四倍體馬鈴薯塊莖蛋白含量分子標記的開發與驗證
2018-07-13 02:41:58單洪波史佳文
作物學報 2018年7期
關鍵詞:檢測
單洪波 史佳文 石 瑛
東北農業大學農學院, 黑龍江哈爾濱 150030
蛋白含量是馬鈴薯塊莖的重要品質性狀, 也是重要的育種目標之一[1]。張澤生等[2]對馬鈴薯蛋白的氨基酸組成深入研究證實, 塊莖內含有較高的必需氨基酸, 占其總量的47.90%左右。潘牧等[3]研究表明, 馬鈴薯蛋白的第一限制氨基酸為色氨酸, 且富含其他糧食作物所缺乏的賴氨酸, 可與大豆蛋白等谷類蛋白互補。通過對動物與人類試驗證明, 馬鈴薯蛋白富有較高的生物價值[4-5]。因此, 馬鈴薯塊莖蛋白是一種優質蛋白, 營養價值極高, 高蛋白含量馬鈴薯品種的選育能夠提升馬鈴薯的營養價值。
傳統的馬鈴薯育種周期較長, 篩選效率較低, 并伴隨精確度低等諸多弊端, 分子標記輔助選擇能極大地縮短育種年限, 提高育種效率。目前, 已有數十個馬鈴薯重要性狀被標記, 例如抗病毒病、抗青枯病、抗晚疫病等抗病性狀[6-7]; 炸片顏色、薯肉顏色等品質性狀[8]; 耐干旱、抗凍性等抗性性狀[9]。祁緣[10]通過BSA策略結合與QTL區域緊密連鎖的分子標記DMC42144af、DMC42152bf, 將2個馬鈴薯晚疫病 AFLP標記定位在第 9染色體上的一個QTL dPI09a置信區間內; 雷劍等[11]根據 BSA混池方法開發了與馬鈴薯青枯病抗性緊密連鎖的SRAP標記; Rak等[12]利用多倍體模型全基因組關聯分析將薯片顏色 QTL定位到第2染色體的短臂上; Tiwari等[13]將源于二倍體馬鈴薯中的BR基因分子標記用于馬鈴薯抗晚疫病性狀的篩選, 但在四倍體馬鈴薯材料的實際應用中, 復雜的遺傳方式使目的性狀與標記分離, 并伴隨篩選準確率低等諸多問題。有關塊莖蛋白含量的分子標記研究較少, 鄔信康等[14]根據限制性片段長度多態性分析(RFLP)和脈沖電泳分析, 表明馬鈴薯蛋白多基因簇位于第 8染色體的 1.4 Mb的DNA片段上。隨著BSA混池方法與測序技術的不斷成熟, 高篩選效率與精準度得到了極大的保障, 能夠為分子輔助育種提供新的標記資源。在四倍體水平上對馬鈴薯蛋白含量相關分子標記的開發與研究有利于加速馬鈴薯蛋白含量分子輔助育種進程。
