菠菜抗壞血酸過氧化物酶基因家族的鑒定及表達分析
2019-01-14 04:41:14郁征宇葛晨輝王小麗徐晨曦王全華蔡曉鋒
上海師范大學學報·自然科學版 2018年6期
郁征宇, 葛晨輝, 王小麗, 徐晨曦, 王全華, 蔡曉鋒
(上海師范大學 生命科學學院 植物種質資源開發協同創新中心,上海 200234)
0 引 言
當植物受到高溫、低溫、干旱和鹽等非生物脅迫時,其體內會產生大量的O2-、HO-、過氧化氫(H2O2)等活性氧(ROS)[1].即使是在正常的條件下,植物體內的許多代謝過程,包括葉綠體、線粒體和質膜相關的電子傳遞系統也會釋放出活性氧[2].當活性氧清除系統無法及時去除細胞中產生的活性氧時,就會造成植物細胞層面上的氧化損傷[3].植物細胞內存在酶類和非酶類兩種清除活性氧的抗氧化系統,其中抗氧化物酶類系統中包括了超氧化物歧化酶(SOD),抗壞血酸過氧化物酶(APX)、過氧化氫酶(CAT)、谷胱甘肽還原酶(GR)等,而對去除過氧化氫起主要作用的是CAT和APX,APX是抗壞血酸-谷胱甘肽(AsA-GSH)循環中的關鍵酶,對H2O2的親和力更強[4],以抗壞血酸(ASA)為電子供體,催化H2O2的還原[5].APX是保護葉綠體免受H2O2氧化損傷的主要酶類.有大量研究證實了APX在植物應對非生物脅迫中起著重要作用.
APX主要存在于高等植物中,在某些真核藻類、藍細菌和昆蟲中也檢測到了APX[6].通過亞細胞定位研究,將APX同工酶分為4種類型:細胞質型、葉綠體型、線粒體型和微體型[7].在擬南芥和水稻中都檢測到了8種APX同工酶,擬南芥APX有3個位于細胞質,2個位于葉綠體,另外3個位于過氧化物酶體中[8];水稻APX中的2個存在于細胞質,3個存在于葉綠體,2個存在于過氧化物酶體,另外1個結合在線粒體上[9].
菠菜(SpinaciaoleraceaL.)為耐寒性蔬菜,含有豐富的營養,生長溫度高于25 ℃時,其品質和產量都會受到影響,嚴重時甚至死亡.高溫、干旱和鹽含量的急劇變化都會導致活性氧的大量累積,APX在清除活性氧中的關鍵作用使得探討菠菜APX基因的表達調控具有重要的理論意義和實踐價值.本文作者對菠菜APX進行了生物信息學的分析,并研究了其組織表達和環境脅迫調控能力.
1 材料與方法
1.1 菠菜APX基因的鑒定
在擬南芥種質信息(TheArabidopsisInformation Resource,TAIR) 數據庫(http://www.arabidopsis.org)中獲得擬南芥AtAPXs(Ascorbate Peroxidases)蛋白序列,在菠菜數據庫(http://www.spinachbase.org/?q=blast)中進行TBLASTN分析得到菠菜APX基因候選序列,同時對所獲得的序列開放閱讀框(ORF),利用GENSCAN (http://genes.mit.edu/GENSCAN.html)預測確認.經篩選后的氨基酸序列再利用ScanProsite軟件(http://www.expasy.ch/tools/seanprosite/)和InterProScan軟件(http://www.ebi.ac.uk/Tools/InterProScan/)來驗證是否含有APX基因保守序列.
1.2 基因結構、蛋白保守序列及進化樹分析
利用基因核苷酸編碼序列與其相對應的基因組序列,通過Gene Structure Display Server (GSDS 2.0,http://gsds.cbi.pku.edu.cn/index.php)確定其基因結構.利用Protpi (https://www.protpi.ch/Calculator/ProteinTool#ProteinInputs)預測氨基酸數量、分子量和等電點.使用ClustalW進行多重序列比對,MEGA5[10]繪制進化樹(NJ tree).利用MEME suite (http://meme-suite.org/tools/meme)進行蛋白質保守基序(motif)分析,通過p-value判斷結果是否達到顯著水平.在線使用WoLF PSORT (http://www.genscript.com/psort/wolf_psort.html)對其蛋白質序列進行亞細胞定位.
