鈣泵的發現歷程
2018-09-09 09:47:30任衍鋼白冠軍宋玉奇路彥文
生物學通報 2018年10期
任衍鋼 白冠軍 宋玉奇 路彥文
(陽泉師范高等專科學校 山西陽泉 045200)
鈣泵的發現具有漫長的歷史背景。早在1848年,德國生理學家雷蒙德(Emil du Bois-Reymond)在研究動物電時就假定細胞膜上存在主動運輸的可能性。1948年,羅森博格(Rosenberg T.)重新闡述主動運輸(Active transport)的概念[1]。1957年,丹麥科學家斯科(Skou J.C.)首先發現了第1 個具有主動運輸意義的離子泵——鈉鉀泵。鈉鉀泵的發現被認為是開啟了人類認識離子泵的先河,斯科也因此分享了1997年的諾貝爾化學獎[3]。鈣泵則是繼鈉鉀泵后被發現的第2 個離子泵。鈣泵的研究主要在肌質網鈣泵(sarco/endoplasmic reticulum Ca2+ATPase,SERCA)、動物細胞質膜鈣泵(plasma membrane Ca2+ATPase,PMCA)和植物細胞鈣泵3 個相對獨立的領域進行,下文按照歷史的進程分別介紹。
1 肌質網鈣泵的發現及揭示
鈣泵的發現源自于鈣離子對動物肌肉收縮的影響研究。早在1885年,英國生理學家榮格(Ringer S.)發現離體青蛙心跳在蒸餾水和自來水中的表現不同(自來水中含有鈣離子),他通過逐漸增加鈣離子濃度的實驗發現,鈣是支持心臟收縮的重要因素[1]。為什么鈣離子能影響肌肉的收縮?1940年,赫爾布輪(Heilbrunn L.V.)通過注射Ca2+到肌纖維中證實,鈣離子是在肌纖維中起作用的[2]。20世紀40年代末,學者希爾(Hill A.V.)等在骨骼肌收縮的研究中也注意到這一點。20世紀50年代安瑪莉(Annemarie W.)通過量化的方法發現在Mg2+和ATP 存在的情況下,肌肉收縮需要低濃度的Ca2+。那么,肌肉收縮時鈣離子來自哪里?人們將肌纖維含有豐富的肌質網聯系起來。雖然早在1902年,意大利生理學家瓦拉蒂(Veratti E.)就用光學顯微鏡對肌質網作了精確的描述,但并不清楚它的作用。直到20世紀50年代后,電子顯微鏡的應用和微量鈣濃度檢測生化技術的改進才為人們深入研究肌質網與鈣離子的活動提供了技術條件。1962年,江橋節郞(Setsuro E.)和李普曼(Lipmann F.,1935年諾貝爾生理學獎獲得者)等發現,ATP 驅動Ca2+脫離肌原纖維進入肌質網后肌肉松弛[3]。此后江橋節郞(1964)進一步發現,Ca2+從原肌球蛋白排出后肌肉才發生松弛[4]。1970—1971年,麥克倫南(Maclennan D.H.)和王(Wong P.T.S.)用脫氫膽酸鹽和鹽沉淀方法首次純化了肌質網鈣泵(SRCa2+Mg2+ATPase)。1985年,他們又用DNA 克隆方法獲得肌質網鈣泵全部氨基酸序列,并根據一級結構和X-光中子衍射圖譜等提出鈣泵的空間結構模型,他們的研究被認為是開辟了分析離子泵轉運機制的新時期。同年,博爾曼(Berman M.C.)等通過外源熒光探針標記實驗發現內質網鈣泵存在2 種蛋白構象,一種為E1,對Ca2+親和力高;另一種為E2,對Ca2+親和力低,2 種構象α-螺旋含量不同,結合位點互相有協同作用[5]。