本研究以四倍體馬鈴薯蛋白含量分離群體為材料,利用BSA混池構建與高通量簡化基因組測序相結合(以下簡稱BSA混池測序), 開發四倍體馬鈴薯蛋白含量的分子標記, 并經極端高蛋白、極端低蛋白群體材料及四倍體馬鈴薯品種的驗證, 獲得與四倍體馬鈴薯蛋白質含量相關的分子標記, 為分子輔助育種提供便利, 也為加快優質馬鈴薯品種的選育提供技術支撐。
1 材料與方法
1.1 試驗材料
1.1.1 F1代分離群體 以蛋白含量 2.32%的高蛋白品種大西洋為母本, 蛋白含量1.31%的低蛋白品種定薯1號為父本, 雜交獲得 173份 F1代分離群體, 以親本和 F1代分離群體作為開發蛋白含量分子標記的試驗材料。
1.1.2 供試品種 選取54個四倍體馬鈴薯品種作為蛋白含量分子標記的驗證材料(表1)。

表1 馬鈴薯品種信息Table1 Information of potato varietie s
1.2 分離群體及四倍體品種蛋白含量的測定
2016—2017年連續 2年將分離群體種植于東北農業大學向陽農場實驗基地, 每份材料種植7株, 于收獲期收獲材料, 選取每份材料3株進行樣品處理。將粉碎并通過40~60目/寸網篩的試樣0.1 g置洗滌干凈且烘干的消化管中, 加入1 mL水和5 mL濃硫酸放置過夜。
將消化管放入通風櫥中的消煮爐上, 設置溫度為400~450℃, 每隔 15 min加3~4滴催化劑過氧化氫。待樣品消化至澄清透明且無雜質, 取下放涼, 同時做空白試驗。
將消化好的樣品經凱氏定氮儀檢測, 得到全氮含量,換算成蛋白含量。
1.3 DNA的提取與極端分離后代DNA混池的構建
采用 CTAB法提取馬鈴薯材料基因組 DNA, 用瓊脂糖凝膠電泳檢測DNA質量。
分別構建極端高蛋白含量和極端低蛋白含量后代的DNA混池, DNA的提取方法與質量檢測方法同上。
1.4 標記開發
建庫、簡化基因組測序由上海歐易生物醫學科技有限公司幫助完成, 根據測序結果獲得了極端高蛋白與極端低蛋白混池間的差異片段, 參照馬鈴薯基因組序列網站(http://solanaceae.plantbiology.msuedu/pgsc_download.sht ml)對差異片段內的差異標簽的位點利用在線引物設計網站(http://www.yeastgenome.org/cgi-bin/web-primer)進行引物的設計。將Tm值設定在55℃~60℃之間, 引物預擴增片段大小設定在400~900 bp之間。將設計好的引物送至博仕生物工程有限公司進行合成。