1.3 菠菜材料的處理與采樣
選取S104菠菜為實驗材料,將種子播種于穴盤,以體積比例為1∶1的草炭土/珍珠巖為基質,在玻璃溫室中育苗.選取長勢一致的幼苗,小心洗去根部基質,移至營養液(大量元素采用Hoagland配方,微量元素采用Arnon配方)中進行水培,營養液每3 d進行一次更換,并且每天對其pH值進行測量與調節,使其保持在5.8~6.0之間.水培菠菜置于人工氣候室中,光周期為10 h光照/14 h黑暗,溫度控制在21 ℃(光照)/18 ℃(黑暗),光照強度約為5000 lx,濕度控制在71%左右.水培兩周后,選取長勢一致的菠菜苗,準備進行環境脅迫處理.采用添加20%(質量分數)PEG 4000的方式進行干旱模擬;通過添加物質的量濃度為400 mmol·L-1的NaCl溶液模擬鹽脅迫環境;噴施物質的量濃度為100 mmol·L-1的H2O2模擬氧化脅迫;調節人工氣候箱的溫度模擬高溫脅迫(40 ℃)及低溫脅迫(4 ℃).分別在脅迫處理后的0,3,6,12 h時對菠菜功能葉進行采樣.
選取長勢一致的幼苗定植于自然條件光照的玻璃溫室中.在菠菜植株開花期采集不同發育階段的根、莖、功能葉、功能葉葉柄、新葉、新葉葉柄、雄花和雌花樣品,并立即于-80 ℃液氮中冰凍保存.
1.4 APX基因表達分析
使用Trizol法提取菠菜總核糖核酸(RNA),并使用NanoDropTMOneC分析RNA的濃度與質量.通過PrimeScriptTMRT reagent Kit with gDNA Eraser (Perfect Real Time)試劑盒將提取的總RNA反轉錄成cDNA(Complementary DNA)模板.使用Primer-BLAST (https:www.ncbi.nlm.nih.gov/tools/primer-blast)進行引物設計.使用TB GreenTMPremix Ex TaqTM(Tli RNaseH Plus) 試劑盒在ABI 7500上進行實時熒光定量聚合酶鏈式反應(qRT-PCR).采用So18s基因作為內參,用2-ΔΔCt法計算相對表達量,用GraphPad Prism 7對結果進行作圖.
2 結 果
2.1 菠菜SoAPXs家族成員全基因組鑒定
在菠菜基因組數據庫中利用8個擬南芥AtAPX蛋白序列進行TBLASTN分析,經過保守序列確認得到7個菠菜SoAPXs基因,數量與擬南芥和水稻的8個相似.將這7個基因分別命名為SoAPX1,SoAPX2,SoAPX3,SoAPX4,SoAPX5,SoAPX6和SoAPX7.
菠菜SoAPXs基因的基因組核苷酸序列長度分布于755 bp(SoAPX7)到6688 bp(SoAPX2)之間.開放式閱讀框長度最短的是429 bp(SoAPX7),最長的為421 bp(SoAPX6),對應的蛋白質氨基酸數量分布于142~421之間.蛋白質相對分子量(MW)在1.554×104~4.572×104u之間,等電點(pI)最大為8.40(SoAPX6),最小為5.26(SoAPX7)(表1).亞細胞定位預測結果顯示SoAPX1,SoAPX3和SoAPX7蛋白定位于細胞質;SoAPX2、SoAPX5和SoAPX6蛋白定位于細胞周質;SoAPX4則定位于細胞外基質.
