流速對不同浮游藻類的生長影響研究
2019-01-09 10:32:34周靜茍婷張洛紅藍郁馬千里梁榮昌趙學敏
生態科學 2018年6期
關鍵詞:生長
周靜, 茍婷, 張洛紅, 藍郁, 馬千里, 梁榮昌, 趙學敏,*
?
流速對不同浮游藻類的生長影響研究
周靜1,2, 茍婷2, 張洛紅1, 藍郁3, 馬千里2, 梁榮昌2, 趙學敏2,*
1. 西安工程大學環境與化學工程學院, 西安 710048 2. 環境保護部華南環境科學研究所, 廣州 510655 3. 暨南大學生命科學技術學院, 廣州 510632
為了研究流速對不同浮游藻類生長的影響, 于2015年3月采集東江流域廣東惠州河段原水, 在環形有機玻璃水槽中分別培養銅綠微囊藻、斜生柵藻和小環藻, 通過調整水體流速, 研究不同流速對不同藻類的細胞密度、葉綠素a濃度、最大比增長率和葉綠素熒光參數Fv/Fm的影響。結果表明, 在不同流速條件下, 不同浮游藻類的生理生化特征有所差異; 銅綠微囊藻表現為在不同流速條件下生長狀況都較弱; 斜生柵藻和小環藻在低流速(<0.075 m·s–1)條件下繁殖速度較快, 且隨著水體流速的增加, 對藻類生長有促進作用, 而在較高流速(>0.075 m·s–1)條件下隨著流速的增加, 藻類的生長受到抑制。可見, 不同持續的流動條件是影響浮游藻類數量和生理指標變化的重要原因。該研究為東江水源的流速流量調節控制富營養化和水華防治提供技術支撐。
東江; 流速; 浮游藻類; 富營養化; 水華
1 前言
由于高強度人類活動影響下大量氮磷營養鹽進入水環境, 造成水體富營養化和水華頻繁發生已嚴重損害了水生態系統服務功能[1]。水華的暴發與藻類的生理特點、營養鹽、溫度、光照、緩慢的水流條件及其諸多環境因素密切相關[2]。其中, 水文水動力的影響尤為顯著[3-5], 水利工程的開發導致水體從河流型向水庫型改變, 大壩攔截成庫造成局部河段流速減緩, 引起營養鹽的累積和藻類大量繁殖[6-7]。如長江三峽大壩截流后, 香溪河庫灣由天然河流變為人工水庫, 引起庫區水生態系統變化暴發甲藻水華[8]; 福建省九龍江北溪江自東引水工程運行后, 2009年曾發生甲藻水華事件[9]; 丹江口水庫大壩建成后, 導致漢江浮游植物群落組成以及細胞密度均發生顯著變化, 1992年初春漢江下游江段發生硅藻水華[10]。
流速作為水動力條件中最基本、最直觀的參數, 對浮游植物的生長和繁殖具有十分重要的影響[11]。研究表明[12-14]低流速有利于維持水環境的穩定、增加光照利用率和延長水力停留時間, 促進懸浮藻類的生長, 而過高的流速則抑制懸浮藻類的生長, 并且較大流速能有效地阻止水華的形成與發展, 消除已經形成的水華。因此, 通過流量管理和水力調度等方式控制富營養化和藻類水華已在河流水環境管理策略中得到廣泛應用[15-16]。