以高蛋白馬鈴薯品種大西洋和低蛋白馬鈴薯品種定薯1號的基因組DNA為模板進行PCR擴增, 選擇僅能在親本之一中擴增出條帶或者在雙親中擴增出不同條帶的引物用于開發SCAR標記。最后對極端分離群體材料進行標記驗證。通過1.2%瓊脂糖凝膠電泳對PCR產物進行質量與特異性檢測。PCR體系為20 μL, 含8.2 μL ddH2O、10 μLTaq-Plus PCR Forest Mix (2×)、0.4 μL 引物 F (10 μmol L–1)、0.4 μL 引物 R (10 μmol L–1)、1 μL DNA (100 ng μL–1)。PCR擴增條件設定為預變性95℃ 3 min; 變性95℃ 30 s, 退火58℃ 30 s, 延伸72℃ 50 s, 35個循環; 終延伸72℃ 10 min。
1.5 標記的檢測
分別以馬鈴薯F1代分離群體的74份高蛋白群體材料、42份低蛋白群體材料及54個四倍體馬鈴薯品種的基因組DNA作為模板, 以分子標記SCAR8-107為引物進行PCR擴增后利用瓊脂糖凝膠電泳進行PCR產物的檢測。
1.6 數據統計和處理
根據分子標記檢驗結果, 將與高蛋白含量親本和低蛋白含量親本帶型一致的基因型分別記作1和0, 通過統計分析軟件SPSS的Pearson’s雙側檢驗對分子標記檢測結果與蛋白含量的鑒定結果進行雙側相關性分析。
2 結果與分析
2.1 蛋白含量的測定
測量獲得蛋白含量大于2.0%的高蛋白材料74份, 蛋白含量小于1.5%的低蛋白材料42份以及介于1.5%~2.0%之間的中蛋白含量材料57份(表2); 經對分離群體蛋白含量的測定得到了分離群體的蛋白含量分布圖, 可以看出極端材料較少, 中等蛋白含量材料較多, 并且曲線大致呈正態分布(圖1)。說明馬鈴薯蛋白含量為數量性狀。
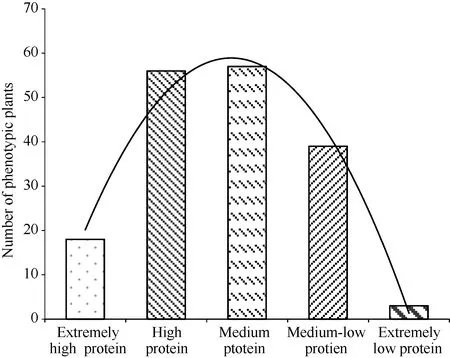
圖1 群體蛋白含量分布Fig. 1 Distribution of group protein content
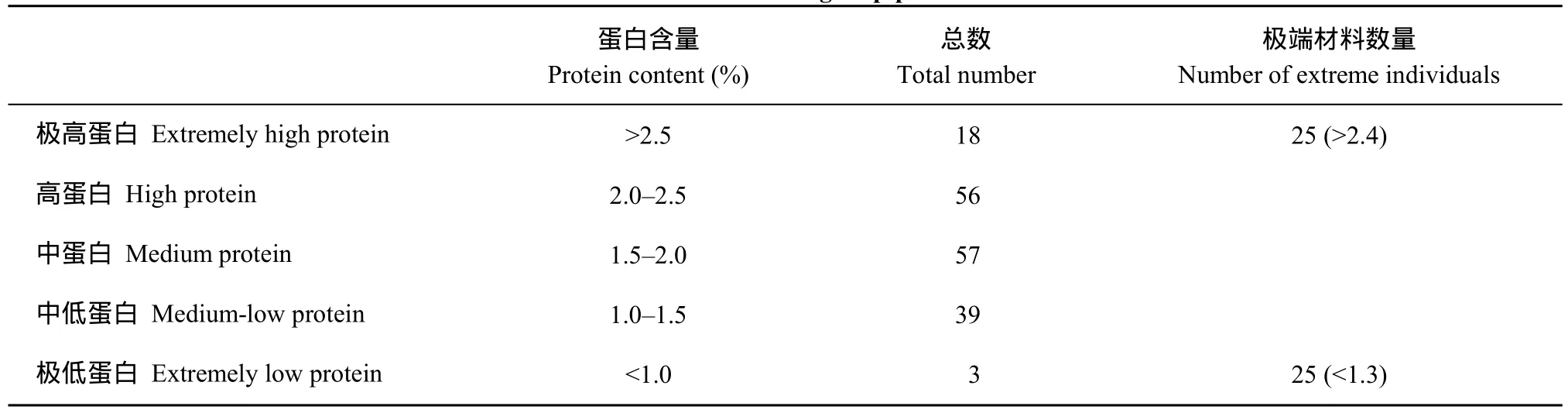
表2 群體蛋白含量的測定結果Table 2 Determination of group protein content
2.2 蛋白含量基因區段引物的設計與檢測
