釀酒酵母高級醇合成路徑及關(guān)鍵基因
2018-09-07 02:58:10劉燦珍秦偉帥孫玉霞趙新節(jié)
中國釀造 2018年8期
關(guān)鍵詞:途徑
劉燦珍,秦偉帥,孫玉霞,趙新節(jié)*
(1.齊魯工業(yè)大學(xué) 山東省微生物工程重點(diǎn)實(shí)驗(yàn)室,山東 濟(jì)南 250353;2.山東省農(nóng)業(yè)科學(xué)院農(nóng)產(chǎn)品研究所,山東 濟(jì)南 250100)
釀酒酵母(Saccharomyces cerevisiae)作為二倍體微生物,在酒類發(fā)酵方面的應(yīng)用較為廣泛,是酒類發(fā)酵的主導(dǎo)菌株。其代謝中產(chǎn)生的揮發(fā)性化合物包括高級醇(杏仁和花的氣味)、乙酸酯、乙基酯(水果和花香味)、中長鏈的揮發(fā)性酸(干酪和汗的氣味)、醛類(奶油、水果和堅(jiān)果氣味)等[1],對酒類風(fēng)味有重要影響。高級醇是指≥3個(gè)碳原子的一元醇類(雜醇油),是酵母在酒精發(fā)酵過程的代謝副產(chǎn)物,其含量對酒類的香氣和品質(zhì)有很大影響,是評價(jià)酒質(zhì)量的重要指標(biāo)之一[3]。適量的高級醇會(huì)增加酒體的香氣復(fù)雜性,是酒體香氣的重要貢獻(xiàn)者,但過量可能會(huì)導(dǎo)致腦性癱瘓,影響人體健康[5],因此在不影響酒風(fēng)味的前提下,降低高級醇的含量具有重要意義。該文綜述了高級醇的主要代謝路徑及部分關(guān)鍵基因,以期為構(gòu)建應(yīng)用于工業(yè)生產(chǎn)的低產(chǎn)高級醇菌株提供理論基礎(chǔ)。
1 高級醇代謝的相關(guān)途徑
高級醇的代謝包括埃里希途徑(Ehrlich)和合成代謝途徑(Harris)。高級醇形成的前體物質(zhì)α-酮酸由氨基酸的轉(zhuǎn)氨作用形成,之后經(jīng)線粒體和胞質(zhì)支鏈氨基酸轉(zhuǎn)氨酶(branched-chain amino acid transaminase,BCAATases)催化[4],由α-酮酸經(jīng)脫羧形成醛,并轉(zhuǎn)化為相應(yīng)的雜醇,稱為埃里希途徑[5]。若α-酮酸來源于碳水化合物代謝,則稱為高級醇的合成代謝途徑[4]。
1.1 埃里希途徑
GIUDICI P等[6]用釀酒酵母研究了氨基酸形成高級醇的形成機(jī)制,指出形成高級醇的埃里希途徑為:氨基酸在轉(zhuǎn)氨、脫羧、還原作用下經(jīng)酮酸、醛最終形成相應(yīng)的高級醇(比與原氨基酸少1個(gè)碳)。之后對該途徑在酶及基因調(diào)控方面進(jìn)行了豐富,有研究發(fā)現(xiàn)[7],支鏈氨基酸通過支鏈氨基酸通透酶(由BAP2基因編碼),轉(zhuǎn)運(yùn)至細(xì)胞;然后由BAT1編碼的線粒體轉(zhuǎn)氨酶作用催化氨基酸為α-酮酸,基因BAT2參與胞質(zhì)支鏈氨基酸轉(zhuǎn)氨酶的作用,催化α-酮酸合成氨基酸的路徑;最后經(jīng)2-酮基酸脫羧酶(2-keto-acid decarboxylase,KDCs)脫羧,脫氫酶(dehydrogenase,ADHs)還原為高級醇。苯丙氨酸、亮氨酸、異亮氨酸、纈氨酸、蛋氨酸也通過該途徑生成相應(yīng)高級醇。