東江是珠江水系三大河流之一, 是河源、惠州、東莞、深圳等地的主要供水水源, 并承擔著向香港地區供水的重要任務[17]。近年來由于流域內經濟發展、城市化加速導致水體污染物增加, 加之流域內的梯級開發, 東江部分河段水質下降明顯, 干流富營養化程度從上游至下游呈逐漸加重趨勢[18-19]。東江干流已開發的梯級電站13座, 最下游的一個梯級為東江水利樞紐(劍潭梯級), 東江下游污染物的遷移擴散主要受該梯級下泄流量的影響[20], 劍潭梯級也是距離下游東深供水取水口最近的一個梯級。因此, 劍潭梯級上下游的水環境質量對東深供水水質的影響較大。研究表明[21]劍潭水庫富營養化程度已較蓄水初期有加劇趨勢, 已有潛在的藻類水華風險, 且水流流速是影響東江干流浮游植物生長的重要因素[22]。為掌握流速對東江藻類水華發生的影響, 本研究采集東江惠州河段原水開展藻類室內培養模擬試驗, 模擬東江水體中浮游藻類生長對流速的響應, 討論不同流速梯度下浮游藻類的生長特征及其變化規律, 探討抑制藻類水華暴發的臨界流速, 為東江流域水華風險防控提供數據支撐。
2 材料與方法
2.1 實驗裝置及流速設定
本研究采用5個環形有機玻璃水槽(圖1), 水槽長寬高尺寸為1150 mm×650 mm×500 mm。將水槽放置于25 ℃恒溫室內, 光照為日光燈照射, 水槽上方光照強度1500 lx。根據2015年3月12日在東江劍潭庫區(壩首至庫尾)現場實測的流速大小0.077—0.15 m·s–1以及東江的水流特征, 設置5個流速梯度(1#、2#、3#、4#和5#), 分別為0、0.045、0.075、0.135和0.25 m·s–1, 流速大小通過循環水真空泵調節控制。
2.2 實驗材料
實驗藻種選取藍藻門的銅綠微囊藻(FACHB-905)、綠藻門的斜生柵藻(FACHB-416)和硅藻門的小環藻sp.(FACHB-1635), 以上藻種均購自中國科學院淡水藻種庫。
2.3 實驗方法
(1)實驗用水取自東江劍潭水庫大壩處的原水, 用25#浮游植物網濾去大型浮游動物, 再用0.45 μm醋酸纖維濾膜進行抽濾后, 分別注入5個水槽, 水槽中水深為30 cm, 水量170 L。測定水槽中水體理化指標初始值, 根據水體氮磷營養鹽濃度, 向水槽中添加濃度為15 g·L–1的NaNO3和3 g·L–1的KH2PO4, 用于調節各水槽水體中總氮、總磷濃度, 分別達到3.0 mg·L–1和0.19 mg·L–1。
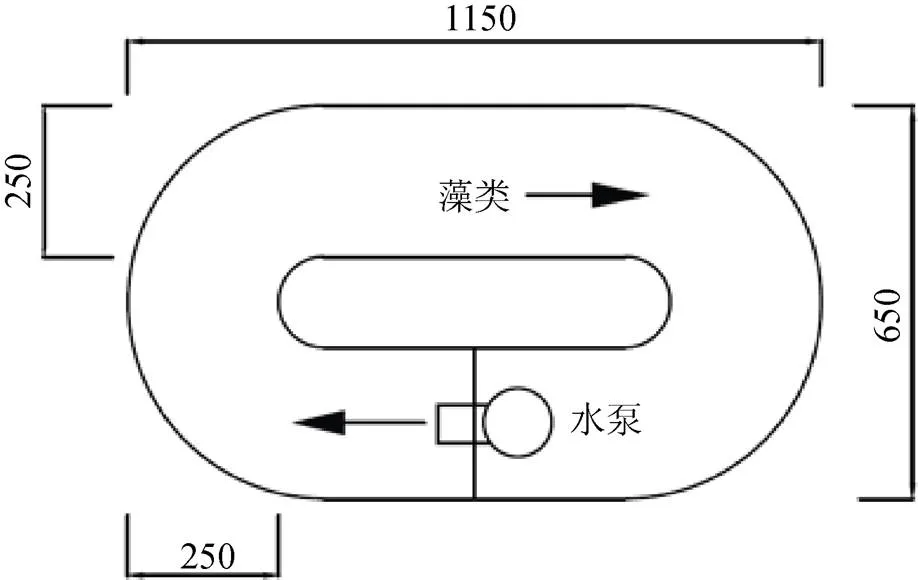
圖1 實驗裝置平面示意圖 (單位mm)
(2)將室內培養至對數生長期的藻種, 收集置于50 mL離心管, 離心收獲藻液, 用15 mg·L–1NaCO3溶液洗滌后再次離心, 棄去上清液, 重復兩次, 以除去附著在藻細胞上的營養鹽, 預處理后轉接至實驗水槽內。
(3)實驗期間隔天測量氮、磷營養鹽濃度, 保證裝置中氮、磷營養鹽濃度維持在高富營養水平, 每天早上(9:00)和晚上(21:00)分別監測1次, 記錄溶解氧(DO)、pH等, 并測量藻細胞密度、葉綠素a和葉綠素熒光參數(Fm、Fv)直到實驗結束。
2.4 樣品分析
pH和DO用便攜式水質分析儀測定, 總氮(TN)、總磷(TP)和葉綠素a的測定均參考《水和廢水監測方法(第4版)》, 顯微鏡(OLYMPUS CX21)下浮游藻類細胞數量計數方法參照《淡水浮游生物研究方法》[23], 葉綠素熒光參數用浮游植物熒光儀(WATER-PAM)測定, Fv/Fm是可變熒光與最大熒光的比值, 反映藻類PSII的活性。
