratosquillaoratoriawith low nucleotide diversity and high haplotype diversity probably undergone sudden population expansion after a period of low effective population size. Once population size increases rapidly following environmental disturbances, haplotype diversity can usually recover via mutation, whereas nucleotide diversity remains low since there was not enough time to accumulate large sequence differences between haplotypes (Avise, 2000; Lui et al., 2010).According to previous phylogeographic studies, the population sudden expansion of marine organisms was common in the Northwestern Pacific (Ni et al.,2014). During the Pleistocene glacial cycles, as one of the most extensive continental shelves, the Yellow Sea and most of the East China Sea were exposed,and the latter was consequently reduced into an elongated enclosed sea with an area <1/3 ofits present size (Wang, 1999; Kitamura et al., 2001). TheO.oratoriain these marginal seas can survived only in common glacial refuge regions, and in postglacial,when sea level rose as glaciers melted, the Yellow Sea and East China Sea must have been recolonized byO.oratoria. Furthermore, our studies revealed a shallow and star-like topology, which suggested an origin from a common ancestral population in the East China Sea refugium.
5 CONCLUSION
In the present study,O.oratoriashowed moderate degree of genetic differentiation between the Yellow Sea and East China Sea populations. The Changjiang River acted as a physical barrier to prevent transport of larval or adults ofO.oratoriabetween these marginal seas, while the considerable gene flow was maintained due to the larval dispersal with ocean currents. And based on our results, theO.oratorianeeds cautious to deal different stocks in the Yellow Sea and East China Sea. Further studies ofO.oratoriathroughout the whole Western Pacific and employ more molecular markers would contribution to enhancing the knowledge of the genetic structure and evolutionary history, thus provides theory basis and instruction for conserving wild germplasm resources.
6 DATA AVAILABILITY STATEMENT
All data generated or analyzed during this study are included in this published article.
7 ACKNOWLEDGEMENT
We are very much indebted to DONG Dong, MA Lin, SUI Jixing, KOU Qi, GAN Zhibin, GONG Lin,WANG Jinbao, WANG Yueyun, XU Yong and SUN Yue for their work in the fi eld and laboratory.
登錄APP查看全文
 Journal of Oceanology and Limnology
2018年3期
Journal of Oceanology and Limnology
2018年3期
- Journal of Oceanology and Limnology的其它文章
- Editorial Statement
- The post-larval and juvenile fi sh assemblage in the Sukhothai Floodplain, Thailand*
- Effects of probiotic on microf l oral structure of live feed used in larval breeding of turbotScophthalmus maximus*
- Comparison ofintestinal microbiota and activities of digestive and immune-related enzymes of sea cucumberApostichopus japonicusin two habitats*
- Otolith shape analysis for stock discrimination of twoCollichthysgenus croaker (Pieces: Sciaenidae,) from the northern Chinese coast*
- The impact of spatial autocorrelation on CPUE standardization between two different fi sheries*