肌質網鈣泵研究取得突破性進展是在21世紀初。2000年,豐島(Toyoshima)等利用分辨率0.26 nm 的透鏡得到肌質網膜上結合Ca2+的鈣泵晶體結構。2年后他們又得到了結合Ca2+的鈣泵的晶體圖,隨后對鈣泵傳導Ca2+過程中的不同鈣泵晶體結構也被結晶出來。他們發現這與肌質網鈣泵的拓撲性質相同[3]。2004年,丹麥科 學家索倫森(Sorensen T.L.)等也結晶出2 個不同狀態的鈣泵晶體結構。李(Lee A.G.)在此基礎上進一步研究Ca2+的結合位點。這期間,斯托克斯(Stokes D.L.)小組提出了一個描述鈣泵酶中催化循環過程的結構模型(圖1)。此外,人們還發現,即使在Mg2+、Na+、K+的濃度比Ca2+的濃度高出數萬倍的情況下,鈣泵酶仍然會選擇性吸收Ca2+。在理論方面,杉田(Sugita Y.)等運用原子-分子動力學模擬的方法確認Ca2+結合位點處酸性殘基的質子化狀態。卡斯特(Costa V.)等運用分子模擬和靜電計算研究Ca2+結合過程中幾個關鍵中間狀態的結構和動態特征[6]。這一系列研究對進一步了解肌質網鈣泵及研究心肌缺血的機制提供了新的參考依據。

圖1 鈣泵循環和鈣離子通道示意圖
2 動物細胞質膜鈣泵的發現及揭示
在鈉鉀泵被發現之前,人們已經注意到細胞對微小的鈣離子濃度變化具有很強的敏感性。但是這種差異是如何形成的,曾經是個迷,鈉鉀泵的發現對研究這種差異無疑是一個重要的啟示。1966年,齊默曼(Zimmerman A.N.E.)等發現,用無鈣的“生理鹽溶液”灌注大鼠離體心臟,短時間內即發生心肌膜損傷,隨后灌注正常含鈣的生理溶液,心臟發生更為嚴重的結構和功能的改變,這種現象被稱之為鈣反常[7]。這種現象顯然與細胞內鈣離子增多(鈣超載)有關。同年,沙茨曼(Schatzmann H.J.)在研究人類紅血細胞時發現,ATP 酶可從細胞質中轉運出鈣離子,他將這種蛋白質命名為質膜鈣泵(PMCA),這是人們首次將鈣轉運與ATP酶聯系起來[8]。1977年,加里特(Jarrett H W)等首先證實有一種蛋白(鈣調蛋白/鈣調素,簡稱CaM)直接參與質膜轉運鈣的活性調節[9]。1979年,尼哥(Niggle V.)等利用鈣調蛋白親和層析方法將較多量的質膜鈣泵純化出來。通過純化發現,質膜鈣泵由4 種基因編碼,其轉錄經不同類型的選擇性拼接后可形成多種類型和變體。這些亞型和變體具有不同的Ca2+的外排性質,并且在不同組織和細胞中具有各異的表達水平,以滿足不同類型細胞對于Ca2+精確調控的需要[10]。1980年,卡羅尼(Caroni P.)和卡洛福林(Carafolin E.)在研究動物細胞鈣泵的過程中發現鈣泵的主要轉運特點是將Ca2+運出細胞之外[11]。