2.4 數據處理
最大比增長速率()是在某一時間間隔內藻類生長的速率。其計算公式為:
=ln (X/X)/(t-t) (1)
式中:X為某一時間間隔開始時的藻類現存量(cells·mL–1);X為某一時間間隔結束時的藻類現存量(cells·mL–1);t-t: 為某一時間間隔(d)。
使用Origin Lab8.5對試驗數據進行圖形分析;使用SPSS17.0統計軟件進行方差分析(ANOVA), 顯著性水平為0.05。
3 結果與分析
3.1 流速對不同藻類生長過程中pH和DO的影響
不同流速條件下銅綠微囊藻、斜生柵藻和小環藻生長過程中pH的變化整體均呈現上升趨勢, 其中銅綠微囊藻和斜生柵藻的pH值變化范圍較接近, 分別為6.82—7.83、6.66—7.69; 小環藻的pH值相對較高, 變化范圍為7.21—8.24(圖2)。流速組(2#—5#)銅綠微囊藻生長期內pH均高于靜止組(1#); 斜生柵藻和小環藻生長的水體中, 有流速的條件下隨著流速增加pH值升高, 而靜止組pH波動明顯, 在小環藻生長的中后期靜止培養組pH值高于流速組。
不同流速條件下銅綠微囊藻生長過程中DO呈先上升后降低的變化趨勢, 變化范圍為7.45—8.42 mg·L–1, 流速組DO高于靜止培養組(1#); 斜生柵藻和小環藻生長過程中DO變化呈上升趨勢, 且在生長的中后期靜止培養組的DO升高較快, 最高值分別達到9.53 mg·L–1和10.65 mg·L–1, 高于流速組(圖2)。
3.2 流速對不同藻類生長速率的影響
不同流速條件下初始接種的銅綠微囊藻細胞密度平均值為6.29×105cells·L–1, 在生長過程中不同流速組銅綠微囊藻細胞密度呈現先上升后降低趨勢(圖3)。在流速為0.135 m·s–1和0.25 m·s–1條件下培養至第3 d時, 銅綠微囊藻細胞密度數量高于其他各流速組和靜止組, 至培養的第4 d, 各流速組和靜止組的銅綠微囊藻細胞密度數量均大幅下降。不同流速條件下銅綠微囊藻生長中葉綠素a濃度與藻細胞密度變化趨勢較為一致, 至培養的第4 d, 葉綠素a濃度已由第1 d的平均值3.42 μg·L–1下降為1.18 μg·L–1。結合顯微鏡下觀察, 以上結果反映出銅綠微囊藻在培養過程中生長狀況較差, 至培養的第4 d藻細胞開始大量死亡。

圖2 不同流速下pH、DO變化趨勢
不同流速條件下初始接種的斜生柵藻細胞密度平均值為0.72×105cells·L–1, 斜生柵藻在生長過程中靜止組的細胞密度和葉綠素a濃度均高于流速組, 最低流速0.045 m·s–1條件下藻細胞密度和葉綠素a濃度一直處于較低水平, 其他各流速組藻細胞密度和葉綠素a濃度波動較大(圖3)。
不同流速條件下初始接種的小環藻細胞密度平均值為2.31×105cells·L–1, 在生長的第5 d小環藻生長開始進入穩定期, 且該時期0.135 m·s–1和0.25 m·s–1高流速組的藻細胞密度和葉綠素a濃度低于0.045 m·s–1、0.075 m·s–1流速組和靜止培養組(圖3)。
3.3 流速對藻類最大比增長速率的影響
