人為干擾對中亞熱帶森林生物量及其空間分布格局的影響
2018-10-18 02:26:48李尚益陳金磊李雷達劉兆丹張仕吉
生態(tài)學報 2018年17期
李尚益,方 晰,2,*,陳金磊,李雷達,辜 翔,劉兆丹,張仕吉
1 中南林業(yè)科技大學生命科學與技術學院,長沙 410004 2 湖南會同杉木林生態(tài)系統(tǒng)國家野外科學觀測研究站,會同 438107
森林生物量是研究和評價森林生態(tài)系統(tǒng)結構與功能過程最基本的參數(shù),不僅能揭示森林生態(tài)系統(tǒng)能量平衡、養(yǎng)分循環(huán)和生產(chǎn)力等功能過程的變化規(guī)律[1- 3],而且能反映生態(tài)系統(tǒng)功能的強弱,對研究森林生態(tài)系統(tǒng)碳循環(huán)過程和全球氣候變化具有重要意義[4- 7]。20世紀70年代初,在國際生物學計劃(International Biological Programme,IBP)和人與生物圈計劃(Man and the Biosphere Programme,MAB)[8]推動下,森林生物量研究迅速發(fā)展,研究方法和技術多樣,生物量估算準確度不斷提高,特別是20世紀90年代國際地圈-生物圈計劃(International-Geosphere-Biosphere Programme,IGBP)的實施以及國際社會對全球氣候變化的關注,森林生物量研究再次成為現(xiàn)代生態(tài)學研究熱點。目前,森林生物量研究已在個體、種群、群落、生態(tài)系統(tǒng)、景觀、區(qū)域、生物圈等多個尺度上進行了研究[9-10],基于樣地調(diào)查[11-12]、遙感技術[6]、激光雷達技術[2]和生態(tài)過程模型[13]對不同地理種源[11-13]、發(fā)育階段[11,14]和自然地帶[15-17]的生態(tài)系統(tǒng)生物量的估算,也有圍繞氣候、環(huán)境、資源等人類社會生產(chǎn)實踐的生態(tài)系統(tǒng)生物量動態(tài)變化研究[18-19]。但基于樣地調(diào)查數(shù)據(jù),對不同程度的人為干擾和保護對森林生物量影響的研究仍少見報道。
中國亞熱帶地區(qū)水熱條件優(yōu)越,森林類型多樣和物種豐富[20],是中國重要的農(nóng)林業(yè)生產(chǎn)基地,也是中國森林受人類干擾最早和最為嚴重的地區(qū)之一。常綠闊葉林是該地區(qū)的地帶性植被類型,是亞熱帶陸地生態(tài)系統(tǒng)的重要組成部分[21-22],但由于人為干擾嚴重,取而代之的是大面積的次生林、次生灌叢和灌草叢等[23],群落結構趨于簡單,抗外界干擾能力下降、涵養(yǎng)水源和土壤肥力質量減弱等特征。研究森林生態(tài)系統(tǒng)結構和功能過程對不同人為干擾程度的響應,無疑對揭示森林生態(tài)系統(tǒng)在人為干擾下的退化機制,探討退化植被恢復和重建具有理論和實踐雙重意義。目前國內(nèi)多數(shù)研究將重點放在不同程度的人為干擾和保護對群落結構和物種多樣性[24-28],土壤肥力質量和蓄水能力[29-31]等方面的研究,在人為干擾對森林生物量影響方面的研究,也主要集中在人為經(jīng)營管理措施(如采伐、撫育間伐、造林、施肥等)對人工林生物量影響的研究[32-35],而對處于不同人為干擾程度的次生林、次生灌叢和灌草叢生物量的比較研究很少,對揭示人為干擾對森林生態(tài)系統(tǒng)結構與功能過程的影響機制仍缺乏相關數(shù)據(jù)。為此,本研究以湘中丘陵區(qū)4種處于不同程度人為干擾的植物群落:檵木(Loropetalumchinense)-南燭(Vacciniumbracteatum)-滿山紅(Rhododendronmariesii)灌草叢、檵木-杉木(Cunninghamialanceolata)-白櫟(Quercusfabri)灌木林、馬尾松(Pinusmassoniana)-石櫟(Lithocarpusglaber)-檵木針闊混交林和石櫟-紅淡比(Cleyerajaponica)-青岡(Cyclobalanopsisglauca)常綠闊葉林為對象,研究不同程度的人為干擾后森林群落生物量的空間分布格局及其影響因素,為探討人為干擾后的森林生態(tài)恢復機制,為準確地反映人類活動與森林生態(tài)功能過程的關系,以及為促進中亞熱帶森林植被恢復與保護、環(huán)境與經(jīng)濟可持續(xù)發(fā)展提供科學依據(jù)。