根據前期測序的結果, 在第 8染色體上 1.151~1.248 Mb區間內得到了目的性狀的差異片段, 此片段與控制馬鈴薯高蛋白含量相關聯, 區間大小在 97 kb,在該區段共設計引物34對(表3), 分別以高蛋白親本品種大西洋與低蛋白親本品種定薯1號的基因組DNA為模板對設計的引物進行特異性檢測(圖 2)。在該區段設計的引物中, 引物8-107展現出在差異蛋白含量親本中的特異性, 即在高蛋白含量親本擴增有條帶, 低蛋白親本無擴增條帶。

表3 引物信息Table 3 Primer information
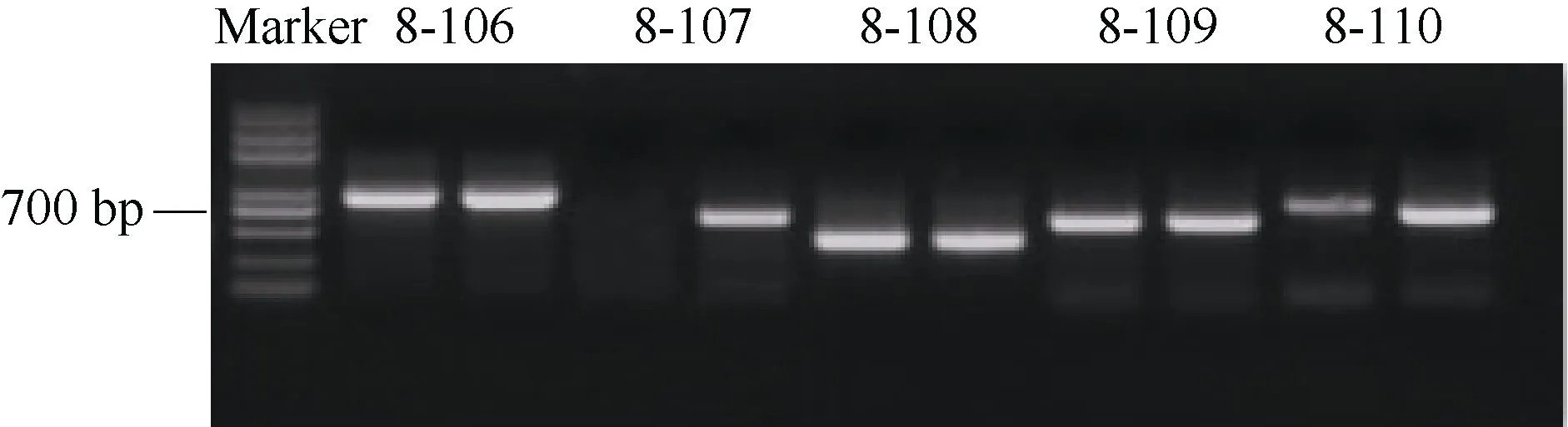
圖2 部分引物在親本定薯1號和大西洋的擴增產物電泳圖Fig. 2 PCR products of some primers in parents Dingshu 1 and Atlantic
2.3 分子標記的開發與驗證
將在雙親中擴增出差異條帶的引物, 通過 25份極端高蛋白與 25份極端低蛋白后代驗證發現, 親本擴增出現差異的引物8-107在極端高蛋白與極端低蛋白后代中擴增結果一致。即引物8-107在極端高蛋白材料中的擴增結果與親本大西洋一致, 均擴增出條帶, 而引物 8-107在極端低蛋白材料中的擴增結果與親本定薯1號保持一致, 均未擴增出條帶(圖3)。因而, 本試驗中, 篩選的引物8-107被選取用于后續的實驗研究, 命名引物 8-107為SCAR8-107。
2.4 標記SCAR8-107在分離群體高蛋白和低蛋白子代中的檢測
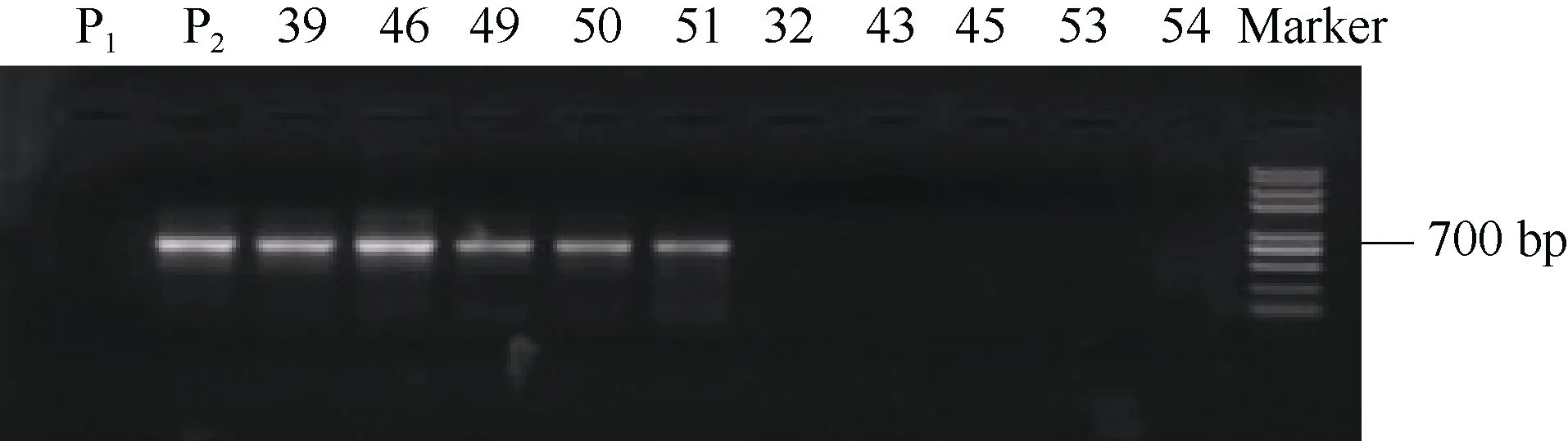
圖3 引物8-107標記在部分極端高蛋白與極端低蛋白分離后代中的PCR擴增結果Fig. 3 PCR amplification results of primers 8-107 in partial extremely high protein and extremely low protein progenies
在四倍體馬鈴薯F1代的173份分離群體中, 存在74份高蛋白材料、42份低蛋白材料以及 57份中蛋白材料,以其中的74份高蛋白群體材料和42份低蛋白群體材料作為SCAR8-107標記檢測的目的材料。在標記為陽性的(有PCR擴增產物)的70份材料中有67份為高蛋白含量材料,分子標記的檢測結果與表型的對應度達到了 95.71%。在標記為陰性的(無PCR擴增產物)的46份材料中有38份為低蛋白材料, 分子標記的檢測結果與表型的對應度達到了82.61%。共116份后代材料的分子標記與蛋白含量表型鑒定的結果總體的對應度達到了90.51% (圖4和表4)。將蛋白含量的測定結果與分子標記的鑒定結果進行SPSS雙側相關性分析, 相關系數(r)為0.794, 達到了極顯著水平。