不同流速條件下銅綠微囊藻、斜生柵藻和小環藻的最大比增長率如圖4所示, 銅綠微囊藻靜止組與流速組最大比增長率呈現明顯差異(<0.05), 靜止組在生長過程中細胞密度很快下降至低于初始接種密度水平, 導致最大比增長速率為負值, 流速組最大比增長率相對較低且呈相同的變化趨勢。斜生柵藻在流速(≤0.045)和流速(≥0.075)呈現明顯差異(<0.05), 且靜止組的最大比增長速率最高, 流速由0.045 m·s–1增加至0.075 m·s–1時, 斜生柵藻最大比增長速率下降幅度較大, 說明流速的增加對斜生柵藻的生長產生一定的抑制作用。小環藻的最大比增長率在不同流速組差異不明顯, 且在流速為0.075 m·s–1時, 最大比增長率相對較高, 流速>0.075 m·s–1時最大比增長速率下降, 說明高流速也不利于小環藻的生長。
3.4 流速對不同藻類葉綠素熒光參數Fv/Fm的影響
不同流速下銅綠微囊藻、斜生柵藻、小環藻的Fv/Fm值變化如圖5所示。銅綠微囊藻初始Fv/Fm值約為0.3, 在生長期內銅綠微囊藻的Fv/Fm值逐漸降低接近為零, 說明水體中銅綠微囊藻光合系統PSII活性很低。在生長期內不同流速條件下的斜生柵藻Fv/Fm均逐漸降低, 小環藻Fv/Fm則呈現先上升至穩定然后又下降的趨勢, 且靜止組(1#)斜生柵藻和小環藻的Fv/Fm均高于流速組(2#—5#)。
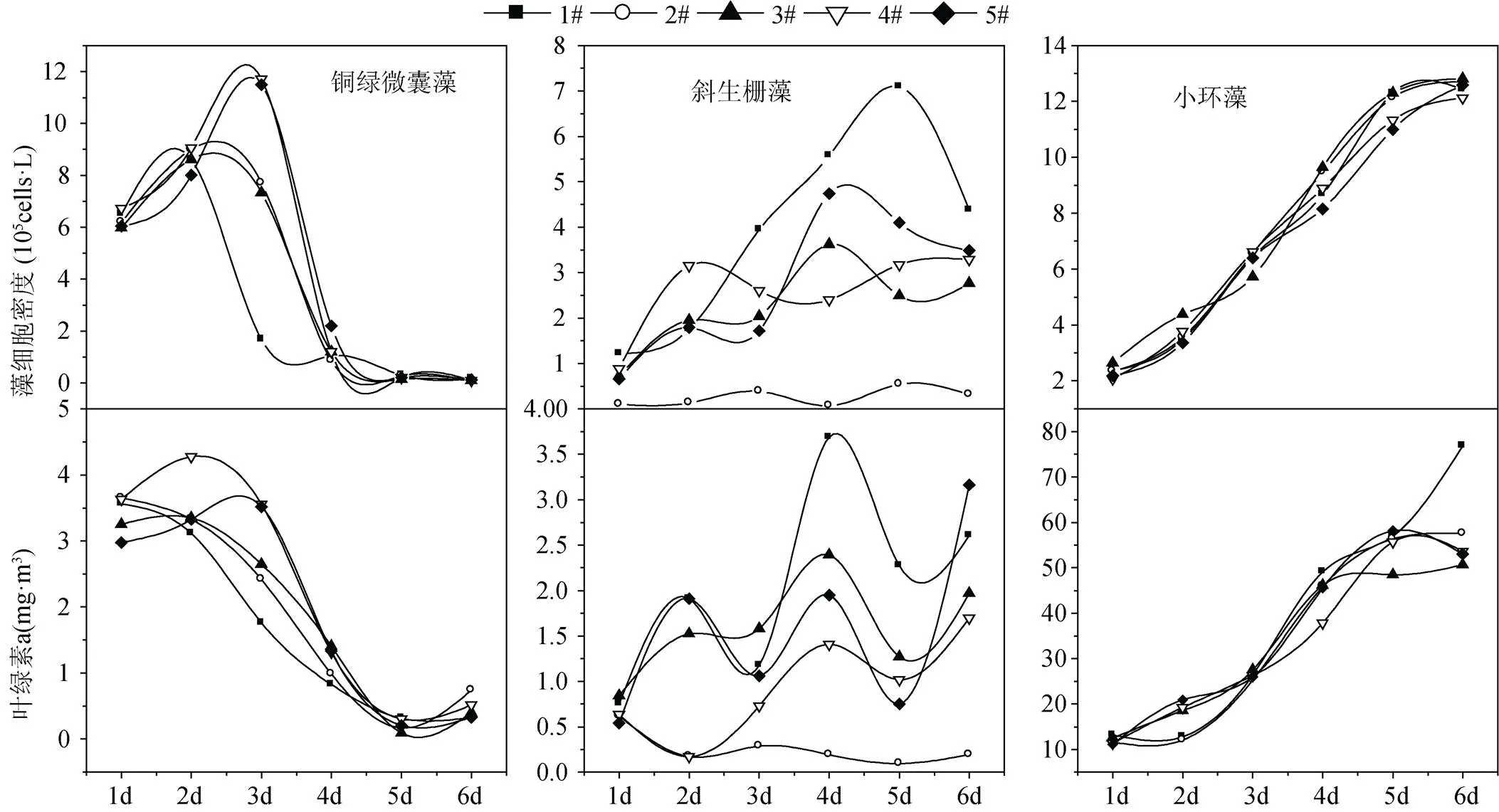
圖3 不同流速下藻細胞密度和葉綠素a的變化情況

注:字母a, b標識最大比增長率在不同流速之間顯著性差異比較, 采用SPSS17.0進行方差分析, 顯著水平0.05。

圖5 不同流速下藻類Fv/Fm的變化趨勢
4 討論
4.1 東江水體發生藻類水華的風險分析
東江作為流域內河源、惠州、東莞、深圳、廣州等城市以及香港約4000多萬人口的主要飲用水源, 其水環境保護一直備受重視, 流域內采取了系列嚴格的水環境保護措施, 尤其是東江流域上游, 為保護好東江優質水資源在經濟發展方面做出了巨大的犧牲[24]。盡管東江干流水質整體優良, 然而由于流域內人口增加和經濟的快速發展[25], 干流仍有少數區域達到富營養化水平[18]; 東江干流TN含量均值為1.99 mg·L–1, TP含量變化范圍為0.01—0.28 mg·L–1, 且越靠近下游TN、TP含量越高, 水體富營養化程度越高[22], 特別是東江下游劍潭水庫內較高的營養鹽已為浮游植物大量生長繁殖創造了有利條件, 東江下游潛在的水華風險受到關注[21]。
近年來我國河流藻類水華發生呈增加趨勢, 與湖泊水華相比, 水動力條件的改變是河流中浮游藻類優勢種變化的最主要影響要素, 也是河流中影響浮游藻類生長的主要限制因素之一[26]。不同藻類的臨界流速不同[11], 澳大利亞達令河卷曲魚腥藻大量暴發的臨界流速為 0.05 m·s–1[14]; 運用非恒定二維生態動力學模型研究發現嘉陵江藻類水華發生的臨界流速為0.04 m·s–1[27]; 室外試驗研究發現小球藻和纖維藻生長的臨界流速為0.05 m·s–1和0.01 m·s–1[28]; 室內實驗表明低流速(0.05—0.12 m·s–1)有利于銅綠微囊藻生長, 而在靜止與高流速條件下 (0.20—0.30 m·s–1)藻類的生長受到抑制[29]。