1 研究地概況
研究地設置在湖南省長沙縣(113°17′—113°27′E,28°23′—113°24′N),兩地空間距離約為28km,地處幕連九山脈中支連云山山脈的余脈,地形起伏較大,海拔高度為100—550 m,坡度多在20°—30°之間,屬于典型的低山丘陵地貌。該區(qū)年平均氣溫17.0℃,7—8月份極端高溫40℃,1月份極端低溫-11℃,雨量充沛,相對濕度較大,年降水量1412—1559 mm,降雨集中在4—7月份,為中亞熱帶東南季風濕潤氣候。土壤以板巖和頁巖發(fā)育而成的紅壤為主,地理位置和自然條件決定了該區(qū)地帶性植被為中亞熱帶常綠闊葉林,屬于湘中湘東山丘盆地栲(Castanopsisfargesii)林、馬尾松林、毛竹(Phyllostachysheterocycla)林、油茶(Camelliaoleifera)林及農(nóng)田植被區(qū)的幕阜、連云山山地丘陵植被小區(qū)。由于該區(qū)人為干擾(采伐、火燒)頻繁,強度各異,原生天然植被常綠闊葉林破壞比較嚴重,形成了多種處于不同程度的人為干擾植物群落(灌草叢、灌木林、馬尾松針闊混交林、常綠闊葉林)以及以杉木、馬尾松、濕地松(Pinuselliottii)為主的人工林群落,為中亞熱帶森林群落恢復(或演替)及其相關研究提供了場所。
2 研究方法
2.1 樣地設置
在地域相鄰,海拔、坡度、坡向以及土壤、氣候條件基本一致,處于不同人為干擾程度的4種植物群落:檵木-南燭-滿山紅灌草叢、檵木-杉木-白櫟灌木林、馬尾松-石櫟-檵木針闊混交林、石櫟-紅淡比-青岡常綠闊葉林(分別簡寫為:LVR、LCQ、PLL和LAG,下同)內(nèi)設置長期定位觀測樣地:LVR群落設置4個20 m×20 m樣地,LCQ群落設置3個20 m×20 m樣地;PLL、LAG群落均設置3個30 m×30 m樣地。4種植物群落的人為干擾的基本概況如下:
(1)LVR群落(嚴重干擾):1965年冬煉山、人工全墾整地,1966年春營造的馬尾松人工純林,經(jīng)營期間無施肥歷史,1990年皆伐后,多年遭到反復砍伐、火燒、放牧活動。2012年后,停止各種人為干擾。
(2)LCQ群落(較重干擾):1965年冬煉山、人工全墾整地,1966年春營造的杉木人工純林,經(jīng)營期間無施肥歷史,1988—1989年皆伐后,每隔3—5年擇伐一次。2004年后,停止擇伐。
(3)PLL群落(中度干擾):20世紀70年代初,天然常綠闊葉林采伐后,自然恢復為39%針葉樹馬尾松,25%常綠闊葉樹石櫟,11%檵木和23%其他樹種組成的針闊混交林。
(4)LAG群落(無明顯人為干擾):20世紀60年代初以來,無明顯人為干擾,保存比較完好,現(xiàn)為為69%常綠闊葉樹(其中38%石櫟,18%紅淡比和5%青岡)和31%其他樹種組成的常綠闊葉林。
2.2 樣地群落調(diào)查
2016年10—11月落葉樹種落葉前完成,采用樣方調(diào)查法進行群落調(diào)查,所有植物均鑒定到種。(1)在LVR每塊樣地,沿對角線均勻設置4個2 m×2 m樣方調(diào)查灌木層、草本層植物,記錄植物名稱、株數(shù)、多度、蓋度、平均高度和生活型等。(2)在LCQ每塊樣地,沿對角線均勻設置4個5 m×5 m樣方調(diào)查灌木層植物,記錄植物名稱、株數(shù)、多度、蓋度、樹高和生活型,對樹高高于1.5 m的灌木,按照一定順序編號掛牌,測定胸徑(DBH)、樹高、冠幅、枝下高、健康狀態(tài);對樹高低于1.5 m的灌木,測定地徑、樹高;草本層植物調(diào)查與LVR草本層植物的調(diào)查相同。(3)在PLL、LAG每塊樣地,對樹高高于1.5 m的木本植物,按照一定順序編號掛牌,記錄植物名稱及其所在層次,測定胸徑、樹高、冠幅、活枝下高、健康狀態(tài);灌木層、草本層植物調(diào)查與LCQ樣地灌木層、草本層植物的調(diào)查相同。