2.5 標記SCAR8-107在四倍體品種中的檢測
在54個馬鈴薯品種中, 蛋白含量大于2.0%的高蛋白含量的品種有10個, 占總材料的18.51%; 蛋白含量小于1.5%的低蛋白含量品種有20個, 占總材料的37.03% (表1)。利用開發的分子標記SCAR8-107對54個馬鈴薯品種檢測表明, 在標記為陽性的14個品種中, 有9個為高蛋白含量, 分子標記的檢測結果與蛋白含量的對應度達到了71.42%; 在標記為陰性的23個品種中, 有17個為低蛋白含量, 分子標記的檢測結果與蛋白含量的對應度達到了73.91% (圖5和表5)。54個品種的標記檢測結果和蛋白含量的測定結果的總體的對應度達到了 72.97%。將分子標記鑒定結果與蛋白含量測定結果進行SPSS雙側相關性分析, 相關系數(r)為0.694, 達到極顯著水平。結果表明, 分子標記SCAR8-107能夠很好地篩選不同馬鈴薯品種的蛋白含量, 此標記能應用于馬鈴薯蛋白含量的分子標記輔助育種。

圖4 SCAR8-107標記在部分高蛋白(A)與低蛋白(B)分離后代中的PCR擴增結果Fig. 4 PCR amplification results of SCAR8-107 marks in partial high protein (A) and low protein (B) progenies
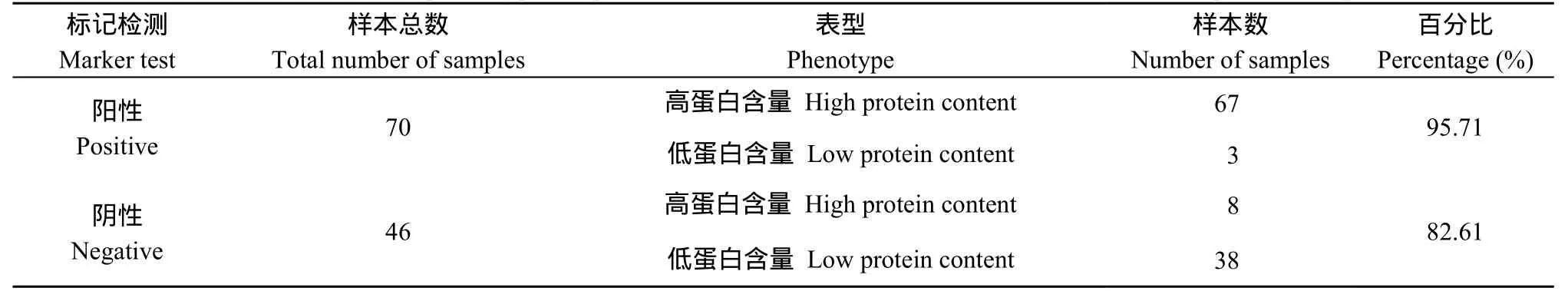
表4 F1群體部分樣品的蛋白含量表型鑒定與標記檢測結果統計Table 4 Identification of phenotype for protein content and statistics of marker test results in partial samples of F1
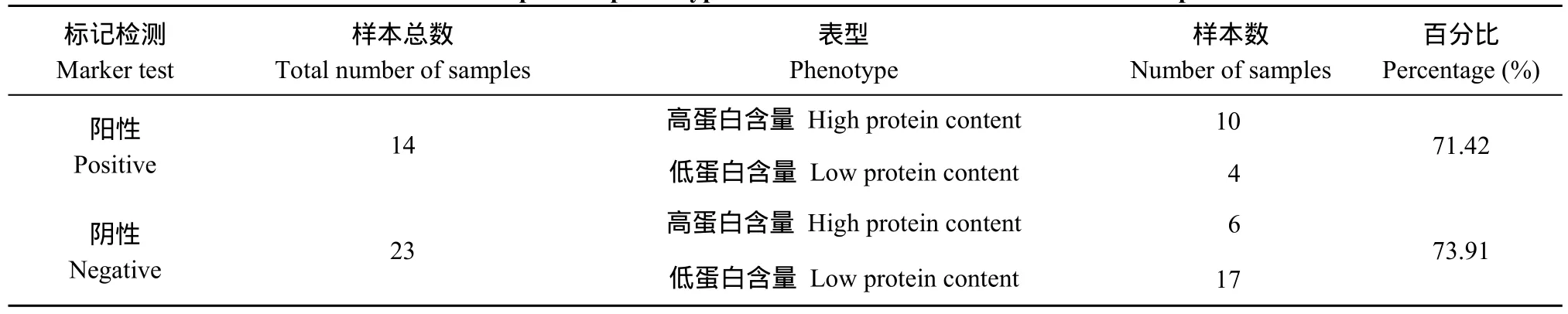
表5 部分品種蛋白含量表型鑒定與標記檢測結果統計Table 5 Identification of protein phenotype and statistics of marker test results in partial varieties