除嫌水藻類外, 中流藻類和喜流水藻類都可以在流水中生長, 由于水體流動使藻類不斷得到新的營養物質的供應, 水流對藻類的生長和繁殖是有利的[30]。微囊藻屬嫌流水藻類[31], 較高的流速對微囊藻的生長有明顯的抑制作用, 但較緩慢的流速在一定程度上有利于微囊藻的生長。本研究發現在靜水、小流速和較大流速條件下, 銅綠微囊藻生長速率均較低, 與實驗所用的銅綠微囊藻活性較低有關, 實驗初始銅綠微囊藻的Fv/Fm僅為0.263—0.321; 其次, 研究發現東江劍潭水庫中微囊藻并不是優勢種, 其藻細胞密度較低[21];此外, 根據實驗氮磷營養鹽濃度水平, 各裝置在實驗第2d氮磷濃度有所降低, 說明微囊藻生長消耗了營養鹽, 而從第3d開始, 氮磷濃度基本保持不變, 可見水體中微囊藻已不再消耗營養鹽。以上結果表明目前東江干流劍潭水庫中并不適宜微囊藻的大量生長, 由此可以判斷東江劍潭水庫發生微囊藻水華的可能性較低。斜生柵藻作為綠藻的一種, 以小流速下生長狀況最好, 而較高的流速不利于其生長。斜生柵藻在流速大于0.02 m·s–1時即產生明顯的抑制生長作用[33]。硅藻是形成河流水華的主要種類之一, 多發生在天然河流成庫后的淹沒回緩流水庫灣或支流。小環藻為喜流水藻類, 適宜生活于較低的水流速度, 而對較高的水流速度耐受較弱[31]。近年來漢江暴發硅藻水華(優勢種為小環藻)時的河道流速為0.07 m·s–1左右[32]。本研究發現小環藻在流速為0.075 m·s–1時最大比增長速率較高, 在相對較高的流速(0.135和0.25 m·s–1)時, 小環藻的生長開始出現抑制, 說明當東江干流劍潭水庫水流大于0.075 m·s–1時, 隨著流速的增加小環藻生長受到抑制; 斜生柵藻也是在流速大于0.075 m·s–1時最大比增長速率有所降低, 以上結果表明劍潭水庫水體流速增加至0.075 m·s–1以上將對藻類生長產生抑制作用。
4.2 東江水體水華防控措施
作為快速有效改善水庫型河流和水庫庫區水環境的手段之一, 通過改善水利條件從而減少水華暴發風險的研究越來越受到重視。Maier等[12]研究發現通過水利調節打破水體熱分層、減少水體滯留時間, 可以消除澳大利亞Darling河的魚腥藻水華; 辛小康等[35]通過建立平面二維水動力數學模型發現, 利用干支流水庫對香溪河庫灣流速的聯合調控可適度控制三峽庫區支流庫灣水華的暴發; 殷大聰等[36]研究提出漢江中下游春季硅藻水華暴發的關鍵水文閾值及生態調度控制策略; 劉德富等[37]研究提出用于防控三峽支流水華的 “潮汐式”生態調度方案。東江作為香港、深圳及廣州等特大城市的重要水源地, 水庫調度是有效解決流域突發性水污染的重要措施, 同時由于新豐江、楓樹壩與白盆珠三大水庫對東江全流域水資源有控制性調蓄作用, 東江具有開展水利調度的有利條件, 已有研究[20,38]表明利用東江水利樞紐進行水量調度可有效改善東江干流水質。