計算4種植物群落木本植物的密度、平均胸徑、平均樹高,Shannon-Wiener多樣性指數(shù)等特征指標。4種植物群落的基本特征及其主要樹種組成如表1所示。
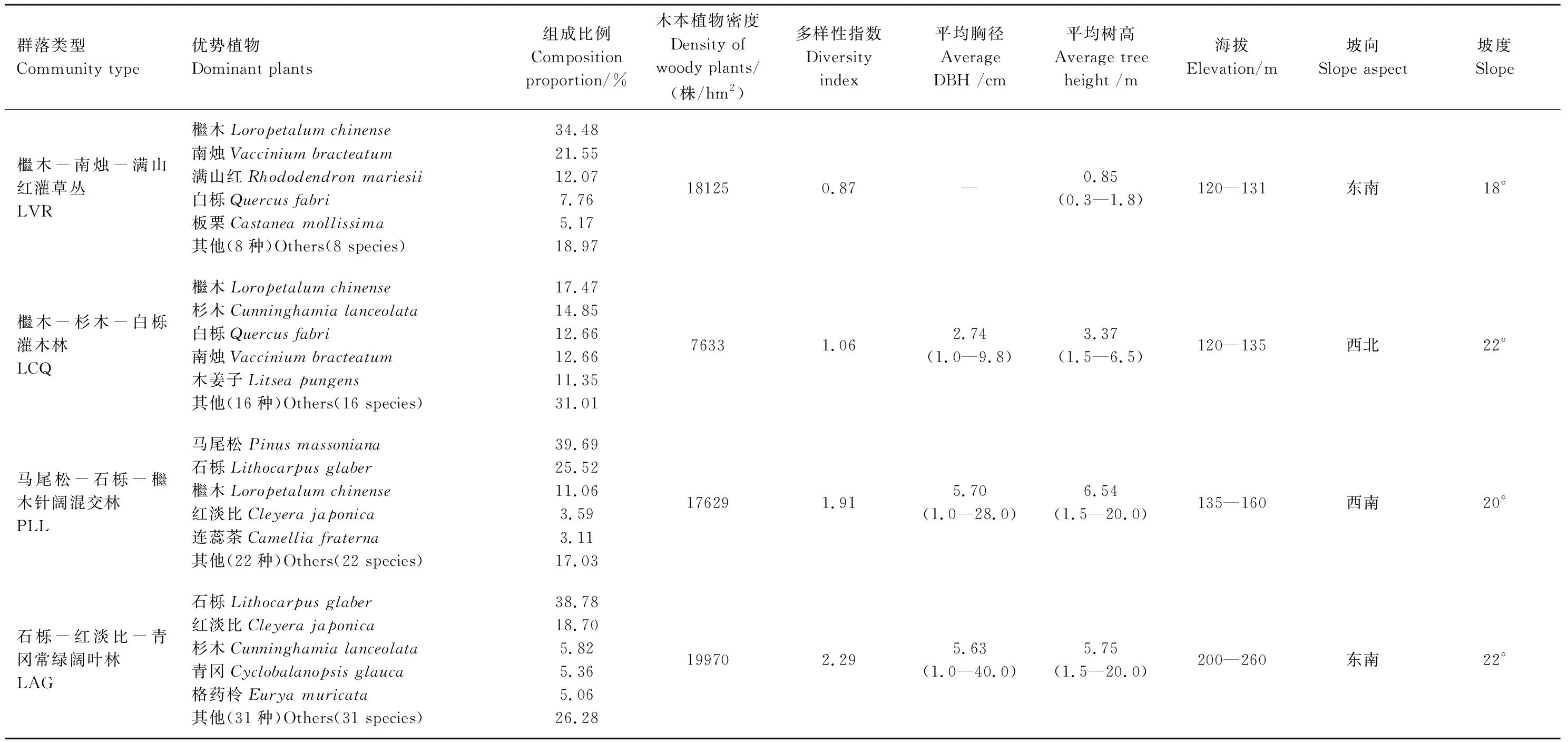
表1 樣地的基本特征
LVR:Loropetalumchinense-Vacciniumbracteatum-Rhododendronmariesiiscrub-grass-land,LCQ:Loropetalumchinense-Cunninghamialanceolata-Quercusfabrishrubbery,PLL:Pinusmassoniana-Lithocarpusglaber-Loropetalumchinenseconiferous-broad leaved mixed forest,LAG:Lithocarpusglaber-Cleyerajaponica-Cyclobalanopsisglaucaevergreen broad-leaved forest
2.3 生物量的測定
在樣地群落調(diào)查基礎上,采用收獲法測定群落生物量,2016年10—11月落葉樹種落葉前完成。4種群落生物量具體的測定方法如下:
2.3.1 檵木-南燭-滿山紅灌草叢
在每塊樣地的4條邊界外圍隨機設置1個2 m×2 m樣方,記錄樣方中灌木、藤本植物名稱、株(叢)數(shù)和草本植物名稱、叢數(shù)。收割樣方內(nèi)的全部植物,同種灌木分為葉、枝、干、根,同種藤本植物分為葉、干和根,同種草本植物分為地上部分和地下部分,測定鮮重。將同樣地的4個樣方同種植物相同器官混合均勻,采集分析樣品(約取0.5 kg)。
同時,設置1個1 m×1 m樣方,根據(jù)地表凋落物層的分層標準[36],按未分解層、半分解層、已分解層收集樣方內(nèi)全部的凋落物,測定鮮重。將同樣地的4個樣方同一分解層的凋落物混合均勻,采集分析樣品(約取1.0 kg)。
將分析樣品置于80℃下烘至恒重,測定含水率。根據(jù)分析樣品的含水率計算各樣方干物質重量,估算樣地單位面積群落的生物量。
2.3.2 檵木-杉木-白櫟灌木林
根據(jù)LCQ樣地群落調(diào)查數(shù)據(jù),計算每塊樣地樹高高于1.5 m的每種灌木平均胸徑、平均樹高,確定為平均標準木。在每塊樣地的外圍,每種灌木分別選取3株平均標準木,按葉、枝、干、根測定平均標準木的鮮重,采集同一樣地3株平均標準木葉、枝、干、根的分析樣品(約取0.5 kg,每樹種每一器官3個重復),用于含水率測定和各組分的干物質重量的換算。根據(jù)各樹種9株平均標準木各組分生物量與胸徑、樹高的關系,構建各種灌木各組分生物量的相對生長方程(表2),計算各種灌木單株生物量。
對樹高低于1.5 m灌木層、藤本層、草本層、地表凋落物層生物量的測定,與LVR群落相同。結合樣地群落調(diào)查數(shù)據(jù),估算樣地單位面積群落各組分的生物量。
2.3.3 馬尾松-石櫟-檵木針闊混交林
根據(jù)PLL樣地群落調(diào)查數(shù)據(jù),計算每塊樣地每一樹種的平均胸徑、平均樹高,在每塊樣地外圍,每一樹種分別選取3株平均標準木(即每一樹種9個重復),按“分層切割法”在1.3、3.6 m處和以后2 m為1個區(qū)分斷開,樹梢部分不足1 m的作梢頭處理,分別測定每株平均標準木葉、枝、干鮮重;地下樹根采用挖掘法測定生物量,以樹樁為中心在1.5 m為半徑范圍進行圓柱形挖掘,深度直至挖出完整的主根,按細根(<0.2 cm)、小根(0.2—0.5 cm)、粗根(0.5—2.0 cm)、大根(>2.0 cm)和根頭測定樹根鮮重,采集各級根系分析樣品。同一樣地同樹種3株平均標準木各組分混合均勻后采集分析樣品(約取0.5 kg,各組分3個重復)。同樣,由各樹種各組分含水率換算成干重,每一樹種根據(jù)9株平均標準木各組分生物量與胸徑、樹高的關系,建立各樹種各組分生物量的相對生長方程(表2),計算各喬木樹種單株生物量;結合樣地群落調(diào)查數(shù)據(jù),估算單位面積每種樹種各組分生物量。