1.2 合成途徑
合成途徑(如圖1所示)是由葡萄糖代謝的中間產(chǎn)物α-酮酸,經(jīng)2-酮基酸脫羧酶(KDCs)脫羧、脫氫酶(ADHs)脫氫還原成高級醇[3]。葡萄糖經(jīng)糖代謝生成丙酮酸,再轉(zhuǎn)化為α-酮丁酸、α-異酮戊酸、α-酮異己酸,最后分別生成活性戊醇、異丁醇、異戊醇。其中活性戊醇途徑為:α-酮丁酸分別在乙酰乳酸合酶(ILV1基因編碼)、還原酶(ILV5基因編碼)、脫水酶(ILV3基因編碼)作用下催化為α-酮-3-甲基戊酸,再經(jīng)脫羧、還原作用形成;異丁醇途徑為:由α-酮異戊酸經(jīng)脫羧作用形成異丁醛,再經(jīng)還原作用形成異丁醇;異戊醇途徑:α-酮基異己酸形成異戊醛,再還原為異戊醇,其中α-酮丁酸在ILVs基因編碼的相關(guān)酶催化下轉(zhuǎn)化為α-酮基異戊酸,在LEU4、LEU1基因編碼的相關(guān)酶的催化下,可轉(zhuǎn)化為α-酮異己酸[7]。而形成α-酮丁酸的途徑有3條,丙酮酸途徑:丙酮酸經(jīng)LEU1基因編碼的異丙基蘋果酸合成酶催化為β-蘋果酸甲酯,β-蘋果酸甲酯再轉(zhuǎn)化為α-酮丁酸,其中α-酮異戊酸到α-酮異己酸(亮氨酸合成途徑的前體物質(zhì))的轉(zhuǎn)化也需要LEU1基因參與,因此,該基因的缺失可能對高級醇及亮氨酸的合成造成一定影響;天冬氨酸途徑:天冬氨酸轉(zhuǎn)化為高絲氨酸,高絲氨酸在THR4基因編碼的蘇氨酸合成酶催化下合成蘇氨酸,蘇氨酸經(jīng)ILV1基因編碼的蘇氨酸脫氫酶作用,轉(zhuǎn)化為α-酮丁酸[8];高絲氨酸合成途徑[7]:可能是高絲氨酸轉(zhuǎn)化為中間物質(zhì)O-乙酰高絲氨酸、胱硫醚,最后形成最終產(chǎn)物α-酮丁酸,該途徑未見詳細(xì)報(bào)道(如圖2所示)。
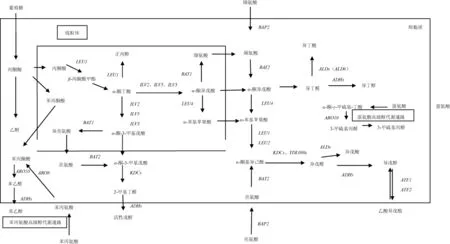
圖1 高級醇代謝通路Fig.1 Metabolic pathways of higher alcohol
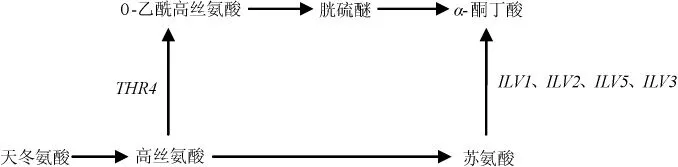
圖2 天冬氨酸合成α-酮丁酸途徑Fig.2 Synthesis pathway of α-keto butyric acid using aspartic acid
2 高級醇調(diào)控關(guān)鍵基因及功能
2.1 埃里希途徑中關(guān)鍵基因及功能
2.1.1 BAP2基因
釀酒酵母BAP2基因編碼支鏈氨基酸通透酶,該酶參與3種支鏈氨基酸包括亮氨酸,異亮氨酸和纈氨酸的轉(zhuǎn)運(yùn),底物特異性較廣泛[9],上述3種氨基酸可通過埃里希途徑形成高級醇[10]。GRAUSLUND M等[9]研究發(fā)現(xiàn),BAP2基因的缺失導(dǎo)致亮氨酸,異亮氨酸和纈氨酸利用量減少20%~50%。也有研究發(fā)現(xiàn)BAP2基因的組成型表達(dá)導(dǎo)致異戊醇生產(chǎn)增加,因此BAP2基因表達(dá)和高級醇的形成之間存在一定的相關(guān)性[11]。
2.1.2 BAT1、BAT2基因