本研究結果發現銅綠微囊藻在東江水體中長勢較弱, 發生水華的可能性較低, 而斜生柵藻和小環藻均在較低流速<0.075 m·s–1時生長速度快, 且隨著流速增大, 對藻類生長有促進作用, 容易發生水華, 而在較高流速>0.075 m·s–1條件下, 隨著流速的增加, 藻類的生長受到抑制。因此, 針對當前枯水期東江存在水華的風險, 及實驗結果東江水華發生臨界流速為0.075 m·s–1, 并結合東江惠州段現場實測流速, 確定東江劍潭水庫防控水華的臨界流速為0.07—0.08 m·s–1。在水華易發的氣象條件下, 可通過流域內水庫聯合調度以保證東江的生態流速, 從而有效防止水華的發生。
5 結論
(1)實驗中斜生柵藻臨界流速值約為0.075 m·s–1; 銅綠微囊藻在不同流速條件下生長都較弱; 小環藻對流速響應關系為在流速>0.075 m·s–1時藻類生長受到抑制。因此, 東江流域惠州劍潭水庫大壩處浮游藻類生長的臨界流速為0.075 m·s–1。
(2)實驗中不同藻類對流速的響應適應性具有明顯差異, 并且流速條件的改變對藻類的生物量和生理生化特征產生較大影響。因此, 若要通過水利調度來預防和控制水華, 需視具體藻類來優化水利調度方案。
[1] SMITH V H. Eutrophication of freshwater and coastal marine ecosystems a global problem [J]. Environmental Science and Pollution Research, 2003, 10(2): 126–39.
[2] MORSE R E, SHEN J, BLANCO-GARCIA J L, et al. Environmental and physical controls on the formation and transport of blooms of the DinoflagellateMargalef in the lower Chesapeake Bay and its tributaries [J]. Estuaries & Coasts, 2011, 34(5): 1006–1025.
[3] HA K, JANG M H, JOO G J. Winterbloom development in the Nakdong River regulated by an estuary dam and tributaries [J]. Hydrobiologia, 2003, 506-509(1-3): 221–227.
[4] 張遠, 夏瑞, 張孟衡, 等.水利工程背景下河流水華暴發成因分析及模擬研究[J].環境科學研究, 2017, 30(8): 1163–1173.
[5] LIU Xia, LI Yunliang, LIU Baogui, et al. Cyanobacteria in the complex river-connected Poyang Lake: horizontal distribution and transport [J]. Hydrobiologia, 2016, 768(1): 95–110.