2.3.4 石櫟-紅淡比-青岡常綠闊葉林
喬木層生物量估算:根據(jù)LAG群落調(diào)查數(shù)據(jù),采用PLL建立的馬尾松、石櫟、紅淡比各器官生物量相對生長方程分別估算馬尾松、石櫟、紅淡比單株生物量,采用Ouyang等[37]建立的青岡各組分生物量的通用生長方程(表2)估算青岡單株生物量,采用劉雯雯等[38]建立的區(qū)域尺度杉木各器官生物量通用相對生長方程(表2)估算杉木單株各器官生物量,其他落葉闊葉樹種、常綠闊葉樹種單株各器官生物量分別采用Ouyang等[37]建立的落葉闊葉林、常綠闊葉林各組分的通用生長方程(表2)進行估算;結合群落調(diào)查數(shù)據(jù)估算樣地單位面積各組分生物量。
PLL、LAG群落灌木層、藤本層、草本層、地表凋落物層生物量的測定和估算,與LVR群落的測定方法相同。
2.3.5 相對生長方程的建立
研究表明,主干比較明顯的樹種,以樹高H(m)、胸徑D(cm)或D2H作為自變量可以很好地預測其生物量。本研究樣地的喬木和灌木多為主干較為明顯的樹種,因此以各樹種各組分生物量W(g)為因變量,以樹高H(m)、胸徑D(cm)為自變量[39-40],選用線性函數(shù)(1)、指數(shù)函數(shù)(2)、對數(shù)函數(shù)(3)和冪函數(shù)(4)進行擬合[41],根據(jù)判定系數(shù)R2的大小選取最優(yōu)相對生長方程。
y=ax+b
(1)
y=abx
(2)
y=a+blnx
(3)
y= axb
(4)
群落各層次地上部分生物量是由葉、枝、干(或莖)生物量之和構成,地下部分生物量是由根系生物量構成;整個群落地上部分生物量是由各層次地上部分生物量之和構成,地下部分生物量是由各層次根系生物量與地表凋落物層現(xiàn)存量之和構成。
2.4 土壤樣品的采集、處理及分析方法
在2016年4月、6月、10月采集土壤樣品。采集土壤樣品時,將每個固定樣地對角線分成3等份,在每一等份中間設置1個采樣點(即每一固定樣地布置3個采樣點),清除采樣點地上植物和死地被物,挖掘土壤剖面,按0—10、10—20、20—30 cm分層,從下至上采集土壤樣品,并觀察和記錄土壤的顏色,同時用環(huán)刀法測定土壤容重。在室內(nèi)清除土壤樣品中動植物殘體、石礫后,將同一固定樣地的3個采樣點同一土層樣品等量混合均勻,約取2 kg(每種植物群落每一土層構成3—4個重復),自然風干后,分別過2、1、0.25 mm土壤篩保存于樣品瓶中備用。過2 mm土壤篩的土壤樣品用于測定土壤顆粒組成,過1 mm土壤篩的用于測定pH、水解氮、速效磷和速效鉀含量,過0.25 mm土壤篩的用于測定有機碳、全氮、全磷、全鉀含量。
(三)豬場常見寄生蟲的成因調(diào)查豬寄生蟲的成功控制與良好的管理有關,但事實證明,如果不結合驅蟲藥的正確使用將不足以控制的寄生蟲感染。現(xiàn)已發(fā)現(xiàn)很多管理良好的豬場也存在著嚴重的蠕蟲感染,這表明寄生蟲有令人難以置信的生存、感染和繁殖的能力,它們幾乎能在所有的豬場中生存。豬只需接觸少量感染性蟲卵或幼蟲就可以保持感染狀態(tài),隨后便感染整個豬場。
土壤顆粒組成用吸管法測定,根據(jù)國際土壤質地分類標準劃分土壤質地[42],pH用土水比1∶2.5 pH計法測定,有機碳用重鉻酸鉀-濃硫酸水合加熱法測定,全氮用KN580全自動凱氏定氮儀測定,全磷用堿熔-鉬銻抗比色法測定,全鉀用火焰分光光度計法測定,水解氮用堿解擴散法測定,速效磷用雙酸浸提UV- 5100B紫外可見分光光度比色法測定,速效鉀用醋酸銨浸提-—火焰光度法測定[42]。不同植物群落土壤理化性質如表3所示。