Bat1p和Bat2p酶分別位于線粒體和細(xì)胞溶質(zhì),分別催化支鏈氨基酸和α-酮酸間氨基的轉(zhuǎn)運(yùn),而α-酮酸是合成高級醇的前體,因此會(huì)間接影響酵母發(fā)酵產(chǎn)生的口感和香氣[12]。YOSHIMOTO H 等[13]研究發(fā)現(xiàn),BAT1和BAT2基因分別編碼Bat1p和Bat2p兩個(gè)支鏈氨基酸轉(zhuǎn)氨酶(BCAATases)。BAT2基因參與支鏈醇、乙酸異戊酯的生產(chǎn),BAT2基因缺失突變體KY1058與野生型相比,異戊醇、異丁醇的量分別減少40%、72%[13],另一方面過表達(dá)BAT2基因,異戊醇、異丁醇量分別增加幾倍[14]。此外,雙敲除BAT1和BAT2基因的單倍體釀酒酵母,仍能產(chǎn)生異戊醇[15],表明其他酶能夠補(bǔ)償支鏈氨基酸轉(zhuǎn)氨酶缺失帶來的影響。此外,有研究表明,Bat1p優(yōu)先參與支鏈氨基酸的合成,Bat2p參與纈氨酸的分解代謝,表明了支鏈氨基酸轉(zhuǎn)氨酶的功能多樣化[16]。MA L等[17]敲除CAR1、BAT2并過表達(dá)BAT1基因構(gòu)建了一株低產(chǎn)氨基甲酸乙酯同時(shí)低產(chǎn)高級醇的酵母菌株。
2.1.3 ARO基因
芳香族氨基酸苯丙氨酸經(jīng)苯丙氨酸轉(zhuǎn)氨酶(ARO8基因編碼)、脫羧酶(ARO10基因編碼)催化形成β-苯乙醇,脫羧酶是β-苯乙醇生物合成途徑的關(guān)鍵酶,過表達(dá)ARO10基因β-苯乙醇的產(chǎn)量提高[18]。ARO80、CAT8、MIG1是β-苯乙醇合成重要轉(zhuǎn)錄因子,過表達(dá)CAT8基因或敲除MIG1基因均影響基因ARO9與ARO10的轉(zhuǎn)錄,使苯乙醇的產(chǎn)量增加[19]。而脫羧酶對轉(zhuǎn)氨產(chǎn)物α-氨酮基-γ-甲硫基-丁酸(蛋氨酸轉(zhuǎn)氨物)的脫羧作用尤為突出[20],脫羧酶也是3-甲硫基丙醇生物合成中的關(guān)鍵酶,ARO10基因過表達(dá)使3-甲硫基丙醇的產(chǎn)量提高[21]。
2.1.4 HOM2基因
生物合成蘇氨酸、蛋氨酸的天冬氨酸β-半醛脫氫酶由HOM2基因編碼,因此HOM2基因表達(dá)可能會(huì)影響2種氨基酸的產(chǎn)量,從而導(dǎo)致高級醇含量的變化。ROBICHONSZULMAJSTER H等[22]提出,天冬氨酸β-半醛脫氫酶是埃里希途徑的關(guān)鍵酶。缺失HOM2基因的菌株,異丁醇和異戊醇的生成量顯著降低[23]。齊亞楠[24]研究了HOM2基因?qū)臧l(fā)酵能力與高級醇的影響,發(fā)現(xiàn)敲除HOM2的一個(gè)等位基因,對酵母菌的發(fā)酵性能沒有明顯影響,但異戊醇的生成量降低。敲除HOM2的2個(gè)等位基因則嚴(yán)重影響酵母菌生長,同時(shí)高級醇含量明顯降低。這可能是由于天冬氨酸β-半醛脫氨酶影響了埃里希代謝途徑,也可能是菌體自身生長減弱造成的。
2.2 合成途徑關(guān)鍵基因及功能
2.2.1 LEU基因