[6] 程輝, 吳勝軍, 王小曉, 等.三峽庫區生態環境效應研究進展[J].中國生態農業學報, 2015, 23(2): 127–140.
[7] 梁培瑜, 王烜, 馬芳冰.水動力條件對水體富營養化的影響[J].湖泊科學, 2013, 25(4): 455–462.
[8] ZHOU Guangjie, ZHAO Xuemin, BI Yonghong, et al. Phytoplankton variation and its relationship with the environment in Xiangxi Bay in spring after damming of the Three-Gorges, China [J]. Environmental Monitoring & Assessment, 2011, 176: 125–141.
[9] 邊歸國, 陳寧, 胡征宇, 等.福建某河流甲藻水華與污染指標的關系[J].湖泊科學, 2010, 22(3): 405–410.
[10] 潘曉潔, 朱愛民, 鄭志偉, 等.漢江中下游春季浮游植物群落結構特征及其影響因素[J].生態學雜志, 2014, 33(1): 33–40
[11] 吳曉輝, 李其軍.水動力條件對藻類影響的研究進展[J].生態環境學報, 2010, 19(7): 1732–1738.
[12] MAIER H R, BURCH, M D, BORMANS M. Flow management strategies to control blooms of the cyanobacterium,, in the river Murray at Morgan, South Australia [J]. Regulated Rivers Research & Management, 2001, 17(6): 637–650.
[13] ACUNA V, VILCHES C, GIORGI A. As productive and slow as a stream can be——the metabolism of a Pampean stream [J]. Journal of the North American Benthological Society, 2011, 30(1): 71–83.
[14] MITROVIC S M, OLIVER R L, REES C, et al. Critical flow velocities for the growth and dominance ofin some turbid freshwater rivers [J]. Freshwater Biology, 2003 48(1): 164–174.
[15] 陳瑞弘, 李飛鵬, 張海平, 等.面向流量管理的水動力對淡水藻類影響的概念機制[J].湖泊科學, 2015, 27(1): 24–30.
[16] MITROVIC S M, HARDWICK L, DORANI F. Use of flow management to mitigate cyanobacterial blooms in the lower Darling River, Australia [J]. Journal of Plankton Research, 2011, 33(2), 229–241.
[17] YAN Liang, FUNG Puika, MAN Fungtse, et al. Sources and seasonal variation of PAHs in the sediments of drinking water reservoirs in Hong Kong and the Dongjiang River (China) [J]. Environmental Monitoring & Assessment, 2008, 146: 41–50.
[18] 王博, 梁志誠, 任斐鵬, 等.東江干流富營養化評價及區域差異性研究[J].環境污染與防治, 2011, 33(6), 15–18.
[19] 廖劍宇, 彭秋志, 鄭楚濤, 等.東江干支流水體氮素的時空變化特征[J].資源科學, 2013, 35(3): 505–513.
[20] 江濤, 鐘鳴, 鄒隆建, 等.石馬河泄洪與東江水利樞紐調節不同情景下東江水質的模擬與分析[J].中山大學學報(自然科學版), 2016, 55(2): 117–123.
[21] 胡芳, 許振成, 姚玲愛, 等.劍潭水庫浮游植物群落特征與水環境因子關系研究[J].環境科學學報, 2014, 34(4): 950–958.
[22] 田時彌, 楊揚, 喬永民, 等.珠江流域東江干流浮游植物葉綠素a時空分布及與環境因子的關系[J].湖泊科學, 2015, 27(1): 31–37.
[23] 章宗涉, 黃祥飛.淡水浮游生物研究方法[M].北京: 科學出版社, 1991.
[24] 劉強, 彭曉春, 周麗旋, 等.城市飲用水水源地生態補償標準測算與資金分配研究——以廣東省東江流域為例[J]. 生態經濟, 2012, (1): 33–37.
[25] GAO Chanjun, ZHOU Ping, JIA Peng, et al. Spatial driving forces of dominant land use/land cover transformations in the Dongjiang River watershed, Southern China [J]. Environmental Monitoring & Assessment, 2016, 188(2): 84.
[26] 陳永燦, 俞茜, 朱德軍, 等.河流中浮游藻類生長的可能影響因素研究進展與展望[J].水力發電學報, 2014, 33(4): 186–195.