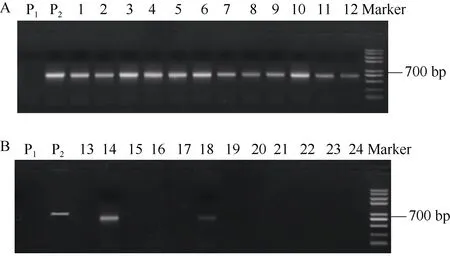
圖5 SCAR8-107標記在部分高蛋白(A)與低蛋白(B)品種中的PCR擴增結果Fig. 5 PCR amplification results of SCAR8-107 marks in partial high protein (A) and low protein (B) varieties
3 討論
開發并獲得與目的基因緊密連鎖的分子標記是分子標記輔助育種的重要基礎, 隨著 AFLP、SSR、SRAP、RAPD等技術的發展, 分子標記輔助育種的研究進程也得到了極大的推動, Takagi等[15]采用當地品種“Hitomebore”進行 EMS誘變處理, 產生耐鹽性的突變體(hst1), 運用BSA策略與野生親本進行全基因組重測序, 結合Mutmap分析法, 找到了水稻耐鹽基因(OSRR22), 僅用2年時間就培育出耐鹽性的水稻新品種; 路洪鳳等[16]采用 BSA混樣策略, 通過全基因組掃描SNP, 分析差異頻率, 檢測F2群體早花性狀的QTL, 找到了一個位于早花的QTL Ef1.1中的候選基因。由于四倍體馬鈴薯減數分裂的復雜性以及分子標記輔助育種復雜程度與通量的雙重限制, 導致難以應用于規模較大的分子標記輔助育種。本研究利用 BSA混池測序的方法對四倍體馬鈴薯進行了 SCAR標記的開發與驗證。該技術能夠極大地避免片段大小選擇的過程,使標簽在基因組中的分布更加均勻, 適用于四倍體馬鈴薯等遺傳性多樣、基因組復雜的染色體片段的挖掘與分子標記的開發。BSA (Bluk Segregant Analysis)集群分離分析法是一種利用極端性狀進行功能基因定位的方法, 比較群體在多態位點(SNP)的等位基因頻率(AF)是否具有顯著差異。因此, BSA混池測序的方法在技術層面上解決了四倍體馬鈴薯標記開發困難的問題, 實用性、篩選效率及準確性都極可靠, 對四倍體馬鈴薯遺傳育種研究和品種選育都有著重大意義。
分子標記輔助育種技術能極大地縮短育種年限并且提高育種中目的性狀的準確性[17]。迄今為止, 抗晚疫病、熟性、抗病毒病、抗青枯病、薯型、炸片顏色、薯肉顏色等十幾個馬鈴薯重要性狀相繼被標記[9,18-23]。但關于馬鈴薯蛋白質含量緊密連鎖的標記開發的報道極少, 因此, 四倍體馬鈴薯蛋白含量分子標記的開發對于四倍體馬鈴薯分子輔助育種有著重要的意義。