2.5 數(shù)據(jù)統(tǒng)計分析
采用Excel 2010和SPSS 21.0進行數(shù)據(jù)統(tǒng)計處理,用ANOVA單因素方差分析方法分析同一組分(或同一器官)生物量不同群落之間的差異顯著性(P<0.05),用Bivariate相關分析方法分析樣地生物量與樣地樹種多樣性指數(shù)、土壤養(yǎng)分含量的相關性。
3 結果與分析
3.1 不同人為干擾程度森林群落生物量的空間分配格局
3.1.1 喬木層生物量的分配
從表4可以看出,隨著人為干擾程度減弱,喬木層生物量增加,LAG比PLL提高了17.86%,其中LAG枝、干、根生物量比PLL分別提高了3.53%、20.57%和55.52%,而葉生物量下降了30.18%,地上部分生物量增加了12.54%,但無論是喬木層生物量還是同一組分生物量,兩個群落之間的差異均不顯著(P>0.05)。PLL、LAG喬木層各組分生物量的分配格局基本一致,均表現(xiàn)為:干最高,占喬木層生物量的59.13%—60.48%,其次是枝,占17.87%—20.34%,再次是根,占12.38%—16.34%,葉最小,僅占5.31%—8.15%;地上部分生物量占喬木層生物量的83.66%—87.62%,明顯高于地下部分(根)。表明人為干擾程度減弱,有利于喬木層生長,特別有利于地下根系的生長,但對各組分生物量的空間分配格局影響不大。
3.1.2 灌木層生物量的分配
如表5所示,4個群落灌木層生物量、各組分生物量以及地上部分、地下部分生物量均呈現(xiàn)出隨著人為干擾程度減弱,先增加后下降的變化趨勢,LCQ均為最高,且與LVR、PLL、LAG差異顯著(P<0.05),但LVR、PLL、LAG之間差異均不顯著(P>0.05)。

表2 主要樹種各組分生物量的相對生長方程
WL、WB、WS、WR分別表示樹葉、樹枝、樹干、樹根的生物量(kg/hm2),H表示樹高(m),D表示胸徑(cm)
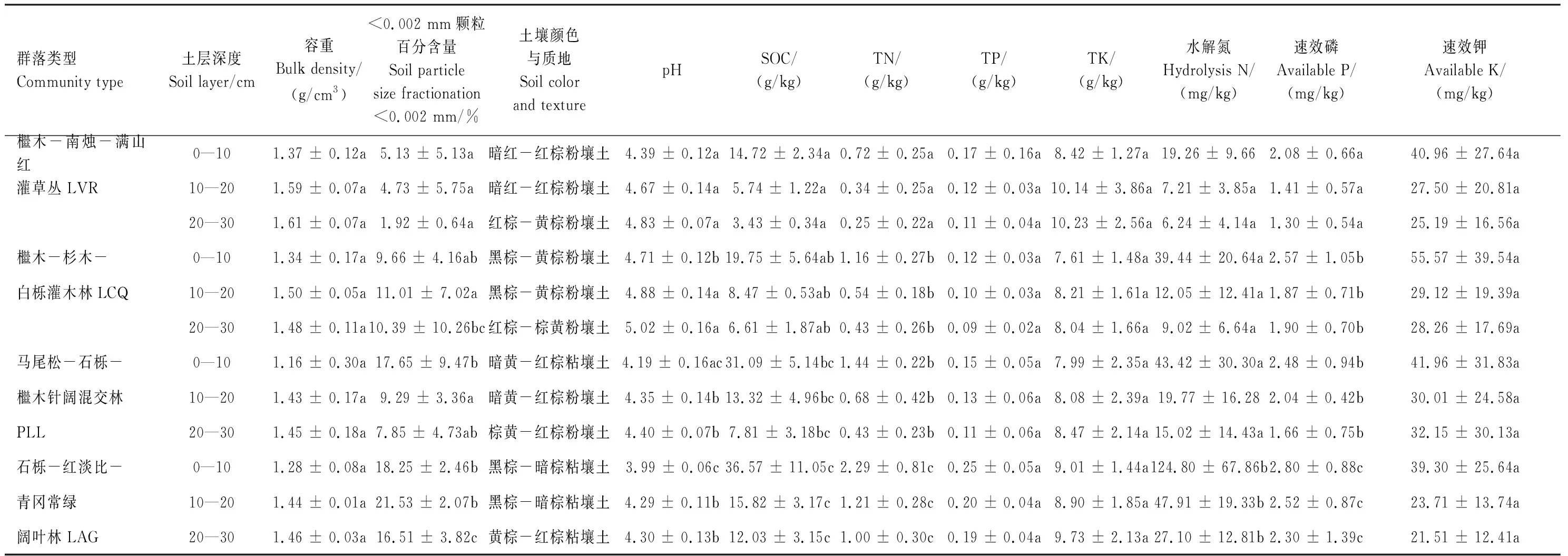
表3 不同森林群落土壤的理化性質
不同字母表示同一土層不同森林群落類型之間差異顯著(P<0.05)

表4 喬木層生物量及其組分分配/(kg/hm2)
括號內(nèi)的數(shù)據(jù)為百分率(%),不同字母表示同一組分不同森林群落之間差異顯著(P<0.05)

表5 灌木層生物量及其組分分配/(kg/hm2)
不同干擾程度群落的灌木層各組分生物量分配格局不同。LVR根生物量最大,占其灌木層生物量的38.93%,其次是枝和干,分別占21.53%和20.43%,葉最小,僅占19.12%;LCQ各組分生物量依次為:干>根>枝>葉,分別占其灌木層生物量的38.63%、33.34%、16.74%、11.28%;PLL依次為:根>干>枝>葉,分別占其灌木層生物量的43.70%、27.47%、15.83%、13.00%;LAG枝(包括干)生物量最高,占其灌木層生物量的42.31%,其次是根,占36.75%,而葉最低。4個群落地上部分生物量顯著高于地下部分(P<0.05),占其灌木層生物量56.25%—66.66%,LCQ群落地下部分生物量占其灌木層生物量的百分比最低,僅為33.34%(表5)。
從表6可以看出,LVR草本層生物量及其地上部分、地下部分生物量均為最高,其次是LCQ和LAG,而PLL最低,呈現(xiàn)出隨著干擾程度減弱先下降再升高的趨勢,但4個群落之間的差異均不顯著(P>0.05)。LVR、PLL和LAG地下部分生物量高于地上部分,其中LVR地上部分與地下部分差異顯著(P<0.05),而PLL、LAG差異不顯著(P>0.05);LCQ地下部分低于地上部分,但差異也不顯著(P>0.05)。

表6 草本層生物量及其組分分配/(kg/hm2)
3.1.4 地表凋落物層現(xiàn)存量