異丙基蘋果酸合成酶由LEU1基因編碼,是形成α-酮丁酸和α-酮異戊酸到α-酮異己酸的轉(zhuǎn)化過程的重要編碼基因。因此該基因缺失會(huì)對丙酮酸到α-酮丁酸的形成途徑有阻斷作用,對正丙醇的產(chǎn)量產(chǎn)生影響;同時(shí)該基因缺失影響α-酮酸(α-酮異戊酸-α-酮異己酸)間的轉(zhuǎn)化,導(dǎo)致異戊醇產(chǎn)量降低;而α-酮異戊酸的積累也會(huì)導(dǎo)致異丁醇的生成量增加[25]。有研究通過敲除LEU1基因發(fā)現(xiàn)正丁醇、異戊醇的含量提高,同時(shí)發(fā)現(xiàn)LEU1基因的敲除,使ILV1基因(編碼蘇氨酸脫氫酶)表達(dá)上調(diào)[25]。LEU1是合成亮氨酸的關(guān)鍵基因,通過單敲除及雙敲除LEU1的等位基因發(fā)現(xiàn)異丙基蘋果酸脫氫酶的酶活降低,異戊醇、總高級醇的產(chǎn)量均降低[26]。LEU2基因編碼β-丙基蘋果酸脫氫酶,可催化β-丙基蘋果酸合成α-酮異己酸(異戊醇的前體)[6],因此該基因缺失會(huì)減少α-酮異己酸的量,從而使異戊醇的產(chǎn)量下降,有研究敲除LEU2基因,異戊醇含量較出發(fā)菌株降低了11.82%,高級醇總量降低9.97%[6]。
2.2.2 ILV基因
與支鏈氨基酸合成相關(guān)的酶有4種,由ILV基因編碼。其中異亮氨酸的合成由蘇氨酸脫氨酶(ILV1基因編碼)、乙酰乳酸和酶(ILV2基因編碼)、乙酰羥酸還原酶(ILV5基因編碼)、脫水酶(ILV3基因編碼)、轉(zhuǎn)氨酶(BATs基因編碼)共同催化。同時(shí)在上述后4種酶的作用下丙酮酸催化為纈氨酸,纈氨酸是合成高級醇的前體物,因此ILV基因的表達(dá)會(huì)影響高級醇的合成[27]。在釀酒酵母中分別過量表達(dá)編碼4種酶的基因,異丁醇的產(chǎn)率均升高[28]。BOLLON A P[29]證明,釀酒酵母蘇氨酸脫氨酶參與催化異亮氨酸生物合成的第一步反應(yīng),而且也參與異亮氨酸-纈氨酸的生物合成的調(diào)控途徑。蘇氨酸脫氨酶催化蘇氨酸轉(zhuǎn)化為α-酮丁酸,有研究證明[30]ILV1基因的缺失對乙酰羥酸還原異構(gòu)酶和支鏈氨基酸轉(zhuǎn)氨酶的調(diào)節(jié)沒影響,但導(dǎo)致異丁醇生產(chǎn)率提高到3.5倍。同時(shí)EDEN A等[31]指出,ILV2基因缺失導(dǎo)致中間物α-酮丁酸積累,使正丙醇的生成量增加。
2.2.3 PDC基因
脫羧酶由PDC1、PDC5、PDC6、YDL080c或YDR380w基因編碼,使酵母能利用α-酮-β-甲基戊酸[32]。過表達(dá)釀酒酵母的ILV2、KDC、ADH基因,并且敲除PDC1基因(編碼丙酮酸脫羧酶),發(fā)現(xiàn)異丁醇的產(chǎn)率是出發(fā)菌株的13倍[33],原因可能是丙酮酸合成乙醇的途徑受到抑制,從而導(dǎo)致過多的碳源流向埃里希途徑,進(jìn)而增加了異丁醇產(chǎn)量。也有研究發(fā)現(xiàn)[33],敲除編碼丙酮酸脫氫酶復(fù)合體(負(fù)責(zé)將丙酮酸催化為乙酰輔酶A(coenzyme A,CoA))的基因:PDA1、PDB1、LAT1或LPD1基因,導(dǎo)致異丁醇生產(chǎn)更多。敲除編碼丙酮酸脫羧酶的同工酶基因PDC1基因,導(dǎo)致異丁醇產(chǎn)量增加[34]。