1983年,科學家依據質膜鈣泵大小約為251kDa,其大小接近單體人紅細胞酶的2 倍推測,該離子泵以二聚體形 式存在[12]。1988年,沙爾(Shull G.E.)和格潤(Greeb J.)以大鼠腦細胞為材料,針對ATP 酶結合位點,設計寡核苷酸探針,純化得到cDNA 后推出質膜鈣泵的氨基酸一級序列,發現質膜鈣泵包括2 個亞型,分別含1 176 和1 198 個氨基酸[13],他們還依據一級結構推測出鈣泵的拓撲學結構。同年,維爾馬(Verma A.K.)等則利用紅細胞質膜鈣泵的胰蛋白酶水解片段設計的寡核苷酸探針,從人畸胎瘤cDNA 文庫中推導出的是含1 220 個殘基的完整亞型的氨基酸序列。比較發現,該序列與大鼠中獲得的第1 個亞型的前1 117 個殘基有大約99%的相同[14]。1994年,費根(Fagan M.J.)和賽爾(Saier M.H.)通過用腎皮質Na+/K+-ATP 酶與質膜鈣泵比較表明,這2 種泵具有類似的反應循環和35%相同的氨基酸序列,同屬于P 型ATP 酶系列[15]。但是,需要指出的是,因三維晶體結構其胞質區部分較大,本身難形成構象,而蛋白質的有序排列又是解晶體結構的必要條件,且對蛋白質純化水平也有較高的要求,所以,至今對質膜鈣泵的了解仍然停留在拓撲學結構上,需要進一步攻克。
3 植物細胞鈣泵的發現及揭示
植物中鈣的作用過去被認為是一個難以捉摸的問題。鈣在植物中的總量并不小(用mmol/L 計算),但就需求而言卻屬于微量營養素(micronutrient,用μmol/L 量計算),其原因在于它很大一部分存在于細胞壁處,極少的一部分存在于細胞內。植物細胞鈣泵的發現主要是受動物細胞鈣泵研究的影響。如早在20世紀50—60年代,科學家就在動物肝和腎細胞中發現,線粒體具有吸收和積累貯存Ca2+的能力,并且這種吸收與呼吸和ATP 活動有關,并且受解偶聯劑的抑制。1965年,霍奇斯(Hodges T.K.)和漢森(Hanson J.B.)在玉米細胞中發現同類的現象[1]。但是這些發現(包括一些用放射性元素45Ca2+)雖然涉及到鈣離子從細胞流出及ATP 的水解活動,但還不足以確定鈣泵是來自于質膜還是細胞內的其他膜系統。直到20世紀70年代,由于科學家發現線粒體中的NADH 脫氫酶受Ca2+的調節等,植物線粒體鈣泵的存在才獲得了學術界的公認。故在植物細胞中的鈣泵首先在線粒體上發現。20世紀80年代進一步發現,植物線粒體鈣泵還受光的影響[1],緊接著發現液泡膜上存在鈣泵。1990年,布萊克福德(Blackford S.)等發現,在植物液泡中同樣存在積累Ca2+的現象,1993年,菲佛(Pfeiffer W.)和海格(Hager A.)在玉米中證實液泡膜鈣泵的存在[16]。
植物細胞質膜鈣泵發現于20世紀80年代末至90年代初,由于技術上的改進,科學家才從蘿卜苗、菠菜葉等植物組織中(通過排除其他膜系統上鈣泵)證實植物細胞質膜上同樣存在鈣泵[17]。