[27] LONG Tianyu, WU Lei, MENG Guohu, et al. Numerical simulation for impacts of hydrodynamic conditions on algae growth in Chongqing Section of Jialing River, China [J]. Ecological Modelling, 2011, 222(1): 112–119.
[28] 焦世珺.三峽庫區低流速河段流速對藻類生長的影響[D].重慶: 西南大學, 2007.
[29] 王華, 逄勇.藻類生長的水動力學因素影響與數值仿真[J].環境科學, 2008, 29(4): 884–889.
[30] 福迪. 藻類學[M].羅迪安, 譯.上海: 上海科學技術出版社, 1980: 392–394.
[31] 楊敏, 畢永紅, 艾鷹, 等.人工控制條件下水流速對香溪河庫灣浮游植物影響的初步研究[J]. 長江流域資源與環境, 2012, 21(2): 220–224.
[32] 盧大遠, 劉培剛, 范天俞, 等.漢江下游突發“水華”的調查研究[J].環境科學研究, 2000, 13(2): 28–31.
[33] 王建慧.流速對藻類生長影響試驗及應用研究[D].北京: 清華大學, 2012.
[34] 邊歸國.九龍江擬多甲藻水華預警和應急處置的初步研究[J].中國環境管理, 2013, 5(3): 30–34.
[35] 辛小康, 尹煒, 葉閩.水動力調控三峽庫區支流水華方案初步研究[J].水電能源科學, 2011, 29(7): 16–18.
[36] 殷大聰, 尹正杰, 楊春花, 等.控制漢江中下游春季硅藻水華的關鍵水文閾值及調度策略[J].中國水利, 2017, (9): 31–34.
[37] 劉德富, 楊正健, 紀道斌, 等.三峽水庫支流水華機理及其調控技術研究進展[J].水利學報, 2016(3): 443–454.
[38] 黃國如, 李春.東江干流突發性重金屬污染的水庫調度效應研究[J].水電能源科學, 2013(6): 82–85.
The effect of flow velocity on the growth of different phytoplankton
ZHOU Jing1,2, GOU Ting2, ZHANG Luohong1, LAN Yu3, MA Qianli2, LIANG Rongchang2, ZHAO Xuemin2,*
1. School of Environmental and Chemical, Xi’an Polytechnic University, Xi’an 710048, China 2. South China Institute of Environmental Sciences, Ministry of Environmental Protection, Guangzhou 510655, China 3. College of Life Science and Technology, Jinan University, Guangzhou 510632, China
To study the effect of flow velocity on the growth of different phytoplankton, a simulation experiment was conducted in the annular flume of organic glass using,andsp., which were cultured in laboratory using surface water from the Huizhou reach of Dongjiang River in March 2015. The effects of different flow velocity on the algal cell density, the concentration of chlorophyll, the maximum specific growth rate and chlorophyll fluorescence parameter (Fv/Fm) were studied by adjusting the flow rate of water. The results showed that the physiological and biochemical characteristics varied at different flow rates. The growth ofwas slow with all flow velocity conditions, the growth ofandsp. was fast at low flow rate (<0.075m·s–1) and increased with the increase of flow velocity. However, under the condition of high velocity (<0.075m·s–1), the growth of algae was inhibited with the increase of flow velocity. The different continuous flow may directly induce the changes of the cell density and physiological indicators of phytoplankton, indicating the control of eutrophication and algae bloom by adjusting flow velocity in Dongjiang water.
Dongjiang River; flow velocity; phytoplankton; eutrophication; algal bloom
10.14108/j.cnki.1008-8873.2018.06.010
X522
A
1008-8873(2018)06-075-08
2018-06-11;
2018-08-02
廣州市科技計劃產學研協同創新重大專項(201704030110); 中央級公益性科研院所基本科研業務費專項(PM-zx703-201602-048); 國家水體污染控制與治理科技重大專項(2014ZX07206-005-04)聯合資助
周靜(1992—), 女, 碩士研究生, 主要從事水環境污染防治, E-mail:1602111799@qq.com
趙學敏, 博士, 研究員, 主要從事水生態風險評估與控制研究, Email: zhaoxuemin@scies.org
周靜, 茍婷, 張洛紅, 等. 流速對不同浮游藻類的生長影響研究[J]. 生態科學, 2018, 37(6): 75-82.
ZHOU Jing, GOU Ting, ZHANG Luohong, et al. The effect of flow velocity on the growth of different phytoplankton[J]. Ecological Science, 2018, 37(6): 75-82.
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