如表7所示,凋落物層現(xiàn)存量及各分解層凋落物現(xiàn)存量總體上隨著干擾程度減弱而逐漸增加,其中LVR與LCQ、PLL、LAG差異顯著(P<0.05),但LCQ、PLL、LAG之間差異不顯著(P>0.05)。LCQ、PLL和LAG凋落物層現(xiàn)存量表現(xiàn)為:已分解層>半分解層>未分解層,而LVR則表現(xiàn)為:半分解層>已分解層>未分解層。未分解層凋落物現(xiàn)存量占其凋落物層現(xiàn)存量的百分比隨著人為干擾程度減弱沒有明顯的變化,半分解層呈下降趨勢,已分解層呈增大趨勢。表明隨著人為干擾程度減弱,未分解層凋落物向半分解層和已分解層遷移速率增大,有利于土壤養(yǎng)分的歸還。
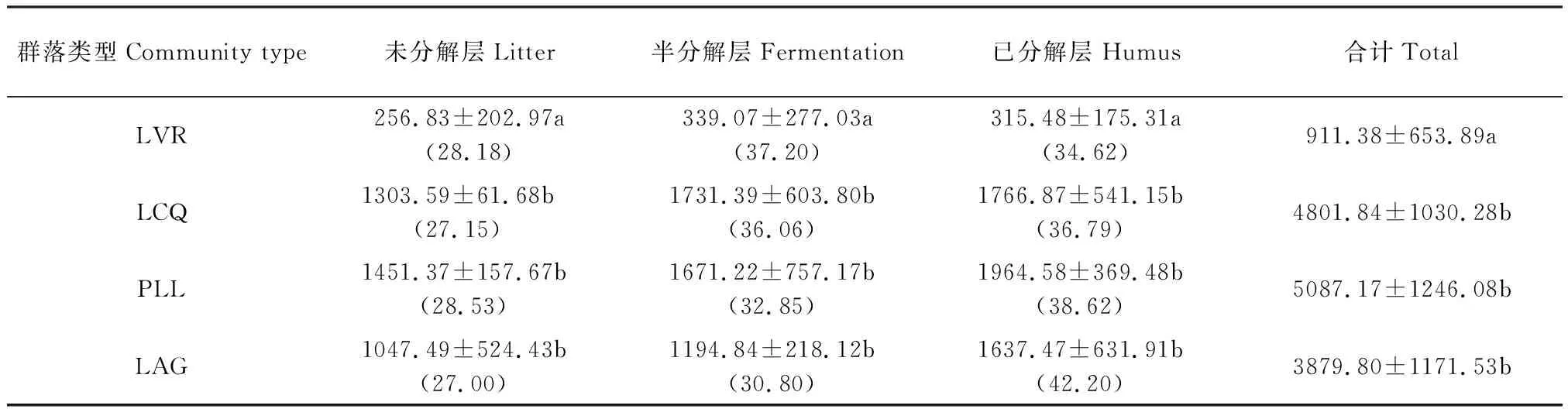
表7 地表凋落物層現(xiàn)存量/(kg/hm2)
3.1.5 群落生物量的空間分配
從表8可以看出,LAG群落生物量最高,分別是PLL、LCQ、LVR的1.15、7.23和28.73倍,且LAG、PLL與LCQ、LVR差異顯著(P<0.05),LCQ與LVR差異顯著(P<0.05),但LAG與PLL差異不顯著(P>0.05),隨著干擾程度減弱,群落生物量呈顯著的指數(shù)函數(shù)增長(y=1927e1.191x,r=0.9594,P<0.05)。不同干擾程度群落生物量的空間分布格局不同。LVR灌木層生物量最高,占群落生物量的42.35%,草本層次之,占40.07%,凋落物層現(xiàn)存量最低。LCQ灌木層生物量最高,占群落生物量的69.98%,其次是地表凋落物層,占23.32%,草本層最低。PLL、LAG喬木層生物量最高,占群落生物量的93.74%—95.76%,占有絕對優(yōu)勢,其次是凋落物層,占2.60%—3.94%,再次是灌木層,占0.78%—1.89%,草本層最低,僅占0.43%—0.86%。與LVR相比,LCQ灌木層、凋落物層生物量顯著增加(P<0.05),但草本層生物量明顯下降;與LCQ相比,PLL、LAG喬木層生物量取代了灌木層的優(yōu)勢地位,而PLL、LAG灌木層生物量顯著下降(P<0.05),草本層、凋落物層生物量變化不大(P>0.05);LAG喬木層生物量的優(yōu)勢比PLL更為明顯。表明群落上層生物量是各群落總生物量的主要來源,且隨著人為干擾程度減弱,群落上層生物量變化顯著,優(yōu)勢逐漸增強,群落下層生物量變化較小,優(yōu)勢逐漸減弱。
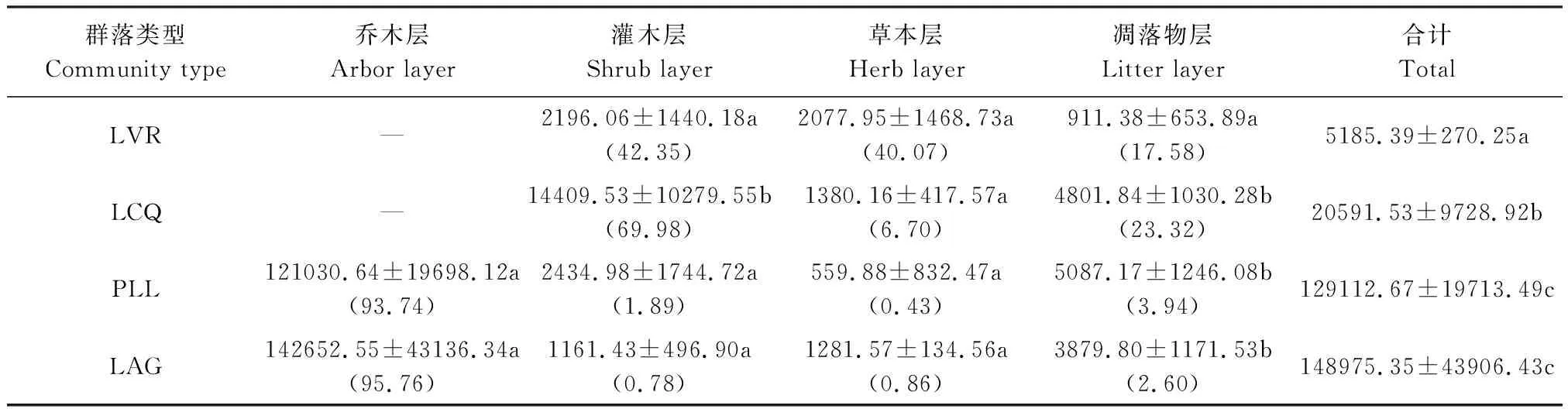
表8 不同干擾程度群落生物量的空間分配/(kg/hm2)
從圖1可以看出,4個群落地上部分、地下部分生物量均隨著人為干擾程度減弱而增加,從LVR到LAG,地上部分生物量增加了118450.7 kg/hm2,提高了5277.47%,地下部分增加了25339.26 kg/hm2,提高了861.61%。表明隨著人為干擾程度減弱,群落地上部分、地下部分生物量表現(xiàn)為異速生長。LVR地下部分生物量比地上部分高31.03%,LCQ地上部分與地下部分相差不大,PLL和LAG地上部分分別比其地下部分高402.76%和326.78%。從LVR、LCQ到PLL、LAG,地上部分生物量占其群落生物量的百分比也呈現(xiàn)增加的趨勢,而地下部分呈下降趨勢。
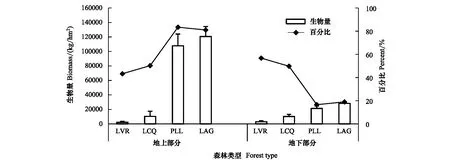
圖1 不同干擾程度群落地上部分生物量和地下部分生物量的分配Fig.1 The distribution of aboveground biomass and underground biomass in different degrees of human disturbance communitiesLVR:Loropetalum chinense-Vaccinium bracteatum-Rhododendron mariesii scrub-grass-land,LCQ:Loropetalum chinense-Cunninghamia lanceolata-Quercus fabri shrubbery,PLL:Pinus massoniana-Lithocarpus glaber-Loropetalum chinense coniferous-broad leaved mixed forest,LAG:Lithocarpus glaber-Cleyera japonica-Cyclobalanopsis glauca evergreen broad-leaved forest