植物細胞內質網鈣泵于1992年在高等植物土豆和煙草中被克隆出來(它的同系物于1996 和1997年在擬南芥中發現)。1993—1994年,大衛(David E.E.)和洛林(Lorraine E.W.)發現在植物內質網中同樣存在動物細胞內質網中的鈣結合蛋白的同系物,且發現內質網膜上的鈣泵不同于質膜上的鈣泵而是與動物細胞內質網膜上的鈣泵類似,植物內質網膜上的鈣泵也不存在Ca/H 的逆向轉運[16]。植物內質網膜上的鈣泵主要參與胞漿中Ca2+的調節及在種子發芽、 分泌中起重要的作用。學者還發現,質膜鈣泵與內質網鈣泵的差異主要是底物的差異,大小119 kDa 的鈣泵既存在于質膜也存在于內質網膜上,而大小124 kDa 的鈣泵僅存在于質膜上[16]。
質體內的鈣泵于1994年被黃(Huang L.Q.)等確定,它被證實與哺乳動物P 型Ca2+ATP 酶具有同源性[17]。葉綠體上的鈣泵主要發現存在于內膜上(1993—1994)。葉綠體上的鈣泵被認為可能在葉綠體基質和細胞質之間的信號傳導中起作用,以及與葉綠體內的代謝和蛋白質進入葉綠體基質等有關[18]。
植物中鈣泵的類型在20世紀90年代下半期被鑒定出來,科學家發現,植物質膜鈣泵的單體普遍小于哺乳動物的單體[17]。但他們也屬于P-型ATP 酶家族[18]。植物質膜鈣泵的主要功能被認為就是維持胞漿Ca2+的低濃度水平,此外,還有細胞內信號傳導、分泌、氣孔等調節的功能,也與有絲分裂、向地性、向光性等相關。科學家還發現,盡管不同類型的鈣泵作用不同,但都參與細胞中第2 信使的活動,并與蛋白質的活性穩定和維持、細胞的分泌和生長等密切相關。
總之,從榮格的發現到今天,對鈣在細胞內作用的探討已經走過漫長的道路,20世紀60年代鈣泵的發現揭開了這項研究新的一頁,20世紀90年代至21世紀初又獲得一系列重要的進展。對上述的要點簡要歸納為圖2和圖3。
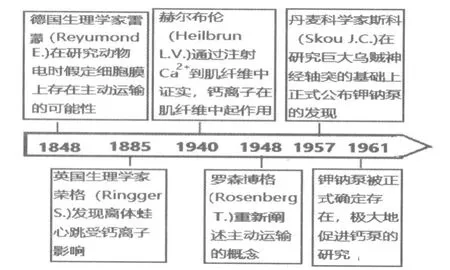
圖2 鈣泵發現的歷史背景
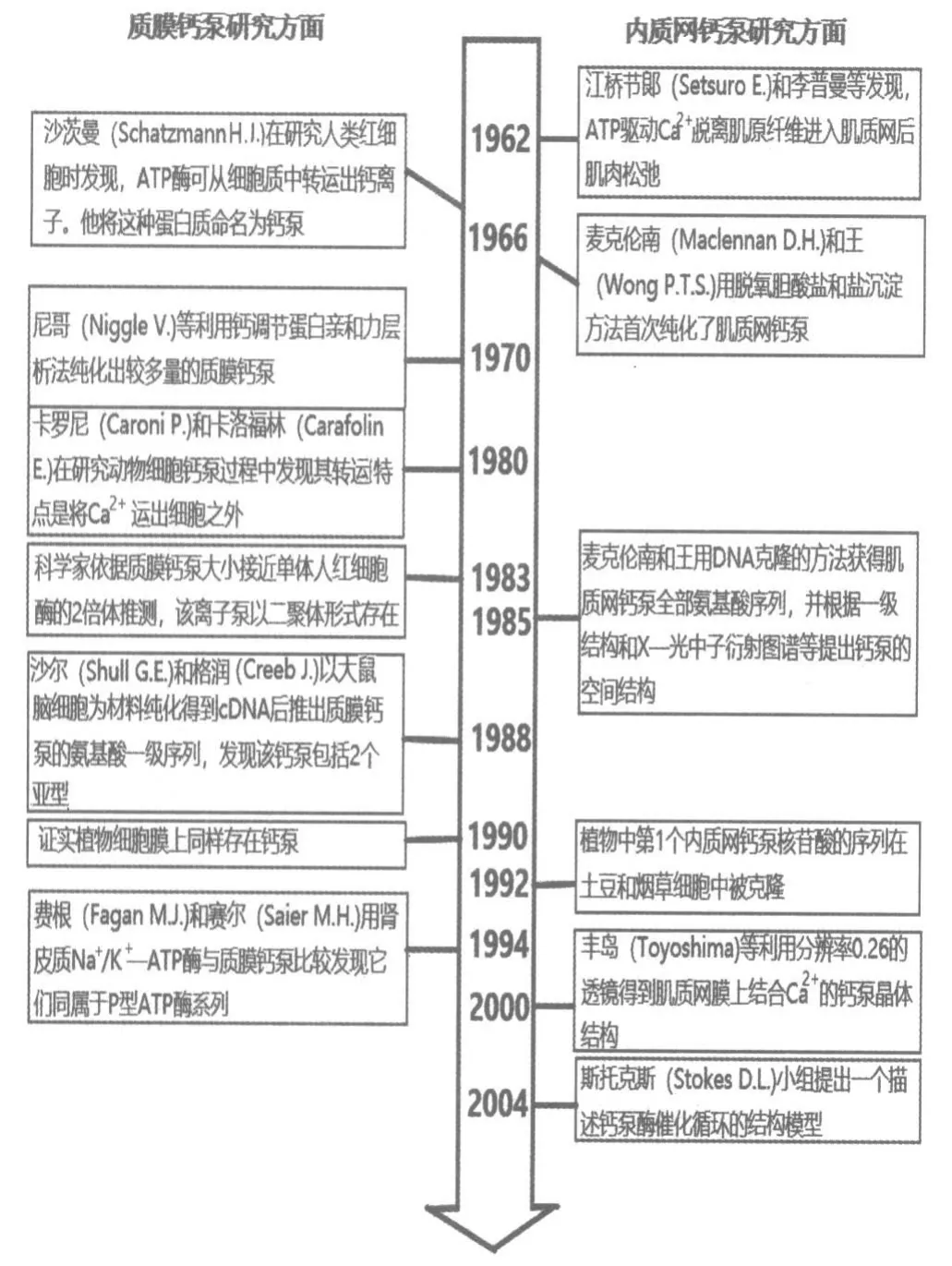
圖3 鈣泵發現中的重要事件
綜上所述,鈣泵發現的歷史順序是沿著肌質網鈣泵—動物細胞質膜鈣泵—植物細胞鈣泵的進程,它們之間既相對獨立又相互影響。與鈉鉀泵研究相比,鈣泵研究相對滯后,例如鈣離子的跨膜傳輸過程及鈣泵在信號傳導和細胞凋亡中的分子機制等現在仍不完全清楚。鈣泵研究的重要性不亞于其他離子泵,它的突破性進展需要引起進一步的關注。鈣泵的作用廣泛而重要,尤其是在鈣行使第2 信使功能時起著非常重要的作用,被認為是生命和死亡的信號。正如著名的生理學家洛維(Otto Loewi 1873—1961,1936年諾貝爾生理學獲獎者)生前預言:是的,鈣是一切!
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
汽車工程學報(2017年2期)2017-07-05 08:13:02
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