本研究開發的分子標記SCAR8-107在四倍體馬鈴薯品種以及四倍體分離群體 F1代材料中 PCR檢測結果與蛋白含量的表型鑒定結果對應度分別達到了72.97%和90.51%, 統計學上達到了極顯著水平, 分子標記 SCAR8-107與馬鈴薯蛋白含量性狀緊密相關, 可以很好地鑒別四倍體馬鈴薯分離群體子代及馬鈴薯品種的蛋白質含量, 對其進行蛋白含量的篩選與輔助選擇。另外, SCAR標記在實際應用中更簡單、有效、更穩定, 可以快速穩定的鑒定大量的馬鈴薯材料[23]。
分子標記的準確性是分子輔助選擇的關鍵, 直接決定著育種的成敗[24]。本研究中, 分子標記 SCAR8-107在四倍體分離群體與四倍體品種中有著極高的篩選率, 但是仍有小部分的分子標記檢測結果與表型的鑒定結果不一致。存在少量的極端材料與對應的親本擴增不一致, 推測這些極端單株為重組單株, 或因為參照二倍體馬鈴薯DM 序列而設計的引物, 存在檢測不完全準確的問題; 其次由于馬鈴薯蛋白含量為數量性狀, 受許多的微效基因控制, 而這些微效基因也會影響標記的關聯度, 本研究尚處前期階段, 只開發了一個準確的分子標記, 隨著后續研究的開展, 更多相關分子標記的開發, 分子標記的檢測將更為精準可靠。另外馬鈴薯蛋白質的含量受組合×地點互作影響顯著[25], 地點的變化也可能導致馬鈴薯蛋白質含量細微變化, 不同品種不同耕作區域內在蛋白質含量上也會產生一定差異。本研究所開發出的SCAR8-107標記在用于檢測的過程中, 所展現出的微小準確性的差異可能也受到此原因的影響。此外, 群體規模較小、氣候原因及表型測定時蛋白質的損耗也有可能對結果產生影響,因此, 要繼續利用多年多點田間種植馬鈴薯材料進行蛋白含量分子標記的開發, 在運用分子標記進行標記輔助選擇時, 要同時選用多個與蛋白含量緊密連鎖的分子標記同步選擇, 結果將更為精確可靠。
更正:發表于本刊 2018, 44(6): 852–858,艾麗娟等 “大豆籽粒硬實加性和上位性 QTL定位”一文中的圖 3更正為:

圖3 檢測到的加性QTL及上位性互作QTL在連鎖群上的分布Fig. 3 Location of additive QTLs and epistatic effects QTLs on linkage groups
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48