3.2 不同程度的人為干擾群落生物量與物種多樣性的關系
從表9可以看出,群落總生物量與物種多樣性指數(shù)呈顯著正相關(相關系數(shù)為0.554,P<0.05)。地上部分、地下部分生物量分別與物種多樣性指數(shù)呈弱顯著的正相關(相關系數(shù)分別為0.551和0.550,P分別為0.051和0.052)。
3.3 不同程度的人為干擾群落生物量與土壤養(yǎng)分含量的相關性
如表9所示,群落總生物量、地上部分、地下部分生物量分別與0—30 cm土層有機碳、全氮、水解氮含量呈極顯著正相關(P<0.01),其中,與土壤有機碳含量呈現(xiàn)出最強的正相關性(相關系數(shù)為0.875—0.912),與速效磷含量呈顯著正相關(P<0.05),與全磷含量呈正相關,但不顯著(P>0.05),與全鉀、速效鉀含量不存在顯著相關性(P>0.05)。
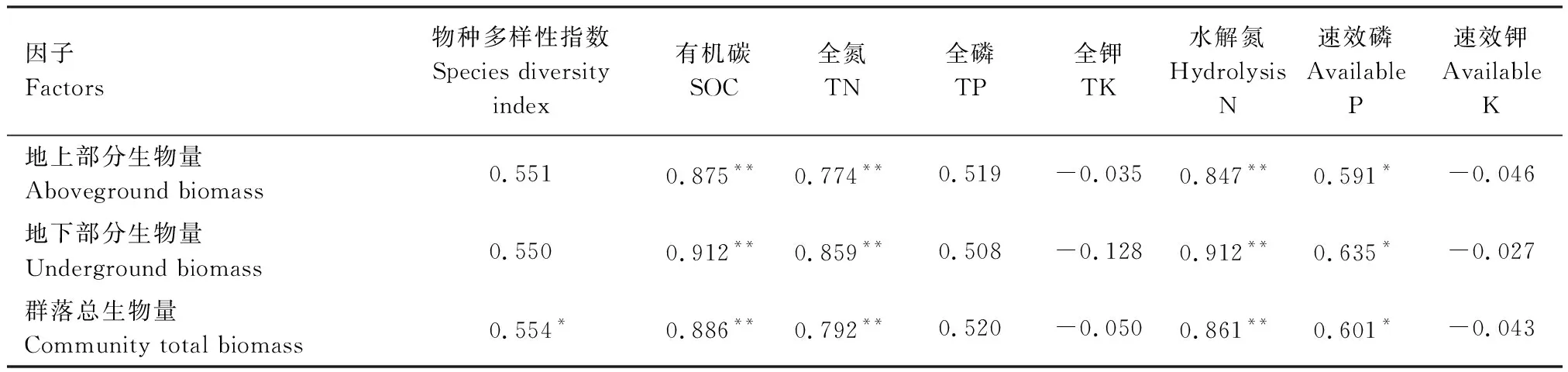
表9 群落生物量與物種多樣性指數(shù)、土壤養(yǎng)分含量的相關系數(shù)
*P<0.05,**P<0.01,n=13
4 結論與討論
4.1 不同程度的人為干擾對群落生物量的影響
群落生物量與植物多樣性的關系一直是陸地生態(tài)系統(tǒng)的研究熱點之一[43],但至今還沒有一致的結論,兩者之間的相互作用機制還不完全清楚[44]。研究表明,在人天混種群中,喬木樹種混交比例不同,在忽略人為干擾歷史(如撫育間伐)的情況下,隨著林齡增加,喬木層樹種增多,物種多樣性指數(shù)增大,生物量顯著增加,多樣性與生物量基本呈單調(diào)的線性增長關系[44]。土壤養(yǎng)分與植物生長密切相關,土壤養(yǎng)分含量直接影響群落的生產(chǎn)力,土壤養(yǎng)分越充足,群落生產(chǎn)力越高[45-47]。本研究中,隨著人為干擾程度減弱,群落生物量呈顯著的指數(shù)函數(shù)增長(P<0.05)。群落生物量與群落樣地樹種多樣性指數(shù)呈顯著的正相關(P<0.05),與0—30 cm土壤層有機碳、全氮、水解氮、有效磷含量呈顯著的正相關(P<0.05)。究其原因是由于,在LVR群落中采樵、火燒、放牧等人為干擾時有發(fā)生,不僅將林木帶出林外,也使得灌木平均樹高增加緩慢甚至下降。另一方面,從林地帶走了大量的養(yǎng)分,不僅顯著減少土壤輕腐殖質輸入量,而且土壤表層大面積裸露,土壤養(yǎng)分流失嚴重,土壤肥力明顯衰退,因而LVR群落以矮小的灌木、草本植物為主,個體數(shù)量雖多但生勢差,群落生物量主要來源于灌木、草本植物,因此LVR群落生物量低。人為干擾程度相對減弱后,LCQ群落灌木層植物平均高明顯增加,而且物種數(shù)明顯增多,地表凋落物層現(xiàn)存量也明顯增加,土壤碳、氮、磷含量明顯提高,灌木層植物得到較好的發(fā)育和生長,因此LCQ群落生物量較顯著增加。停止了采樵等人為干擾活動后,PLL、LAG群落優(yōu)勢樹種由低矮的灌叢或灌木植物恢復為高大的喬木樹種,更具有生長優(yōu)勢,生長速度加快,平均胸徑和平均樹高顯著增加,林分樹種多樣性指數(shù)明顯提高(表1),地表凋落物層現(xiàn)存量明顯積累(表7),土壤養(yǎng)分(C、N、P)含量及其有效性明顯提高(表3),更有利于林木的生長,因此PLL、LAG群落生物量顯著增加。表明不同程度的人為干擾造成群落物種多樣性、土壤養(yǎng)分(C、N、P)含量變化,是導致群落生物量變化的主要因素。群落生物量與0—30 cm土壤層有機碳含量之間存在著最高的相關性,是由于土壤有機碳是生態(tài)系統(tǒng)生產(chǎn)力和更新中最重要的一部分,是植被與土壤恢復的關鍵因素[48]。因此減弱或停止人為干擾,促進灌草叢群落變?yōu)檎蜓萏?提高群落物種多樣性,提高土壤養(yǎng)分含量,改善土壤養(yǎng)分循環(huán),以形成林木生長與土壤改善相互促進的狀態(tài)。