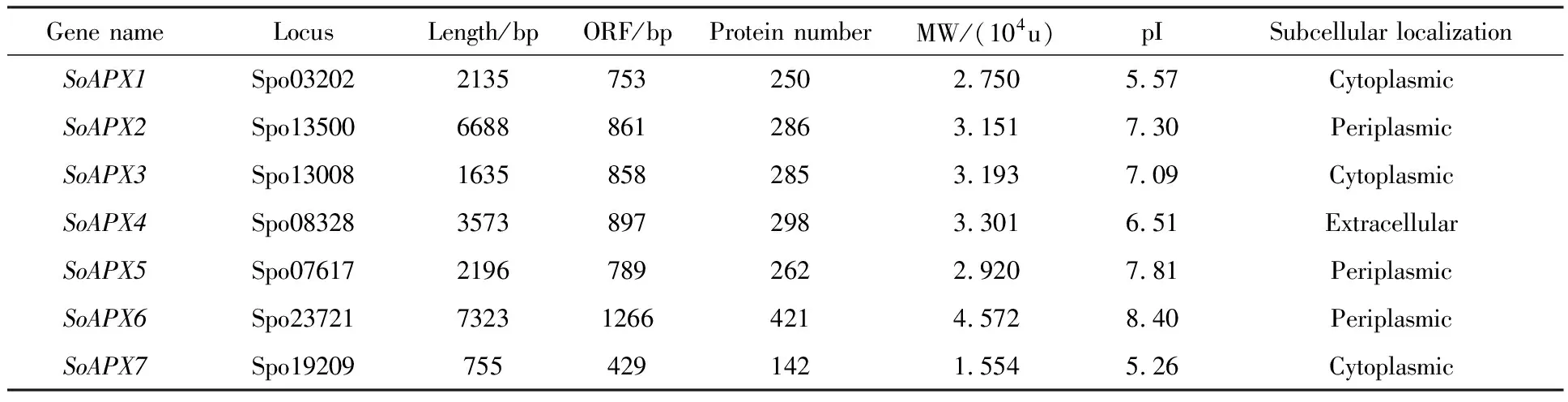
表1 菠菜SoAPX家族基因信息
菠菜SoAPXs基因的外顯子數量為5~12個(圖1),其中SoAPX2,SoAPX3,SoAPX4和SoAPX5都含有9個外顯子,8個內含子;SoAPX1含有8個外顯子,SoAPX6含有12個外顯子,SoAPX7含有5個外顯子.

圖1 菠菜SoAPX家族基因結構
2.2 菠菜SoAPX家族蛋白保守基序分析
利用Pfam對擬南芥和菠菜APX蛋白序列進行分析,結果顯示每個基因都含有過氧化物酶(peroxidase)的結構域.MEME對菠菜和擬南芥共15個APX蛋白質序列進行分析,共鑒定了9個保守基序,長度在11~50個氨基酸殘基之間(圖2).

圖2 菠菜SoAPX基因中保守基序的氨基酸序列
這些motifs的分布如圖3所示:每個APX蛋白中的motif數量在4~9之間,其中motif 4存在于所有蛋白中,其次是motif 1和motif 7,存在于除SoAPX6外的其余14個蛋白中,其中motif 7都位于C末端,出現次數最少的是motif 8,在SoAPX1,SoAPX2,SoAPX3,SoAPX4,SoAPX5,AtAPX1,AtAPX2和AtAPX5中出現,并且都位于N端.