類酮酸脫羧酶(YDL080c基因編碼),是α-酮異己酸生成異戊醇的關(guān)鍵酶,研究發(fā)現(xiàn)[34],敲除該基因不能合成類丙酮酸脫羧酶,導(dǎo)致異戊醇的產(chǎn)量顯著減少,可能是由于糖代謝合成高級醇的途徑受阻后,氨基酸代謝為高級醇的分解途徑活躍,而高級醇產(chǎn)量主要來自于糖代謝,因此異戊醇的產(chǎn)量下降。但也有研究發(fā)現(xiàn),YDL080c基因缺失并未造成異戊醇的生成量的降低[35]。
3 展望
高級醇的研究開始于1906年,經(jīng)過110多年的發(fā)展,相關(guān)研究已經(jīng)從生化分析階段進(jìn)入到分子生物學(xué)階段。目前對高級醇的研究已取得很大進(jìn)展,并且不斷豐富高級醇的代謝路徑,找到了高級醇的部分代謝通路及調(diào)控基因。該文綜述了部分高級醇形成路徑埃里希途徑、合成途徑及主要的調(diào)控基因BAP2、BAT1、BAT2、LEU、ILV、THR4、PDC、ARO、HOM2,由于上述基因是埃里希途徑及合成途徑的關(guān)鍵基因,因此綜述上述基因?qū)τ诶斫飧呒壌嫉拇x具有重要意義。目前在高級醇的研究方面由集中于高級醇產(chǎn)量的影響因素和通過何種生產(chǎn)工藝控制高級醇的產(chǎn)量方面,進(jìn)一步深入研究高級醇代謝機(jī)理,以期通過分子生物學(xué)的方式將釀酒酵母進(jìn)行菌種改造,從而達(dá)到工業(yè)生產(chǎn)的要求。然而通過基因工程手段構(gòu)建高產(chǎn)酯,低產(chǎn)高級醇的酵母菌株也存在相應(yīng)問題,如構(gòu)建的酵母菌株中存在抗生素選擇標(biāo)記,因此在應(yīng)用于工業(yè)生產(chǎn)方面還存在安全問題。同時(shí),對于基因調(diào)控、各通路間相關(guān)關(guān)系方面還存在研究空間。因此在研究代謝通路的同時(shí),也應(yīng)在工業(yè)應(yīng)用及發(fā)酵條件對高級醇調(diào)控基因的表達(dá)方面繼續(xù)研究。
猜你喜歡
語數(shù)外學(xué)習(xí)·高中版中旬(2023年2期)2023-05-10 13:26:53
語數(shù)外學(xué)習(xí)·高中版中旬(2022年5期)2022-07-13 20:47:51
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2019年10期)2019-11-25 07:33:58
中學(xué)生數(shù)理化·高一版(2018年9期)2018-10-09 06:46:50
湖南教育·C版(2018年3期)2018-06-05 16:54:36
中學(xué)生百科·大語文(2017年10期)2017-11-04 06:56:38
中國衛(wèi)生(2016年3期)2016-11-12 13:23:26
公民與法治(2016年22期)2016-05-17 04:20:13
中國衛(wèi)生(2014年12期)2014-11-12 13:12:52
癌變·畸變·突變(2014年6期)2014-02-27 06:15:03