研究表明,群落凋落物層現(xiàn)存量主要取決于群落凋落物量及其分解速率兩個因素[49]。本研究中,凋落物層現(xiàn)存量總體上隨著人為干擾程度減弱而逐漸增加,是由于隨著人為干擾程度減弱,物種種類增加,大型樹木比例增加(表1),群落中物種的新陳代謝加快,凋落物量增加[50];也是由于PLL林下凋落物以針葉為主,較難腐爛分解,而LAG林下凋落物以闊葉樹葉為主,較易于分解,因此PLL凋落物層現(xiàn)存量高于LAG。
4.2 不同程度人為干擾對群落生物量空間分布格局的影響
群落生物量在不同層次上的分配受到群落特性和物種組成等因素的綜合影響[43]。影響群落生物量的因素較多,包括光合、呼吸、脫落、病蟲害消耗等,其中光合作用是一個重要因素,在不同群落中,相同層次的生物量與光照強度密切相關,人為干擾會造成群落光照強度的差異[51]。本研究中,不同干擾程度群落生物量的空間分布格局不同。可能是由于LVR群落遭受采樵、火燒、放牧等嚴重人為干擾后,以矮小的灌木、草本植物為主,沒有形成明顯的灌木層,凋落物不能有效積累,因而灌木層、草本層生物量相當,凋落物層現(xiàn)存量最低;隨著人為干擾程度減弱,沒有形成喬木層的LCQ群落,灌木層植物在全光照條件下能充分發(fā)育和生長,灌木平均高明顯增加,形成明顯的灌木層,生物量顯著增加,林下光照強度減弱,不利于草本層植物生長,生物量明顯下降,另一方面,在調(diào)查過程中發(fā)現(xiàn),LCQ群落落葉灌木樹種較多,大量枯枝落葉的產(chǎn)生使得林下地表凋落物層現(xiàn)存量較高,因而群落生物量呈現(xiàn)出灌木層>凋落物層>草本層的分布格局,與湖南瀏陽大圍山自然保護區(qū)杜鵑灌叢生物量不同層次間的分布格局一致[1];停止了采樵等人為干擾活動后,PLL、LAG形成喬木層,更具有生長優(yōu)勢,代謝更為強烈,凋落物量明顯增加,上層郁閉度增加,林下光照強度減弱,不利于灌木層、草本層植物生長,生物量下降,從而呈現(xiàn)出喬木層生物量占有絕對的優(yōu)勢,其次是凋落物層,灌木層、草本層最低。表明不同程度的人為干擾下,群落植物組成和群落結構不同,群落生物量的空間分配格局不同。
在不同環(huán)境條件下,群落生物量的分配格局反映了植物對環(huán)境的響應規(guī)律和資源分配策略[3]。最優(yōu)分配理論認為,植物通過調(diào)節(jié)各器官生物量的分配來適應外界環(huán)境,從而最大限度地獲取有限資源,以維持其最大生長速率[52]。當光照受到限制時,植物會將資源更多地分配到莖和葉等地上部分,而當水分和養(yǎng)分受到限制時,植物會將更多的資源分配到根系中[53]。本研究中,LVR群落由于人為干擾嚴重,造成地上部分生物量不能有效積累,而樹蔸及根部殘留在林地里,生物量沒有明顯變化,因此無論群落生物量還是灌木層、草本層地下部分生物量高于地上部分。在LCQ群落,沒有形成喬木層,灌木層植物在全光照條件下,有利于地上部分的生長,生物量明顯提高,占其灌木層生物量的比例增高,地上部分生物量與地下部分相差不大。人為干擾減弱或停止后,PLL、LAG群落形成喬木層,物種組成和群落結構逐漸趨于穩(wěn)定,凋落物量顯著增加,土壤水分和養(yǎng)分得到明顯改善,有利于植物的生長和發(fā)育,地上部分、地下部分均有明顯生長,生物量明顯增加,但由于大型喬木樹種增多(表1),對光照的競爭更為激烈,喬木層植物將更多的生物量分配到地上部分,增強對光照的競爭能力,以獲得更大的生長空間和更多的資源,因而地上部分生物量增長速度明顯高于地下部分,從而表現(xiàn)為異速生長,也體現(xiàn)了PLL、LAG群落對亞熱帶地區(qū)溫暖濕潤環(huán)境條件的適應[1,54-55]。