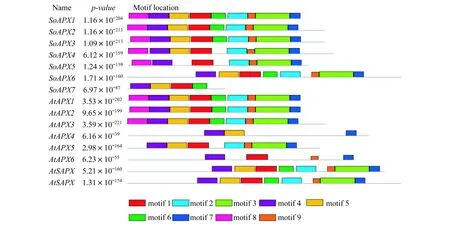
圖3 菠菜與擬南芥APX蛋白保守基序分布
2.3 菠菜和其他植物APX蛋白系統發育分析

圖4 菠菜及其他3種植物的APX蛋白進化樹分析
利用菠菜、擬南芥和水稻的APX蛋白質長序列構建了系統進化樹(圖4).結果表明AtAPX4和AtAPX6未被聚類,其他APX被聚為4個大類:SoAPX1與定位于細胞質的AtAPX1,AtAPX2,OsAPX1和OsAPX2為一類,SoAPX2,SoAPX3及SoAPX7與定位在過氧化物酶體上的AtAPX3,OsAPX3及OsAPX4聚為一類,SoAPX5和SoAPX4與定位在過氧化物酶體上的AtAPX5聚為一類,SoAPX6與定位于葉綠體上的AtSAPX,AtTAPX,OsAPX5,OsAPX8,以及OsAPX6,OsAPX7聚為一類.
2.4 菠菜SoAPX基因在各組織中的表達
利用菠菜的7個SoAPX基因特異引物(表2)進行qRT-PCR檢測,表達結果表明菠菜SoAPXs基因在所使用檢測部位中均有表達,除SoAPX2在根中表達較低外,其他6個基因在根中都有較高的表達(圖5).SoAPX7在各個組織中的表達較低,其次是SoAPX3.SoAPX7在雄花中表達最多,在功能葉、功能葉葉柄和莖中表達較低.SoAPX3主要在根、新葉和新葉葉柄中表達,在功能葉中表達較低.SoAPX4和SoAPX5的組織表達模式類似,在根部表達最多,其次是功能葉、功能葉葉柄,在莖中表達較低,在新葉、新葉葉柄、雄花和雌花中幾乎不表達.SoAPX1則與SoAPX3具有類似的表達模式,主要在新葉和新葉葉柄中表達,其次是根,在功能葉中的表達較低.SoAPX6除了在根中表達較高外,在其他各組織中均有類似的表達量(圖5).

表2 菠菜APX基因qRT-PCR引物

圖5 SoAPX基因在菠菜不同組織中的表達分析.(a)~(g) SoAPX1~SoAPX7,R:根;S:莖;L:功能葉;P:功能葉柄;YL:新葉;YP:新葉葉柄;FF:雌花;MF:雄花
2.5 菠菜SoAPX基因在不同環境脅迫下的表達
圖6為菠菜SoAPX基因經不同脅迫處理后的表達結果.由圖6可知,經低溫脅迫處理6 h后,各SoAPX基因的表達都出現了不同程度的上調,而在低溫脅迫12 h后,大部分基因的表達下調.SoAPX3和SoAPX6在低溫脅迫時長為3 h時上調,在時長為6 h和12 h時的表達則逐漸降低,特別當時長為12 h時其表達相比對照存在明顯的下調.SoAPX1,SoAPX2和SoAPX7在低溫脅迫時長為1,3,6 h時的表達呈上升趨勢,在6 h時表達量已達到對照的2.5倍,在處理12 h后的表達則下調.
冬天。然而,潛伏在這種表象之下的,是永不停滯的創造力。這座城市就像一個巨大的試驗場,永遠不會缺少新的生存形態。因此,理所當然的,我們選擇了為21世紀用車生活而生的XF SPORTBRAKE。

圖6 菠菜SoAPX基因對不同環境脅迫的響應.(a)~(g) SoAPX1~SoAPX7
經鹽脅迫處理后,在3 h時SoAPX3和SoAPX7的表達有明顯上升,表達量是對照的3.8倍和4.6倍,其他基因的表達也都有小幅的上調.SoAPX3和SoAPX6的表達僅在3 h時有所上調,其余處理時長的表達均下調表達,并在12 h時達到最低.SoAPX5的表達在12 h時有小幅上調,其他基因的表達在處理12 h后都被不同程度地抑制.
SoAPX6經高溫脅迫處理后表達下調.經高溫脅迫12 h后,除SoAPX7外,其他基因的表達都明顯受到抑制,SoAPX3與SoAPX7類似,它們的表達均于高溫脅迫3 h后顯著上調,在其他時長下基本都被抑制.
SoAPX7對干旱處理的響應最為強烈,在處理3 h和12 h后其表達量分別上調到對照組的5.5倍和4.2倍.經干旱處理1 h后,SoAPX1,SoAPX2,SoAPX3和SoAPX6的表達都被抑制,在處理3 h后都出現回升,隨后又呈下降趨勢.SoAPX4和SoAPX5在干旱處理下各個時長的表達相比對照組都有小幅上調.
經氧化脅迫3 h和6 h后,各SoAPX基因的表達都有所上調,處理6 h后表達量的上調最為明顯,其中最明顯的是SoAPX1的表達量達到了對照組的5.2倍,其次是SoAPX4(4.9倍).
3 討 論
植物體內含有多個APX蛋白的編碼基因,本研究在菠菜中發現的7個APX蛋白家族成員,在數量上與擬南芥(8個)、水稻(8個)以及玉米(9個)[11]相近.通過Blast比對發現篩選到的7個APX蛋白均與水稻和擬南芥相對應的氨基酸序列有較高的同源性,這7個蛋白也與茶樹[12]以及白樺[13]的APX蛋白同源性較高,都含有過氧化物酶的保守結構域.亞細胞定位預測到菠菜SoAPX蛋白的表達部位和與其同源的擬南芥、水稻APX并不完全相同,還有待進一步實驗的驗證,然而由多部位的表達可以推測它的功能具有多樣性,可通過多重機制多細胞器配合清除細胞內多余的活性氧,實現抗氧化功能.
植物APX基因家族在植物生長發育和逆境脅迫響應等生理過程中都發揮著非常重要的作用.當植物受到多種生物和非生物脅迫時,APX將通過調控植物的一系列生理生化過程,快速清除細胞中過量的H2O2,使細胞免受活性氧的毒害.本研究結果表明菠菜SoAPX6基因在各個組織均有表達,與劉慧春[14]發現的紅掌AnAPX基因表達相似,由此推斷該基因在菠菜生長發育過程中發揮著多重作用.此外,紅掌AnAPX基因在根中表達最高,與本研究中除SoAPX2外的其他6種菠菜SoAPX基因表達類似.SoAPX2在根中的表達比葉中低很多,這與張蕾等[15]對毛白楊PcAPX進行的組織表達分析結果相似,只是SoAPX2主要在新葉中表達,而PcAPX主要在老葉中表達.SoAPX1和SoAPX3的組織表達模式基本一致,由此推測兩者在植物體內所發揮的作用相似.SoAPX7在雄花和雌花中相對較高的表達量,表明其可能在菠菜性器官的發育過程中起重要作用.
本研究中SoAPX3和SoAPX7在受鹽脅迫的過程中被誘導表達的結果與王超等[13]SsAPX在鹽脅迫下的反應一致,表明它們可能參與了菠菜抗鹽的生理反應,具有保護菠菜免受鹽脅迫造成的氧化損傷的作用.
在本研究中噴施H2O2對菠菜進行模擬氧化脅迫時,7種SoAPX基因的轉錄水平都有不同程度的升高,這可能是由于APX蛋白的主要作用就是清除植物體內的活性氧,當外源H2O2進入植物體內時,APX基因的表達作出響應,以應對氧化脅迫對植物所產生的影響.
4 結 論
在菠菜基因組中鑒定出7個APX基因,對其基因結構、保守基序和進化關系進行了分析.同時對SoAPX在菠菜不同組織中的表達進行了檢測與分析,明確了菠菜SoAPXs的組織表達模式.此外,還對SoAPX在不同環境脅迫下的表達進行分析.這些結果表明菠菜SoAPXs可能在抗鹽、耐寒、抗旱以及抗氧化脅迫中起作用,為后續深入鑒定APX家族成員的功能提供參考.
猜你喜歡
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
山東工業技術(2016年15期)2016-12-01 05:31:22
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·兒童版(2015年1期)2015-07-07 04:12:52
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
